菜粉蝶氣味結(jié)合蛋白和化學(xué)感受蛋白生物信息學(xué)分析
2019-01-24 10:09:28朱宇,劉洋
浙江農(nóng)業(yè)學(xué)報 2019年1期
關(guān)鍵詞:模式識別
朱 宇,劉 洋
(溫州科技職業(yè)學(xué)院,浙江 溫州 325006)
昆蟲的氣味識別系統(tǒng)在昆蟲的取食、交配、生殖、躲避天敵等行為過程中都發(fā)揮著重要的作用。對于昆蟲來說,氣味結(jié)合蛋白(odorant-binding proteins,OBPs)和化學(xué)感受蛋白(chemosensory proteins,CSPs)大量存在于化學(xué)感器的淋巴液中,它們被認為是化學(xué)感受過程中信息素和氣味的載體[1]。在氣味識別的過程中,外界環(huán)境中的化學(xué)信號(信息素、氣味等)與氣味結(jié)合蛋白或化學(xué)感受蛋白相結(jié)合形成復(fù)合物,然后這些化學(xué)信號被轉(zhuǎn)運和釋放到突觸膜,最后激活特異的氣味受體[2]。氣味受體對化學(xué)信號進行識別后,會將化學(xué)信號轉(zhuǎn)變?yōu)閯幼麟娢唬瑐鬟f到昆蟲的中樞神經(jīng)系統(tǒng),引發(fā)相應(yīng)的行為反應(yīng)[2-5]。氣味結(jié)合蛋白和化學(xué)感受蛋白通常存在于昆蟲的觸角、口器等感覺器官中[6]。然而,在昆蟲的其他部位,也發(fā)現(xiàn)了這些蛋白的存在,說明除了檢測和傳遞化學(xué)刺激外,這兩種氣味結(jié)合蛋白還有一些其他的功能[6-11]。
OBPs通常由120~150個氨基酸組成,在N端有信號肽,是一類水溶性的球形蛋白,在無脊椎動物和脊椎動物中都存在,但昆蟲的OBPs與脊椎動物的OBPs序列完全不同源[12]。依據(jù)蛋白序列的特征,昆蟲的OBPs被分為4類,分別是Classical OBPs、Plus-C OBPs、Minus-C OBPs和Atypical OBPs[13-15]。Classic OBPs蛋白序列中包含一個由6個半胱氨酸組成的模式識別序列,這6個半胱氨酸能形成3對二硫鍵[16]。OBPs的第二大類為Plus-C OBPs,大多數(shù)都是在雙翅目昆蟲中被鑒定出,與Classical OBPs相比,Plus-C OBPs的模式識別序列中一般有8個半胱氨酸,能形成4對二硫鍵[13]。Minus-C OBPs從Classic OBPs演化而來,缺少C2-C5這對二硫鍵[17]。Atypical OBPs主要在吸血昆蟲中被鑒定出,它的模式識別序列一般含有9~10個半胱氨酸[15]。昆蟲CSPs的蛋白序列非常保守,典型的CSPs序列有3個共同特征:一般由100~120個氨基酸組成;在N端有一個信號肽;蛋白模式識別序列中包含有4個半胱氨酸[18]。
菜粉蝶屬鱗翅目粉蝶科,是世界范圍內(nèi)為害嚴重的十字花科蔬菜害蟲[19]。目前,菜粉蝶的防治方法以化學(xué)防治為主[20],化學(xué)防治的長期使用會導(dǎo)致菜粉蝶抗藥性的產(chǎn)生以及農(nóng)藥殘留等問題。因此,急需開發(fā)出綠色可持續(xù)的方法來防治菜粉蝶,如利用性信息素或氣味誘集等。本文通過生物信息學(xué)方法在菜粉蝶基因組中找出了24個OBPs和38個CSPs基因序列。根據(jù)多序列比對、系統(tǒng)進化樹構(gòu)建、序列模式識別等方法對鑒定的OBPs和CSPs基因進行驗證和亞型分類。通過對菜粉蝶基因組中OBPs和CSPs基因的查找鑒定,豐富了昆蟲OBPs和CSPs基因數(shù)據(jù)庫,也為菜粉蝶氣味識別和化學(xué)感受相關(guān)實驗和應(yīng)用提供參考。
1 材料與方法
1.1 菜粉蝶OBPs和CSPs基因序列獲取
在NCBI的BioProject數(shù)據(jù)庫 (https://www.ncbi.nlm.nih.gov/bioproject/397594)中下載得到菜粉蝶基因組數(shù)據(jù)庫。從NCBI數(shù)據(jù)庫中獲得鱗翅目和雙翅目的OBPs和CSPs蛋白序列318條作為查詢序列,對菜粉蝶基因組數(shù)據(jù)庫進行本地BLAST(e值為10-5)搜索,得到推測的OBPs和CSPs蛋白序列。然后,通過SignalP 4.1(http://www.cbs.dtu.dk/services/SignalP/)來預(yù)測OBPs和CSPs蛋白N端是否含有信號肽[21]。通過Hmmer(https://www.ebi.ac.uk/Tools/hmmer/)來預(yù)測OBPs和CSPs基因所屬的蛋白家族,篩選出真正的OBPs和CSPs蛋白序列[22]。
1.2 保守序列分析
利用在線網(wǎng)站ScanProsite(https://prosite.expasy.org/scanprosite/)對OBPs和CSPs蛋白序列進行模式識別分析[23]。
1.3 多序列比對和系統(tǒng)進化樹構(gòu)建
利用Kyoto University Bioinformatics Center在線程序clustalw(http://www.genome.jp/tools-bin/clustalw)對蛋白序列進行多序列比對分析,之后利用在線程序ESPript3.0(http://espript.ibcp.fr/ESPript/cgi-bin/ESPript.cgi)繪制多序列比對圖[24-25]。使用MEGA5.10軟件中的鄰位相鄰(neighbor-joining)算法構(gòu)建系統(tǒng)進化樹[26]。具體設(shè)置為:程序重復(fù)次數(shù)為1 000,替換方法Jones-Taylor-Thornton (JTT) model,空缺數(shù)據(jù)的處理方法為95%部分刪除,其他參數(shù)選擇默認設(shè)置[26]。
2 結(jié)果與分析
2.1 菜粉蝶OBPs和CSPs基因的鑒定
通過對菜粉蝶基因組數(shù)據(jù)庫的本地BLAST搜索和Hmmer在線蛋白家族預(yù)測軟件分析,我們得到了24個OBPs基因蛋白序列(表1),我們對這24個蛋白依次命名為PrOBP1-PrOBP24。除PrOBP18缺失N端外,其余OBPs都為全長。PrOBPs的氨基酸大小差異很大,最短的有113個氨基酸,最長的有168個氨基酸,平均長度為141個氨基酸。通過SignalP 4.1在線網(wǎng)站預(yù)測我們發(fā)現(xiàn),除了PrOBP1、PrOBP2、PrOBP3、PrOBP5、PrOBP6、PrOBP14、PrOBP15、PrOBP16外,其他PrOBPs的N端都有信號肽。利用與OBPs相同的搜索方法,我們在菜粉蝶的基因組數(shù)據(jù)庫中得到了38個CSPs基因蛋白序列全長(表1)。我們對這38個蛋白依次命名為PrCSP1-PrCSP38。昆蟲CSPs的序列比較保守,蛋白序列長度相差不大。PrCSPs蛋白氨基酸的平均長度為121個。
通過SignalP 4.1在線網(wǎng)站預(yù)測發(fā)現(xiàn),所有鑒定出的PrCSPs在N端都有信號肽。
2.2 菜粉蝶OBPs和CSPs基因的保守序列分析
通過在線網(wǎng)站ScanProsite篩選和多序列比對分析可知(圖1),鑒定出的24個PrOBPs都具有昆蟲OBPs的半胱氨酸模式識別序列。PrOBP1-PrOBP14、PrOBP16、PrOBP17這16個OBPs屬于Classic OBPs蛋白家族,它們的半胱氨酸模式識別序列為C1-X25-30-C2-X3-C3-X36-42-C4-X8-14-C5-X8-C6(C為半胱氨酸,X為任意氨基酸),與其他鱗翅目Classic OBPs蛋白家族相同[27]。PrOBP15也屬于Classic OBPs蛋白家族,但PrOBP15在C1-C2之間只有19個氨基酸。PrOBP19-PrOBP24這6個OBPs屬于Minus-C OBPs蛋白家族,它們的蛋白序列中缺少C2和C5這兩個半胱氨酸。通過與OBPs相同的分析方法,我們發(fā)現(xiàn)鑒定出的38個PrCSPs的半胱氨酸模式識別序列為C1-X6-C2-X11-19-C3-X2-C4(C為半胱氨酸,X為任意氨基酸),與其他鱗翅目CSPs蛋白家族相同[28](圖2)。
表1菜粉蝶基因組中鑒定得到的氣味結(jié)合蛋白和化學(xué)感受蛋白
Table1PrOBPs and PrCSPs identified from thePierisrapaegenome database
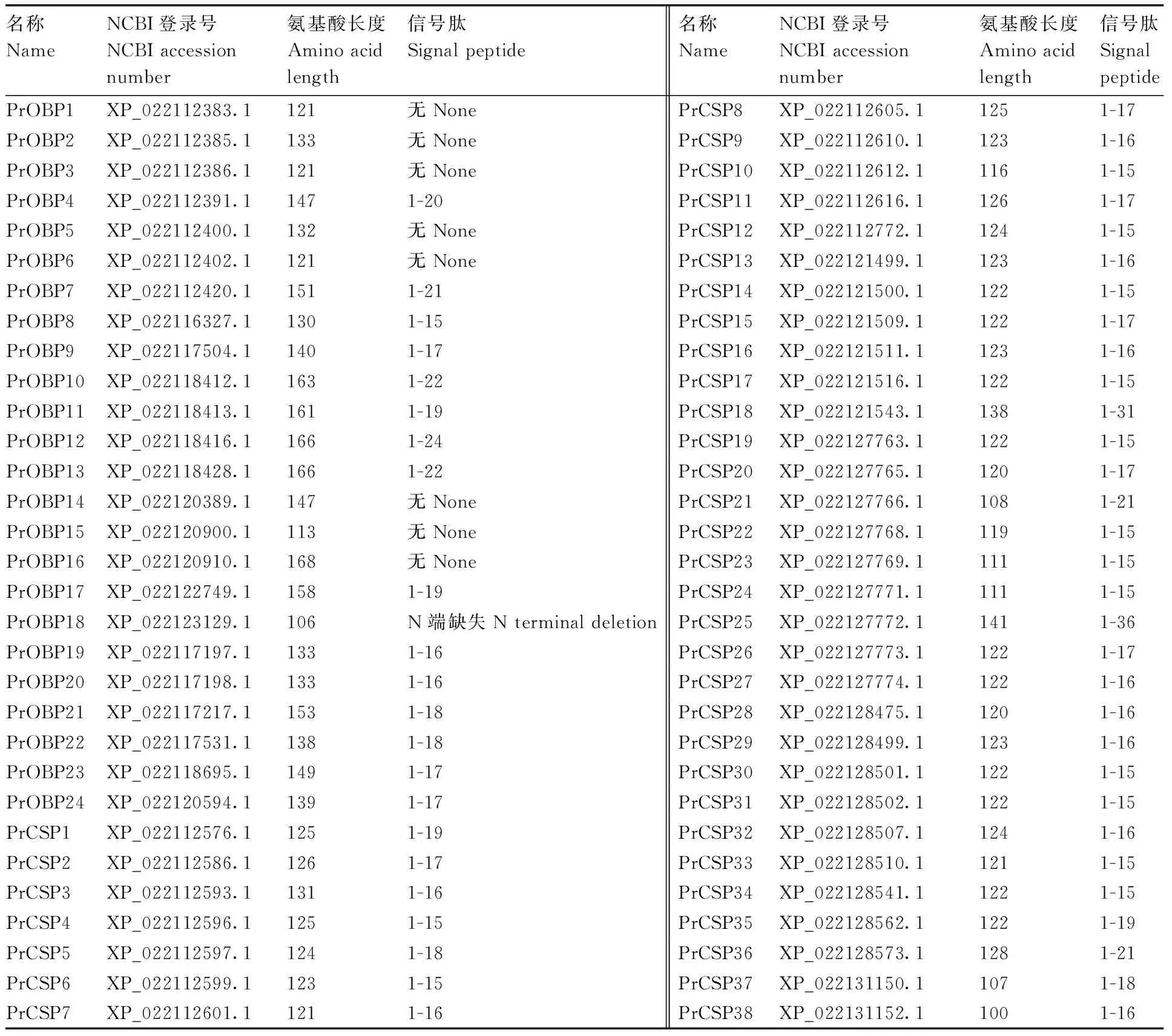
名稱NameNCBI登錄號NCBI accessionnumber氨基酸長度Amino acidlength信號肽Signal peptide名稱NameNCBI登錄號NCBI accessionnumber氨基酸長度Amino acidlength信號肽SignalpeptidePrOBP1XP_022112383.1121無NonePrCSP8XP_022112605.11251-17PrOBP2XP_022112385.1133無NonePrCSP9XP_022112610.11231-16PrOBP3XP_022112386.1121無NonePrCSP10XP_022112612.11161-15PrOBP4XP_022112391.11471-20PrCSP11XP_022112616.11261-17PrOBP5XP_022112400.1132無NonePrCSP12XP_022112772.11241-15PrOBP6XP_022112402.1121無NonePrCSP13XP_022121499.11231-16PrOBP7XP_022112420.11511-21PrCSP14XP_022121500.11221-15PrOBP8XP_022116327.11301-15PrCSP15XP_022121509.11221-17PrOBP9XP_022117504.11401-17PrCSP16XP_022121511.11231-16PrOBP10XP_022118412.11631-22PrCSP17XP_022121516.11221-15PrOBP11XP_022118413.11611-19PrCSP18XP_022121543.11381-31PrOBP12XP_022118416.11661-24PrCSP19XP_022127763.11221-15PrOBP13XP_022118428.11661-22PrCSP20XP_022127765.11201-17PrOBP14XP_022120389.1147無NonePrCSP21XP_022127766.11081-21PrOBP15XP_022120900.1113無NonePrCSP22XP_022127768.11191-15PrOBP16XP_022120910.1168無NonePrCSP23XP_022127769.11111-15PrOBP17XP_022122749.11581-19PrCSP24XP_022127771.11111-15PrOBP18XP_022123129.1106N端缺失N terminal deletionPrCSP25XP_022127772.11411-36PrOBP19XP_022117197.11331-16PrCSP26XP_022127773.11221-17PrOBP20XP_022117198.11331-16PrCSP27XP_022127774.11221-16PrOBP21XP_022117217.11531-18PrCSP28XP_022128475.11201-16PrOBP22XP_022117531.11381-18PrCSP29XP_022128499.11231-16PrOBP23XP_022118695.11491-17PrCSP30XP_022128501.11221-15PrOBP24XP_022120594.11391-17PrCSP31XP_022128502.11221-15PrCSP1XP_022112576.11251-19PrCSP32XP_022128507.11241-16PrCSP2XP_022112586.11261-17PrCSP33XP_022128510.11211-15PrCSP3XP_022112593.11311-16PrCSP34XP_022128541.11221-15PrCSP4XP_022112596.11251-15PrCSP35XP_022128562.11221-19PrCSP5XP_022112597.11241-18PrCSP36XP_022128573.11281-21PrCSP6XP_022112599.11231-15PrCSP37XP_022131150.11071-18PrCSP7XP_022112601.11211-16PrCSP38XP_022131152.11001-16
“無”表示沒有預(yù)測到信號肽。
“None” indicated that no signal peptide was predicted.
2.3 菜粉蝶OBPs和CSPs基因的系統(tǒng)進化分析
我們利用鄰位相鄰算法分別將菜粉蝶OBPs和CSPs與其他報道的鱗翅目昆蟲OBPs和CSPs一起構(gòu)建了系統(tǒng)發(fā)育樹。從菜粉蝶OBPs的系統(tǒng)發(fā)育樹來看(圖3),PrOBPs散落在各個分支上,PrOBP 4和PrOBP17屬于ABPⅠ亞家族;PrOBP1-3,PrOBP5-6和PrOBP8-9屬于ABPⅡ亞家族;PrOBP10-13屬于PBP/GOBP亞家族;PrOBP7、PrOBP14-16和PrOBP18屬于CRLBP亞家族;PrOBP19-24屬于Minus-C OBP家族。總體來說,菜粉蝶OBPs的同源性較低,分化程度較高。從菜粉蝶CSP的系統(tǒng)發(fā)育樹來看(圖4),PrCSP13-19、PrCSP 22-24、PrCSP 29-31和PrCSP 33-36這17個PrCSP聚類到一起,另外21個PrCSP聚類到其他鱗翅目CSPs的分支上。

圖1 PrOBPs的多序列比對Fig.1 Alignment of the identified Pieris rapae OBPs

圖2 PrCSPs的多序列比對Fig.2 Alignment of the identified Pieris rapae CSPs
3 結(jié)論與討論
通過菜粉蝶基因組數(shù)據(jù)的分析,我們得到了24個菜粉蝶OBPs基因,其數(shù)目少于黑腹果蠅Drosophilamelanogaster(51個OBPs)[29]、岡比亞按蚊Anophelesgambiae(57個OBPs)[15]、家蠶Bombyxmori(44個OBPs)[30]、埃及伊蚊Aedesaegypti(66個OBPs)[31]、麗蠅蛹集金小蜂Nasoniavitripennis(90個OBPs)[32]、致倦庫蚊Culexpipiensquinquefasciatus(53個OBPs)[33]。OBP蛋白序列的結(jié)構(gòu)很保守:在蛋白的N端有一信號肽,后面連接著一段含有多個半胱氨酸的結(jié)構(gòu)域序列[12]。通過信號肽預(yù)測,發(fā)現(xiàn)PrOBP1-3、PrOBP5-6、PrOBP14-16這8個OBPs的N端不含有信號肽,說明它們N端序列缺失,不是全長序列。通過半胱氨酸序列的模式識別分析,我們發(fā)現(xiàn)菜粉蝶OBPs主要分布在兩個亞家族:Classic OBPs和Minus-C OBPs,這與其他鱗翅目昆蟲OBPs的分布情況相同[30, 34-35]。從多序列比對結(jié)果來看,菜粉蝶OBPs分子結(jié)構(gòu)變異較大,說明這些從菜粉蝶基因組中,我們鑒定得到38個菜粉蝶CSPs基因,其數(shù)目高于家蠶(24個CSPs)[30]、煙粉虱Bemisiatabaci(19個CSPs)[36],意大利蜜蜂Apismellifera(6個CSPs)[37]、蚜蟲Acyrthosiphonpisum(13個CSPs)[16]。CSPs除了在昆蟲的觸角中表達外,在昆蟲的腿、口器、腹部等部位均有表達[10, 38-39]。在菜粉蝶蛹的轉(zhuǎn)錄組數(shù)據(jù)中(未發(fā)表的數(shù)據(jù)),我們檢測到PrCSP2、PrCSP5、PrCSP19、PrCSP30、PrCSP37、PrCSP38的表達,說明PrCSPs除了氣味識別外,還有一些其他的功能。通過菜粉蝶CSPs進化發(fā)育樹可知,PrCSPs散落在各個分支上,說明菜粉蝶CSPs和其他鱗翅目CSPs有著共同的祖先和相似的功能。PrCSP13-19,PrCSP22-24,PrCSP29-31和PrCSP33-36這17個PrCSP聚類到一起,說明這17個CSPs可能有著相似的起源。
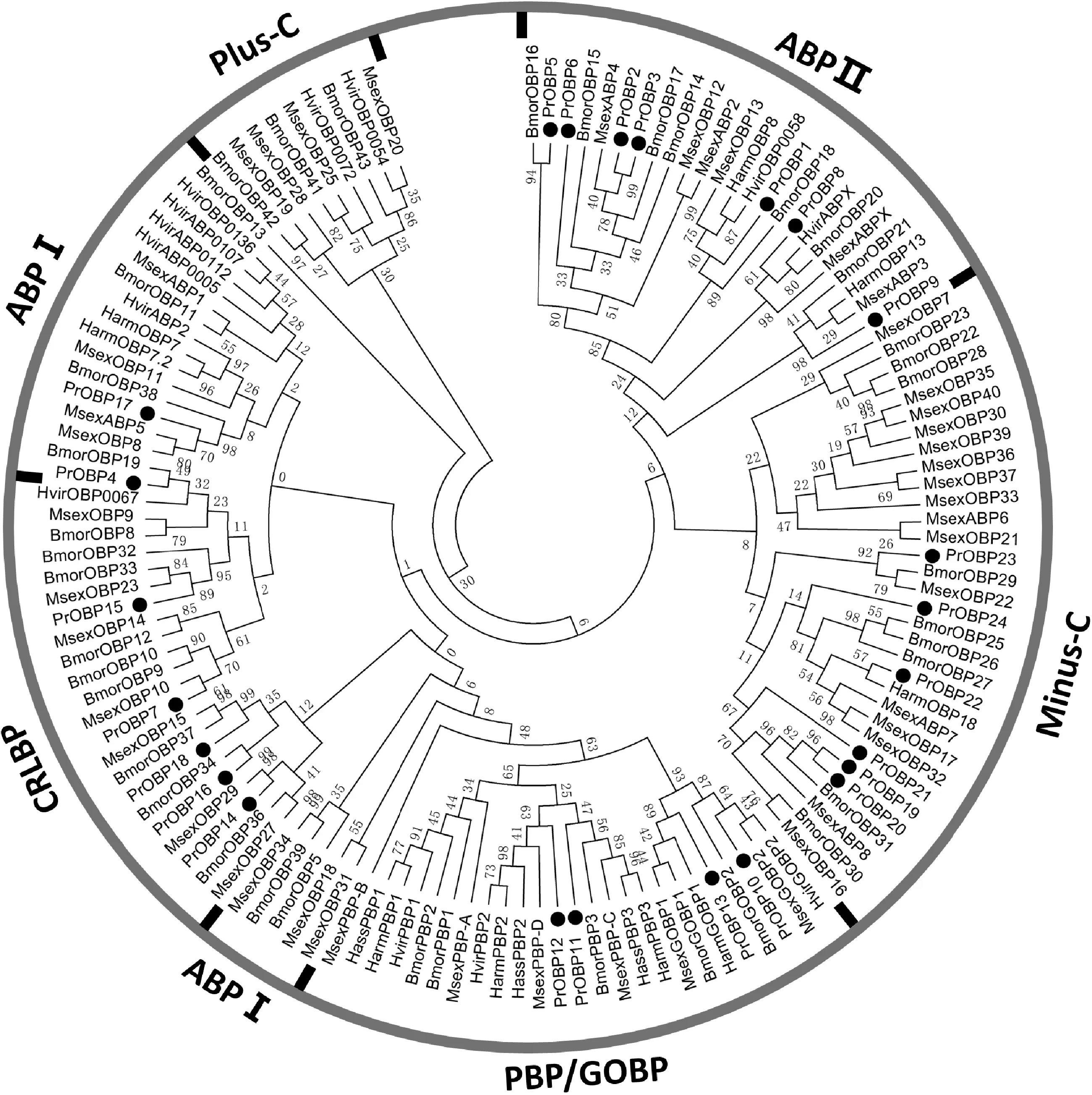
Msex,煙草天峨;Hvir,煙蚜夜蛾;Bmor,家蠶;Harm,棉鈴蟲;Hass,煙青蟲。CRLBP,化學(xué)感受性親脂性配體結(jié)合蛋白;PBP,信息素結(jié)合蛋白,GOBP,一般氣味結(jié)合蛋白;ABP,觸角結(jié)合蛋白。Msex, Manduca sexta; Hvir, Hdiothis virescens; Bmor, Bombyx mori; Harm, Helicoverpa armigera; Hass, Heliothis assulta. CRLBP, Chemical sense-related lipophilic ligand-binding protein; PBP, Pheromone-binding protein; GOBP, General odorant-binding proteins; ABP, Antennal-binding protein.圖3 PrOBPs的進化發(fā)育樹Fig.3 Phylogenetic analysis of PrOBPs
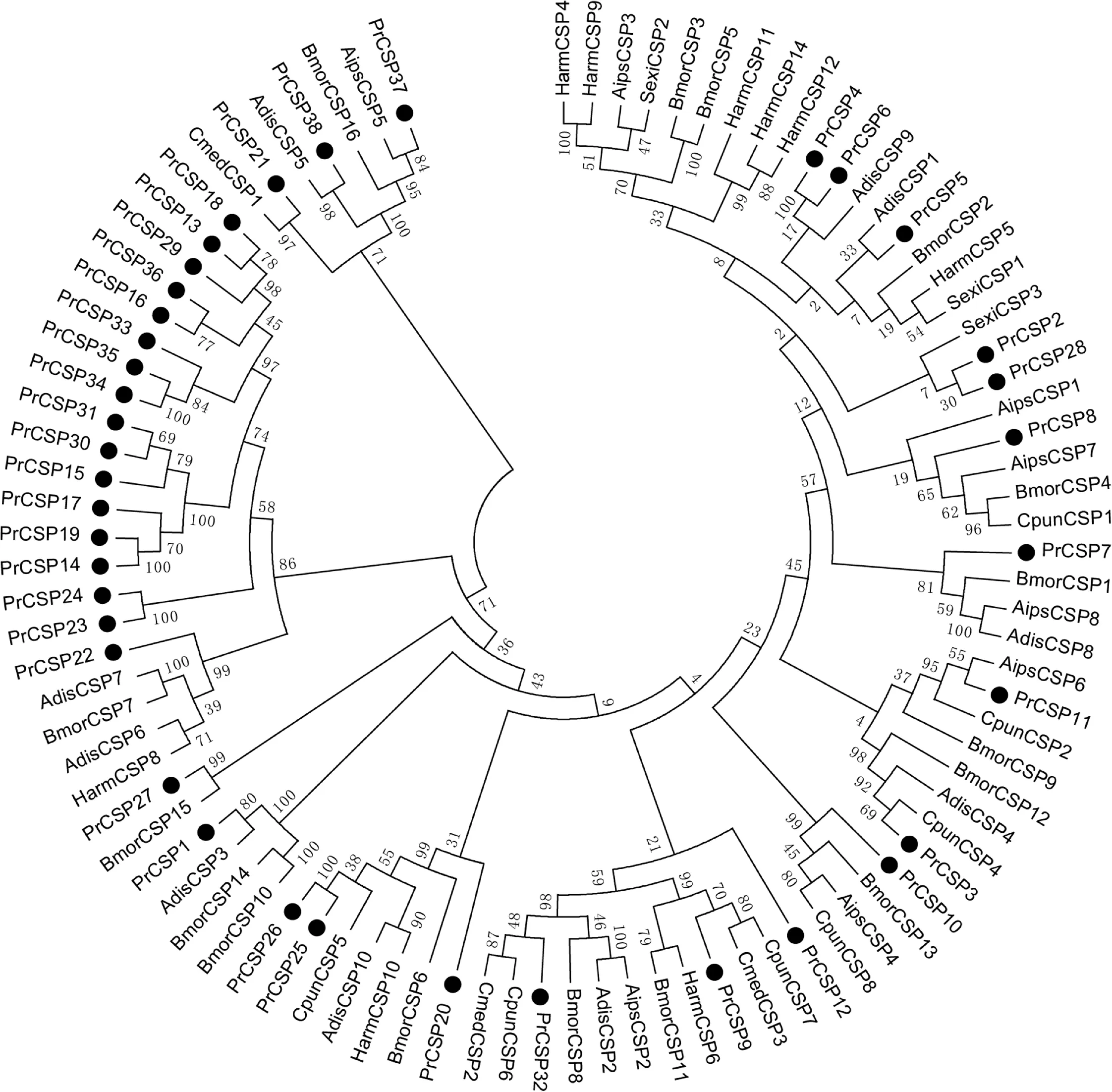
Aips,小地老虎;Bmor,家蠶;Adis,雙委夜蛾;Cmed,稻縱卷葉螟;Harm,棉鈴蟲;Cpun,桃蛀螟;Sexi,甜菜夜蛾。Aips, Agrotis ipsilon; Bmor, Bombyx mori; Adis, Athetis dissimilis; Cmed, Cnaphalocrocis medinalis Giienee; Harm, Helicoverpa armigera; Cpun, Conogethes punctiferalis; Sexi, Spodoptem exigu.圖4 PrCSPs的進化發(fā)育樹Fig.4 Phylogenetic analysis of PrCSPs
OBPs可能發(fā)揮著不同的生物功能。
昆蟲OBP和CSP在植物揮發(fā)物、昆蟲信息素等氣味的識別過程中發(fā)揮著關(guān)鍵作用[2]。本文利用生物信息學(xué)方法,對菜粉蝶基因組中的OBPs和CSPs蛋白家族進行了鑒定,發(fā)現(xiàn)PrOBPs和PrCSPs都具有較高的多樣性。未來,這些鑒定出的PrOBPs和PrCSPs有望用于篩選能與這些氣味結(jié)合蛋白相結(jié)合的化合物,通過化學(xué)合成等技術(shù)開發(fā)出環(huán)境友好的性誘劑或氣味引誘劑來消滅菜粉蝶。同時,也可以通過轉(zhuǎn)基因技術(shù)[40]或CRISPR/Cas9基因編輯技術(shù)[41]來敲除菜粉蝶OBPs和CSPs,通過釋放這些轉(zhuǎn)基因昆蟲到自然界,達到防治菜粉蝶的目的。
猜你喜歡
中成藥(2018年2期)2018-05-09 07:19:52
智能系統(tǒng)學(xué)報(2018年6期)2018-02-04 11:23:57
中成藥(2017年10期)2017-11-16 00:50:05
電子測試(2017年23期)2017-04-04 05:06:50
智能系統(tǒng)學(xué)報(2017年5期)2017-01-22 11:21:30
電測與儀表(2016年6期)2016-04-11 12:07:48
海軍航空大學(xué)學(xué)報(2015年1期)2015-11-11 17:17:57
智能系統(tǒng)學(xué)報(2015年3期)2015-01-29 15:20:12
電測與儀表(2014年15期)2014-04-04 12:05:20
河南科技(2014年5期)2014-02-27 14:08:35
