日本三角渦蟲Caspase-7基因的克隆及功能研究
2019-02-15 05:20:54王志紅王超彭銳
四川動物 2019年1期
王志紅, 王超, 彭銳
(四川大學生命科學學院,生物資源與生態環境教育部重點實驗室,成都610065)
日本三角渦蟲Dugesiajaponica屬于扁形動物門Platyhelminthes渦蟲綱Turbellaria,因其強大的再生能力成為研究再生的最佳模式生物之一(Rink,2013;Mangeletal.,2016)。對于任何類型的創傷,渦蟲幾乎都能夠完成修復和再生,被譽為是“切不死的動物”(Tanetal.,2012)。渦蟲強大的再生能力源于其體內成體干細胞的增殖與分化(Wagneretal.,2011;DeHaan & Ebert,2015)。渦蟲在再生過程中不僅有新組織的產生,創傷后剩余的舊組織(先存組織)還會通過細胞死亡和增殖的相互協調進行身體重塑,以適應身體的比例(Baguá & Romero,1981),因此,在渦蟲再生過程中,細胞凋亡可能起到了重要的作用。
細胞凋亡是細胞死亡的一種形式,用來消除機體中不需要的細胞,從而調節組織內的平衡(Jangetal.,2007)。細胞凋亡主要由Caspase介導。Caspase是一種在多細胞生物中高度保守的含半胱氨酸的天冬氨酸蛋白水解酶,其功能已被廣泛研究(郭素英等,2009),但是Caspase家族在渦蟲中的研究還很少。有學者從渦蟲cDNA文庫中鑒定出了Caspase-likegene 1、2、3,分別命名為Djclg-1、Djclg-2、Djclg-3,并發現只有Djclg-3基因的表達量較高,而Djclg-3基因在控制渦蟲體細胞數量、消除不必要的組織或者細胞,以及在組織器官重塑過程中起到了重要作用,這是第一次對渦蟲凋亡相關基因的報道(Hwangetal.,2004)。Caspase-7與Caspase-3同為Caspase家族中細胞凋亡的重要執行者,執行蛋白質水解的協調程序,并且在癌癥的發生和進展中起重要作用(Thomsenetal.,2013),除了參與細胞的調控外,還參與非凋亡活動的調控,如調節細胞的增殖、分化等(Parketal.,2018),但在渦蟲體內的作用還未見報道。
1 實驗與方法
1.1 實驗材料
日本三角渦蟲由清華大學吳畏教授實驗室提供。室溫下以飲用水飼養,每天換水1次,并隔1天喂食豬肝1次。
1.2 實驗試劑
本實驗中所用rTaq DNA聚合酶、RNAiso、SMARTerTMRACE cDNA Amplification Kit購于TaKaRa公司;瓊脂糖凝膠回收試劑盒購于天根生化科技(北京)有限公司;實時熒光定量PCR(quantitative real-time PCR,qPCR)Master Mix購于Innovagene公司;引物合成及測序由北京六合華大基因科技有限公司完成。
1.3 實驗方法
1.3.1Caspase-7基因的克隆根據SMARTerTMRACE cDNA Amplification Kit說明書制備cDNA模板,利用SMART通用引物和Caspase-7-RACE-F1/R1引物(表1)進行巢式PCR第一輪擴增。然后利用巢式PCR通用引物和Caspase-7-RACE-F2/R2引物(表1)進行第二輪擴增。瓊脂糖凝膠電泳檢測PCR產物,并切膠回收與預期片段大小一致的條帶,將測序結果拼接,與轉錄組測序結果比對。
1.3.2Caspase-7基因全長拼接及序列分析利用MEGA 6對5’-RACE和3’-RACE結果進行拼接;利用DNAMAN將核苷酸序列翻譯為氨基酸后,與其他19個物種(表2)的Caspase-7氨基酸序列進行同源性分析;利用SWISS-MODEL網站在線預測渦蟲Caspase-7蛋白三維結構;最后利用MEGA 6中的鄰接法進行Caspase-7系統發育樹的重建,選擇氨基酸取代模型,并采用自舉檢驗法(bootstrap test,重抽樣1 000次)評估系統發育樹中各節點的置信度。

表2 19個物種Caspase-7的GenBank登錄號Table 2 GenBank accession number of Caspase-7 sequences from 19 species
1.3.3qPCR檢測渦蟲切后再生不同時間點Caspase-7基因的表達量分別提取切后再生0 h、6 h 和1~3 d渦蟲的RNA,以反轉錄后的cDNA為模板,通過qPCR方法檢測Caspase-7基因在不同時間段的表達量,引物序列參見表1。以0 h為對照,β-actin為內參。每組設置3個重復。qPCR體系:2×qPCR Master mix 5 μL,cDNA 0.5 μL,Caspase-7-qPCR正、反向引物各0.3 μL,ddH2O 3.9 μL。qPCR反應條件:95 ℃ 10 min;95 ℃ 15 s,60 ℃退火并延伸60 s,39個循環。結果按2-ΔΔCt法進行計算分析并繪圖。
1.3.4Caspase-7dsRNA體外轉錄及干擾用2對帶有T7啟動子的Caspase-7 PCR引物(表1)從渦蟲cDNA中擴增得到Caspase-7不同區域的DNA,并以此為模板,利用T7 RNA聚合酶在37 ℃體外轉錄獲得干擾用的2條Caspase-7 dsRNA。選取40條長度約1 cm的渦蟲,平均分為2組。一組將制備好的2條Caspase-7 dsRNA與渦蟲食物混合,然后喂食渦蟲,以干擾渦蟲體內Caspase-7基因的表達,作為Caspase-7干擾組;另一組將DEPC水與渦蟲食物混合并喂食渦蟲,作為對照組。Caspase-7干擾組與對照組都隔1天喂食1次,共喂6次,具體方法參見Rouhana等(2013)。
1.3.5Caspase-7干擾效率檢測及干擾后表型觀察
在對渦蟲進行第6次喂食dsRNA后的第4天,渦蟲體內dsRNA被消化后(Rouhanaetal.,2013),分別提取對照組和Caspase-7干擾組的RNA,反轉錄成cDNA,利用Caspase-7 qPCR引物檢測各組Caspase-7基因的表達量。另外,在第6次喂食后的第3天,將渦蟲分為2組:一組于渦蟲咽前部位進行切割,另一組不切割,然后每天觀察并拍照記錄渦蟲形態變化。2組渦蟲在觀察表型的過程中均不喂食。
1.3.6Caspase-7干擾后Djclg-3及PCNA基因表達變化檢測Caspase-7干擾組與對照組渦蟲在不切割直接饑餓7 d后,提取RNA,并反轉錄成cDNA,分別利用Djclg-3基因、PCNA基因和β-actinqPCR引物(表1),通過qPCR技術檢測Djclg-3基因與PCNA基因的表達水平。以β-actin為內參,每組設置3個重復。結果按2-ΔΔCt法進行計算分析并繪圖。
投資活動現金流的使用需要考慮到多種因素,既需要對投資的盈利性、回報時間等方面進行考慮,還需要對資金流總量的合理性進行分析,以此確保現金流管理能夠有效發揮作用。
2 結果
2.1 Caspase-7基因全長克隆及序列分析
對5’-RACE結果中約250 bp的片段(圖1:a中箭頭所指)和3’-RACE結果中約770 bp的片段(圖1:b中箭頭所指)進行瓊脂糖凝膠回收并測序驗證,將5’-RACE和3’-RACE所得序列進行拼接,得到渦蟲Caspase-7基因全長(圖2;GenBank登錄號:MH256116)。該基因全長935 bp,編碼251個氨基酸,5’-UTR區長86 bp,3’-UTR區長93 bp。
為進一步確認所得序列為渦蟲Caspase-7基因序列,將克隆得到的日本三角渦蟲Caspase-7氨基酸序列與其他19個物種進行同源比對,發現Caspase家族中2大保守而關鍵的結構域LSHG和QAC-(Q/R)G(Hwangetal.,2004)也存在于渦蟲Caspase-7基因(圖3)中。在這2大結構域中,組氨酸和半胱氨酸是催化活性中心的重要殘基。
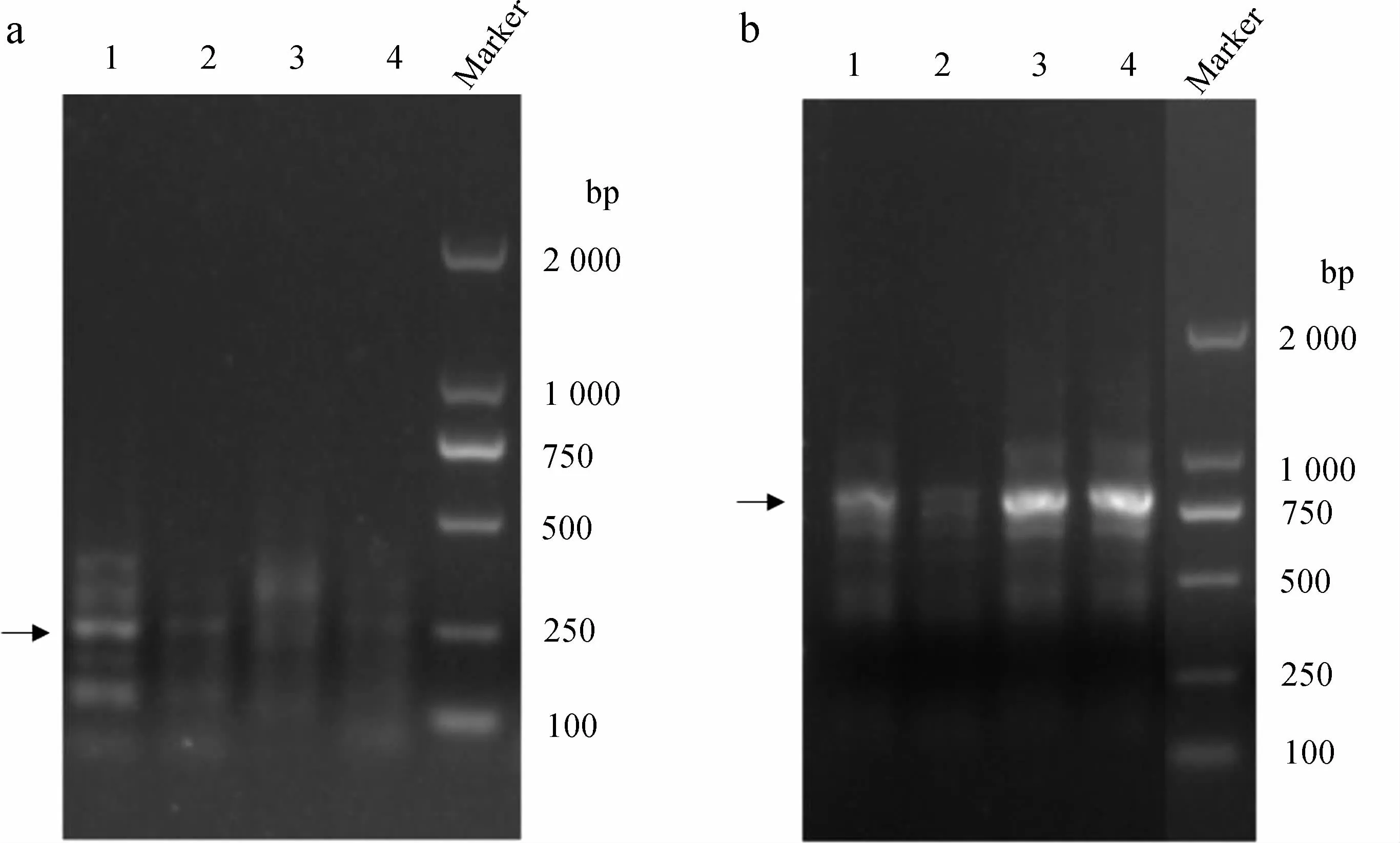
圖1 渦蟲Caspase-7基因RACE擴增結果
Fig. 1 RACE amplification of planarianCaspase-7 gene
a. 5’-RACE, b. 3’-RACE, 1~4. 4個重復組, Marker. DL2000
a. 5’-RACE, b. 3’-RACE, 1-4. 4 replicates, Marker. DL2000

圖2 渦蟲Caspase-7基因全長序列Fig. 2 The complete sequence of planarian Caspase-7 gene
編碼序列上方為Caspase-7基因結構示意圖(共935 bp), 其中, 左右兩端白色區域分別代表5’-UTR(86 bp)和3’-UTR(93 bp)區, 中間陰影部分表示編碼區(CDS, 756 bp), CDS區兩端黑色和灰色部分分別代表ATG起始密碼子和TAA終止密碼子; 堿基序列下方為每個密碼子編碼的氨基酸, *終止密碼子
Upper: a schematic representation of the planarianCaspase-7 gene (935 bp in total), the white regions at the ends of 2 sides represent the 5’-UTR (left, 86 bp) and 3’-UTR (right, 93 bp) regions, respectively, the middle shadow region indicates the coding sequence (CDS, 756 bp), the black and gray portions at both ends of the CDS region represent the ATG start codon and the TAA stop codon, respectively; lower: the complete sequence of planarianCaspase-7 mRNA, amino acids encoded by each codon are shown under the nucleotide sequences, and * represents the stop codon
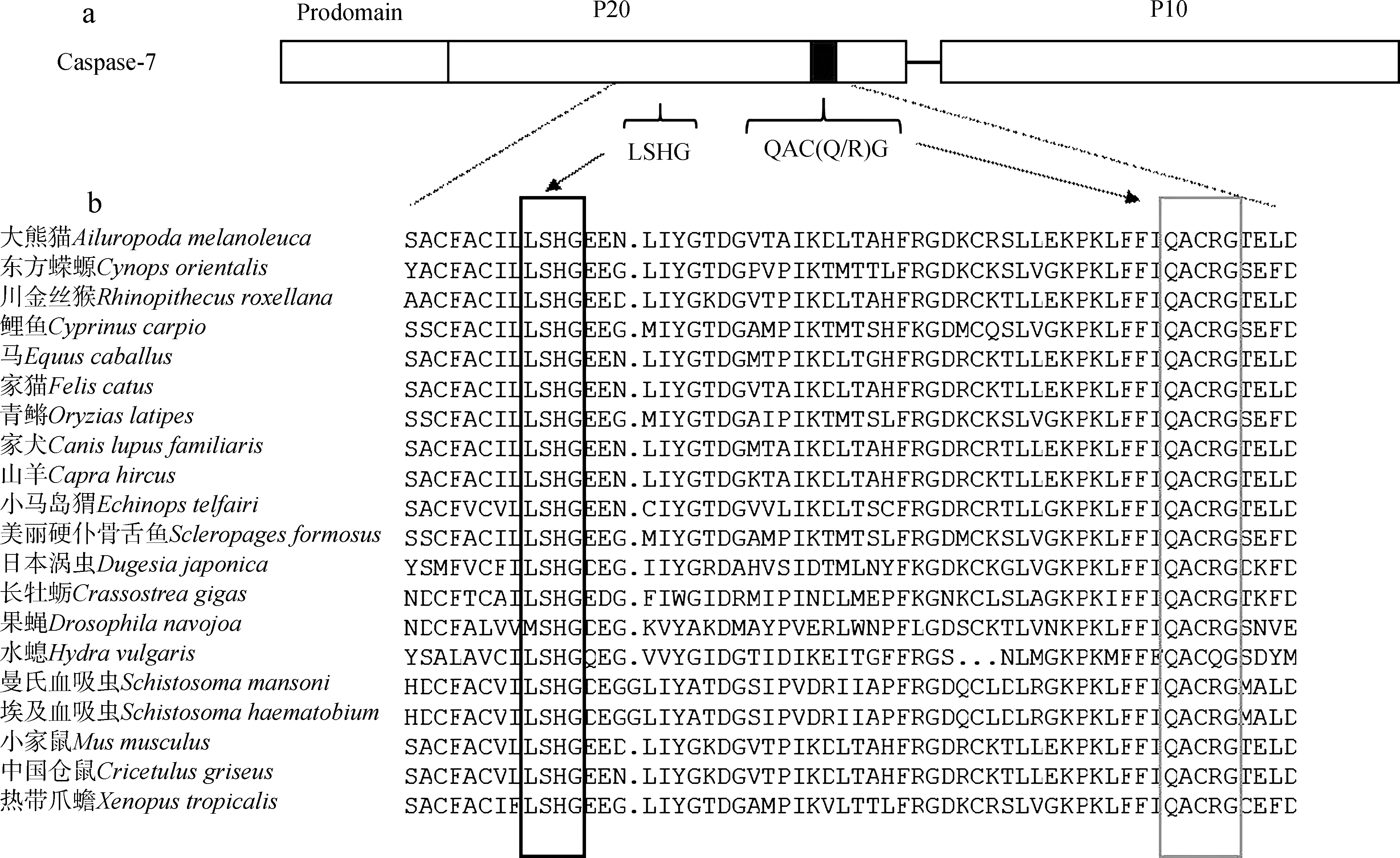
圖3 渦蟲Caspase-7與其他物種同源比對部分結果Fig. 3 Homology comparison of planarian Caspase-7 and other species
a. Caspase-7的結構特征, b. 同源序列比對; 黑色框線和灰色框線為Caspase家族2個保守的關鍵結構域
a. structural characteristics of the Caspase-7, b. homologous sequence alignment; the 2 key domains conserved in the Caspase family are shown in the black frame and gray frame, respectively
2.2 Caspase-7蛋白酶三維結構預測
根據克隆所得的渦蟲Caspase-7氨基酸序列預測得到了其三維結構(圖4),其中包含2個催化單位,每個催化單位都具有5個α螺旋和6個β折疊;渦蟲Caspase-7的潛在催化位點由4個表面環(L1~L4)構成;另外,渦蟲Caspase-7蛋白酶活性中心的關鍵殘基——半胱氨酸殘基(圖4:黑色三角形所指)位于L2環上。
2.3 Caspase-7基因的進化分析
進化樹顯示,渦蟲Caspase-7基因和同為扁形動物門的曼氏血吸蟲Schistosomamansoni及埃及血吸蟲S.haematobium沒有較近的親緣關系,而與刺胞動物門Cnidaria水螅Hydravulgaris有最近的親緣關系(圖5),說明不同物種之間Caspase-7基因可能發生了趨同進化。
2.4 渦蟲再生不同時間段Caspase-7表達量變化
為分析Caspase-7基因是否參與渦蟲細胞凋亡調控,通過qPCR檢測渦蟲切后再生不同時間段Caspase-7基因的表達量變化(圖6)。Caspase-7基因在渦蟲切后再生6 h輕微升高,在渦蟲切后再生3 d升高明顯(P<0.05),其余時間點變化不明顯。

圖4 渦蟲Caspase-7蛋白三維結構的預測Fig. 4 Predicted three-dimensional structure of planarian Caspase-7 protein
α1~α5(α1’~α5’)分別代表5個α螺旋; L1~L4(L1’~L4’)分別代表4個表面環; 2個黑色三角形所指為半胱氨酸殘基所在位點
α1-α5 (α1’-α5’) represent 5 alpha helices, respectively; L1-L4 (L1’-L4’) represent 4 surface loops, respectively; two black triangles point to the cysteine site, key amino acid residue in active center
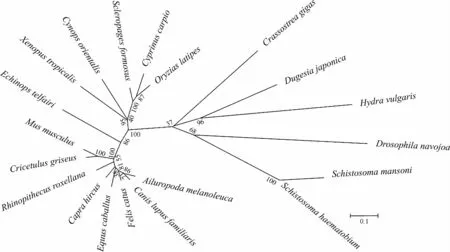
圖5 渦蟲與其他物種Caspase-7基因進化樹
Fig. 5 Phylogenetic analysis ofCaspase-7 gene between planarian and other species
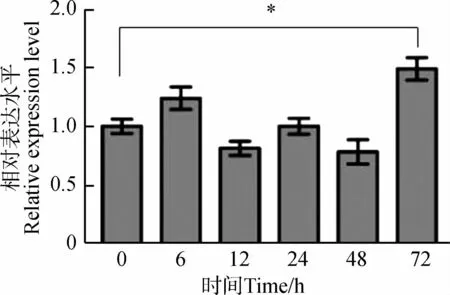
圖6 渦蟲切后再生不同時間段Caspase-7基因表達量變化Fig. 6 The expression changes of Caspase-7 gene at different times during planarian regeneration
*P<0.05
2.5 干擾Caspase-7基因影響渦蟲組織重塑
為了進一步分析Caspase-7基因在渦蟲中的作用,通過喂食Caspase-7 dsRNA干擾渦蟲體內Caspase-7基因的表達。qPCR結果顯示,喂食Caspase-7 dsRNA能成功干擾渦蟲體內的Caspase-7基因的表達(圖7:a,P<0.01)。Caspase-7干擾組渦蟲切后的頭部片段(5/5)和尾部片段(5/5)均能完成再生,說明干擾Caspase-7并不影響渦蟲的再生。但在Caspase-7干擾組渦蟲切后的第11天,其頭部及尾部發生溶解(5/5)(圖7:d),而此時對照組渦蟲形態仍然正常(5/5)。此外,Caspase-7干擾組未切割渦蟲在饑餓7 d后(圖7:c),頭部和尾部也發生部分溶解(5/5),而對照組正常(5/5)。這說明干擾Caspase-7的表達對饑餓環境下的成體渦蟲(再生完成后的或未切割的成蟲)會造成明顯的影響,導致蟲體發生溶解。
2.6 Caspase-7干擾后Djclg-3與PCNA基因表達變化
Caspase-7干擾組渦蟲在饑餓環境下發生溶解時,檢測細胞凋亡的另一關鍵基因Djclg-3以及增殖相關基因PCNA的表達變化。qPCR結果顯示,成體渦蟲饑餓處理7 d后,Djclg-3和PCNA基因的表達量相較于對照組顯著降低(P<0.001),即細胞凋亡與增殖大幅度減少(圖8)。
3 討論
Caspase-7是Caspase家族的重要成員之一,在細胞凋亡過程中發揮著促凋亡作用。未激活的Caspase-7以酶原的形式存在,一旦被上游分子剪切激活后,活化的Caspase-7可以對其底物進行切割,導致細胞凋亡發生(Hilletal.,2016)。細胞凋亡的缺失會導致多種癌癥的發生(Puttetal.,2006;Wolanetal.,2009;Thomsenetal.,2013)。此外,Caspase-7還在炎癥反應、神經激活和細胞分化中起著重要的作用(Martinon & Tschopp,2007;Kuranaga,2012)。Caspase-7不僅參與凋亡的調控,還參與調節細胞周期、細胞增殖及分化等(Chaudharyetal.,2016;Leietal.,2017;Parketal.,2018)。

圖7 干擾Caspase-7對渦蟲的影響Fig. 7 Effect of Caspase-7 interference on planarian growth
a.Caspase-7干擾檢測結果, b. 渦蟲于咽前切后再生頭部第6天的表型, c. 未切割渦蟲饑餓第7天的表型, d. 渦蟲于咽前切后再生頭部第11天的表型;**P<0.01
a. theCaspase-7 interference efficiency, b. the regenerative planarian heads after 6 days of amputation, c. the planarians after starvation for 7 days, d. the regenerative planarian heads after 11 days of amputation;**P<0.01
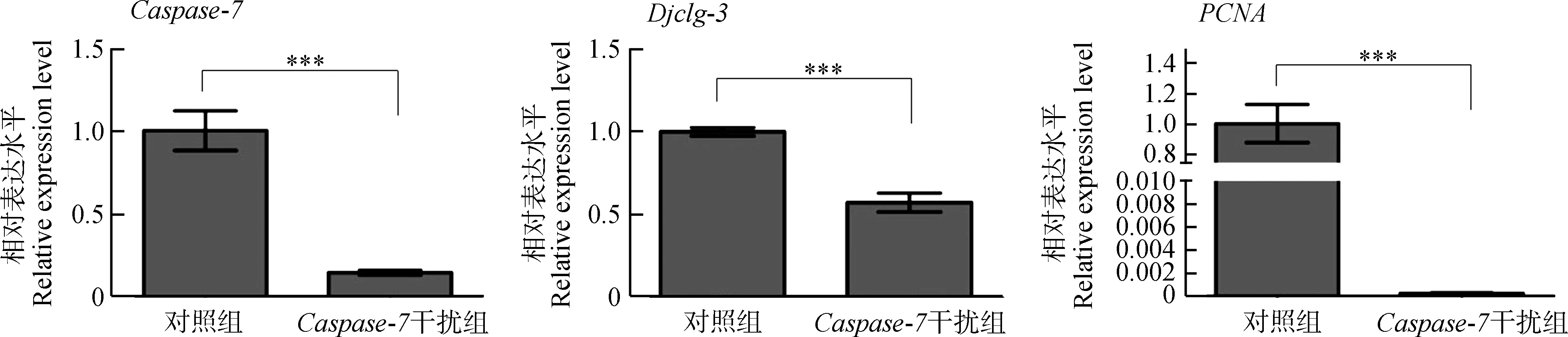
圖8 Caspase-7干擾后各基因表達變化Fig. 8 Gene expression changes after Caspase-7 interference
***P<0.001
本研究發現,在渦蟲切后再生6 h和3 d,Caspase-7基因表達量升高,這2個時間點是渦蟲切后凋亡發生的2個高峰期(Hwangetal.,2004),這表明Caspase-7可能在渦蟲中參與細胞凋亡的調控。進一步分析發現,干擾Caspase-7基因后,渦蟲再生沒有受到影響,但再生完成后或未切割的成體渦蟲在饑餓環境下,Caspase-7干擾組蟲體會逐漸溶解直至死亡。其發生溶解時,Djclg-3與PCNA基因表達量顯著降低。
細胞凋亡在渦蟲組織重塑的過程中起著重要的作用(Pellettierietal.,2010)。Caspase-3是Caspase家族中另一重要的凋亡執行者(Chaietal.,2001;Jangetal.,2007),可以被Caspase家族中凋亡起始者Caspase-8和Caspase-9所剪切,活化的Caspase-3可以降解多種細胞蛋白,并在細胞凋亡過程中負責細胞形態的改變和DNA的斷裂,同時還可以調節細胞的增殖和組織器官的尺寸(Liuetal.,2017;Yosefzonetal.,2018)。已有研究表明,Djclg-3基因在渦蟲再生過程中參與渦蟲細胞數量的控制,對組織重塑過程有調控作用(Hwangetal.,2004)。在饑餓環境下,渦蟲沒有外在能量的供應,只能靠自身細胞的死亡來提供生存所需的能量和物質,因此渦蟲會發生退行性生長,即渦蟲軀體會不斷縮小。此時,渦蟲組織和器官會通過細胞死亡和增殖的協調而重塑,以適應不斷縮小的身體比例(Mead & Christman,1998;Weissman,2000)。受Caspase-7基因干擾后,Djclg-3與PCNA基因的表達量顯著降低,可能導致饑餓環境下渦蟲的細胞凋亡與增殖減少,渦蟲組織重塑難以完成,以至于蟲體不斷溶解并最終死亡。該結果說明,Caspase-7可能在渦蟲的組織重塑中發揮著重要作用。
本文首次報道渦蟲中Caspase-7基因全長并分析了其結構特點,檢測了渦蟲再生不同時間點的表達量變化,并對Caspase-7在渦蟲中的功能進行了初步探索,為渦蟲細胞凋亡的研究及細胞凋亡與再生關系的探索奠定了基礎。
