外源性雄激素通過誘導自噬促大鼠睪丸萎縮
2019-02-15 10:00:24單光明聶黎虹郭鳳英朱亞洲秦凱悅杜丹丹田國林趙瑞寧
中國老年學雜志 2019年2期
關鍵詞:水平
單光明 聶黎虹 郭鳳英 朱亞洲 秦凱悅 杜丹丹 田國林 趙瑞寧
(寧夏醫科大學 1總醫院泌尿外科,寧夏 銀川 750004;2研究生學院;3基礎醫學院;4 2015級本科生)
睪丸是男性生殖器官的一部分,具有分泌雄性激素、產生精子、維持正常男性生育功能和性功能的作用〔1〕。由于雄激素特殊的生理功能,部分人群選擇外源性給予雄激素以達到提高性欲、提高運動成績、男性藥物避孕等目的〔2,3〕。但外源性雄激素的不合理應用,可能致使睪丸萎縮,進而影響其正常生理功能〔2〕。目前外源性雄激素致睪丸萎縮的機制尚未闡明。自噬是真核生物細胞調節自身生長、死亡、能量代謝的重要機制,其在增生、衰老、退行性變、炎癥變等病理生理過程中發揮重要的作用〔4,5〕。研究表明,雄激素可以調控細胞自噬的水平〔6〕;而精索靜脈曲張可致睪丸生精小管上皮細胞自噬水平上調〔4〕。關于外源性雄激素是否通過調節細胞自噬水平參與睪丸萎縮病理過程的發生未見報道。本研究通過外源性給予雄激素丙酸睪丸酮(TP)及自噬抑制劑,采用分子生物學及形態學等技術,探討外源性雄激素是否通過調節細胞自噬水平變化參與睪丸萎縮病理過程的發生。
1 材料與方法
1.1動物與主要試劑 雄性SD大鼠,體重250~270 g,購于寧夏醫科大學實驗動物中心。TP(北京Solarbio公司,中國);氯喹(CQ,Sigma公司,美國);甘草甜素(Gly,東京化成工業株式會社,日本);全蛋白提取試劑盒、電化學發光(ECL)檢測試劑盒;二喹啉甲酸(BCA)蛋白濃度測定試劑盒(江蘇凱基生物,中國);兔抗Beclin-1;兔抗p62(Proteintech公司,美國);兔抗LC3B(Sigma公司,美國);兔抗HMGB1(Abcam公司,英國);鼠抗β-actin和辣根過氧化物酶(HRP)標記的二抗和HE染料(北京中杉金橋公司,中國)。
1.2動物分組與模型建立 48只SD大鼠隨機分為6組:對照(control)組、TP組、TP+CQ組、TP+Gly組、CQ組、Gly組,每組8只。TP組皮下注射TP(3 mg·kg-1·d-1);TP+CQ組皮下注射TP(3 mg·kg-1·d-1)加腹腔注射CQ(50 mg·kg-1·d-1);TP+Gly組皮下注射TP(3 mg·kg-1·d-1)加腹腔注射Gly(10 mg·kg-1·d-1);CQ組僅腹腔注射CQ(50 mg·kg-1·d-1);Gly組僅腹腔注射Gly(10 mg·kg-1·d-1);control組腹腔注射生理鹽水(2.5 ml·kg-1·d-1)。每組處理時間均為21 d。
1.3標本收集與睪丸指數計算 各組稱重后麻醉(10%水合氯醛,0.3 ml/100 g),摘取睪丸組織,迅速測量睪丸濕重,計算睪丸指數。大鼠睪丸指數=睪丸濕重(g)/體重(kg)。測量后兩側睪丸分別進行病理學和Western印跡法蛋白檢測。
1.4HE染色觀察睪丸組織形態學改變 取一側睪丸用冷生理鹽水沖洗后,置于4%多聚甲醛中固定,24 h后將其修剪后置于包埋盒中脫水過夜;次日進行浸蠟、包埋,冷凝后進行組織切片、烤片;將切片進行脫蠟水化并染色,流水沖洗,梯度酒精脫水,二甲苯透明,中性樹膠封片;在光學顯微鏡下觀察睪丸組織病理學變化。
1.5Western印跡法檢測自噬相關蛋白表達水平 取另一側大鼠睪丸,全蛋白提取試劑盒提取蛋白,BCA法檢測蛋白濃度后定量。將每組蛋白60 μg加入不同泳道中行電泳70 min后,采用半干轉方式進行轉膜。5%牛奶室溫封閉1.5 h,磷酸鹽吐溫緩沖液(PBST)洗膜3次,加一抗4℃孵育過夜后,PBST洗膜3次,二抗室溫孵育1.5 h,PBST洗膜3次,ECL化學發光液顯色,于凝膠成像儀拍照。
1.6統計學處理 采用SPSS22.0軟件進行析因設計的方差分析。
2 結 果
2.1各組體重、睪丸濕重及睪丸指數的比較 與control組比較,TP組睪丸濕重、指數均顯著下降(P<0.01);與TP組比較,TP+CQ和TP+Gly組睪丸指數均顯著上升(P<0.05);TP+CQ和TP+Gly組與control組比較,睪丸濕重顯著減輕(P<0.05),但睪丸指數無明顯變化(P>0.05);CQ和Gly組與control組比較,睪丸濕重和指數均無明顯差異(P>0.05);見表1 。
2.2各組睪丸組織形態學分析 control組睪丸生精小管形態規則,各級生精細胞層次分明、有序排列;管腔內可見大量成熟的精子,旋渦狀表現。TP組生精小管形態不規則;生精細胞層數明顯減少、排列紊亂;管腔內成熟的精子稀少,部分管腔內出現脫落的生精細胞團塊。TP+CQ組和TP+Gly組生精小管管腔稍不規則,與TP組比較生精細胞層數和排列狀況均明顯改善;管腔內可見成熟的精子,未見脫落的生精細胞團塊。CQ組和Gly組睪丸生精小管形態、結構與control組類似,見圖1。

表1 各組體重、睪丸濕重和睪丸指數的比較
與 control組比較:1)P<0.05,2)P<0.01;與TP組比較:3)P<0.05,4)P<0.01

圖1 各組睪丸組織染色
2.3各組睪丸組織相關蛋白的表達 與control組(0.66±0.06,0.69±0.10,0.77±0.07)比較,TP組睪丸組織Beclin-1、LC3B-Ⅱ和HMGB1蛋白表達水平(0.91±0.09,1.05±0.08,1.07±0.15)均顯著上調(P<0.05),p62蛋白表達無統計學差異(0.50±0.04,0.46±0.10,P>0.05)。與TP組比較,TP+CQ組睪丸組織Beclin-1、LC3B-Ⅱ和HMGB1蛋白表達水平(0.65±0.06,0.71±0.04,0.74±0.08)均顯著下調(P<0.05),p62蛋白表達仍無統計學差異(0.53±0.06,P>0.05);TP+Gly組上述各蛋白表達(0.58±0.05,0.74±0.04,0.82±0.06,0.50±0.09)與TP+CQ組結果類似,兩組各蛋白表達均無統計學差異(P>0.05)。CQ組上述各蛋白表達(0.58±0.07,0.61±0.04,0.84±0.08,0.48±0.04)和Gly組(0.65±0.13,0.61±0.05,0.80±0.11,0.46±0.07)與control組無統計學差異(P>0.05),見圖2。
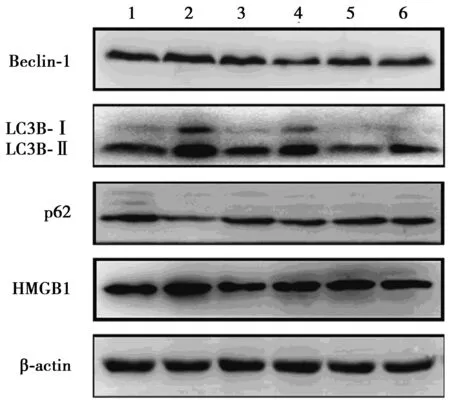
1~6:control組、TP組、TP+CQ組、TP+Gly組、CQ組、Gly組圖2 各組睪丸組織相關蛋白的表達
3 討 論
雄激素主要由睪丸間質細胞合成、分泌,可發揮促男性性器官成熟、第二性征出現、維持正常性欲和生殖等作用。在睪丸內,雄激素通過受體介導影響睪丸支持細胞、間質細胞、血管平滑肌細胞及內皮細胞的發育,進而促進各級生精細胞的分化及精子的成熟。體內雄激素水平異常變化會影響睪丸結構與功能。外源性雄激素可通過影響生精細胞的分化,致使成熟精子數量明顯減少以達到男性避孕的目的〔3〕。研究表明,雄激素可以調控細胞自噬的水平;其可通過增加細胞自噬和自噬流發揮促前列腺癌細胞代謝和增殖的作用〔6〕;雄激素受體可以通過轉錄調控自噬核心基因和溶酶體基因促進前列腺癌的發展〔7〕。且在精索靜脈曲張等疾病發生過程中,睪丸細胞自噬水平也發生改變〔4〕。這些現象提示雄激素可能通過調控細胞自噬參與睪丸萎縮的病理過程。
本研究表明外源性雄激素可致睪丸萎縮。LC3是自噬啟動的特異性標記蛋白,LC3B-Ⅱ的蛋白水平是反映自噬最基本的指標。Beclin-1是首個被鑒定介導哺乳動物自噬的蛋白,在自噬體形成過程中是不可或缺的條件〔8〕。本文結果提示雄激素致睪丸萎縮過程中可能上調了細胞自噬水平。HMGB1
是重要的自噬調控因子〔9,10〕,細胞內不同空間定位的HMGB1均可通過相應機制調控自噬發生。那么表達增多的HMGB1是否參與了雄激素致睪丸萎縮過程中自噬水平上調呢?本文提示細胞自噬水平的上調參與了雄激素致睪丸萎縮的病理過程。睪丸萎縮過程中表達增多的HMGB1參與了睪丸細胞自噬水平的上調。
綜上,外源性雄激素可能通過上調自噬水平引發大鼠睪丸萎縮病理過程的發生,HMGB1可能在此過程中調節自噬水平改變。但在睪丸萎縮病理過程中,雄激素通過何種方式調控自噬,HMGB1在此過程中如何發揮促自噬的作用及雄激素和HMGB1是否具有協同調控自噬的作用均需進一步探討。
猜你喜歡
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
火花(2019年12期)2019-12-26 01:00:28
人大建設(2019年6期)2019-10-08 08:55:48
人大建設(2019年12期)2019-05-21 02:55:32
雜文月刊(2018年21期)2019-01-05 05:55:28
人大建設(2017年6期)2017-09-26 11:50:44
學苑創造·A版(2015年11期)2016-01-14 09:03:27
俄羅斯問題研究(2012年1期)2012-03-25 09:54:45
中國火炬(2010年12期)2010-07-25 13:26:22
中國火炬(2010年8期)2010-07-25 11:34:30
