四川部分地區禽源空腸彎曲菌MLST分型及遺傳進化分析
2019-02-19 11:00:04,,,,,,,,
中國人獸共患病學報 2019年1期
, , ,,,, ,,
空腸彎曲菌(Campylobacterjejuni)是廣泛存在于家禽、家畜等動物體內的一種常見的人畜共患的食源性感染致病菌,主要引起人的急性胃腸炎,進入血液會引發敗血癥、腦炎、心內膜炎、心肌炎、反應性關節炎和泌尿系統感染等,嚴重的還會引發神經系統損傷性疾病格林巴利綜合征(Guillain-Barre syndrome, GBS)[1-2]。禽類是空腸彎曲菌最主要的傳染源,消費禽肉及制品與該菌的流行緊密相關,故而該菌早已成為全球公共衛生關注的重點[3],因此對于空腸彎曲菌的分類研究,以探究其致病相關性顯得尤為重要。
多位點序列分型(Multilocus sequence typing,MLST)技術是一種基于核酸序列測定的新型細菌分型方法,其與多位點酶電泳分析(Multilocus enzyme electrophoresis,MEE)原理相同,不同的是MLST通過測定細菌7個管家基因的核苷酸序列后根據序列的不同,經比對后分別命名不同序列號管家基因的等位基因值,7個管家基因不同序列號的組合決定菌株特異的序列型(Sequence type, ST)[4-5]。MLST數據庫針對每個菌株個體都有相應的、唯一的ST型別且每個菌屬的管家基因及引物均被標準化與統一,以便全球菌株MLST結果的比較分析。
依此,本研究以四川部分地區分離到的48株禽源空腸彎曲菌作為研究載體,使用MLST分子分型技術對其進行分子分型研究及遺傳進化分析,以期進一步從分子水平研究該菌的遺傳特性及分布特點。
1 材料與方法
1.1材料 48株分離株源于對四川多地區采集的禽類盲腸及內容物樣品的分離[6],哥倫比亞培養基、Skirrow培養基及其對應抗生素添加劑、腦心浸液均購自于成都康迪生物科技有限公司,細菌基因組提取試劑盒、膠回收試劑盒均購自于天根生化科技(北京)有限公司,其他試劑均為國產分析純產品。
1.2分離株MLST分型方法、反應體系及參數 參照MLST數據庫(http://www.mlst.net/ databases/)推薦的方法選取空腸彎曲菌 MLST分型相關的7個管家基因(aspA,glnA,glyA,gltA,tkt,pgm和uncA)作為目的基因,分別進行擴增與測序。
反應體系及參數:反應體系為25 μL:其中2×TaqPCR Mix 12.5 μL,引物(10 μmol/L)各1 μL,模板2 μL,ddH2O補足至25 μL;反應參數為:94 ℃ 5 min;94 ℃ 30 s、50 ℃ 1 min、72 ℃ 1 min,35個循環,72 ℃ 10 min。反應結束后,將產物經瓊脂糖凝膠電泳后,使用試劑盒回收條帶單一且明亮的擴增片段送至華大基因測序。
1.3序列類型(Sequence type,ST)的確定 對7個管家基因的雙向測序圖譜進行比對,確保序列一致后將不同的基因序列在MLST數據庫中提交,進行檢索比對。根據數據庫提供的每種等位基因的序列號,確定7個管家基因的組合,即為該菌株的ST型,并根據該菌株的ST型在數據庫中的排列序號對其命名。經命名的ST型或出現新ST型,其歸因于兩種情況,存在新的等位基因序列或存在各等位基因位點的新組合[7]。相似的ST型菌株可組成一個ST序列群,即一個克隆系(Clonal complex,CC)[8]。
1.4遺傳進化分析 確定所有分離株的ST型別及克隆系后,從MLST數據庫網站(http://www.mlst.net/databases/)下載分離株及近5年國內外多個國家不同源分離株的7個等位基因的串聯序列(按照aspA-glnA-gltA-glyA-pgm-tkt-uncA的順序排列),采用MEGA6.0軟件中的Neighbor-jioning方法構建系統進化樹,分析本次分離株和其他地區分離株之間的遺傳進化關系。
2 結 果
2.1分離株管家基因擴增結果 使用空腸彎曲菌7個管家基因擴增引物對分離株的管家基因進行擴增,產物經瓊脂糖凝膠電泳后,結果顯示(圖1,其中1株分離株)各管家基因的片段大小與預期結果一致。
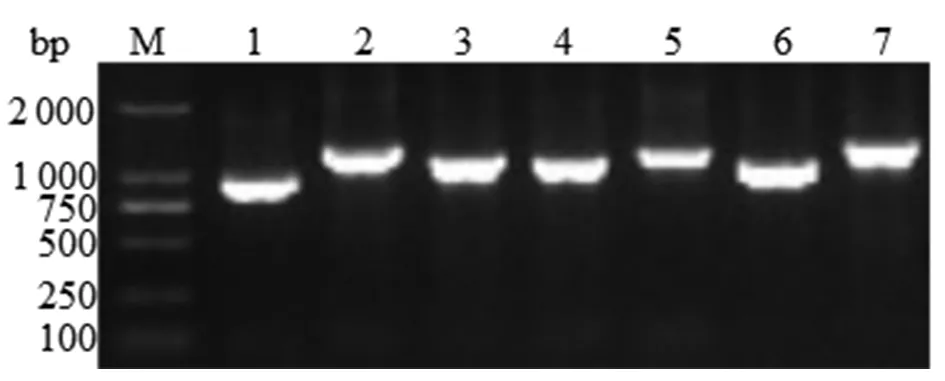
Note: M: DNA Marker; 1:glyA;2:glnA;3:gltA;4:tkt;5:uncA;6:aspA;7:pgm圖1 空腸彎曲菌的MLST擴增結果Fig.1 MLST amplification results of Campylobacter jejuni
2.2 MLST分型結果與分析
2.2.1新序列型及新等位基因值 48株分離株中共含24種不同的序列型,其中22株分離株含有新序列型屬11個新序列型,占分離株的45.83%。新序列型中眉山鵪鶉源的8株分離株和雅安雞源的3
株分離株全為新序列型;溫江雞源的分離株中有7株屬于新序列型,占溫江雞源分離株的26.92%;溫江鴨源的分離株中有1株屬于新序列型;金堂雞源的分離株有3株屬于新序列型,占金堂分離株的33.33%。
48株分離株中共有14株分離株含有新等位基因序列,7個管家基因中的aspA,pgm,tkt,uncA基因存在新的等位基因序列,而其余3個等位基因均未發現新的等位基因序列。14株分離株各存在1個新等位基因序列,其中tkt位點共有10株分離株(V5、V6、V13-20)發現新等位基因序列,分離株V26、V27的pgm位點為新等位基因序列,aspA和uncA位點各有1株分離株為新等位基因序列;分離株中的V26、V27所屬的序列型有對應的克隆系均為ST-1332CC。除此之外,另有8株分離株(C2、C4、V7、V8、V25、V33、V34、V47)存在等位基因位點的新組合,屬于新序列型且均無對應的克隆系,為獨特型(表1)。
表1 空腸彎曲菌分離株中的新等位基因及新序列型
Tab.1 New allele sequences and new series of alleles ofCampylobacterjejuniisolates
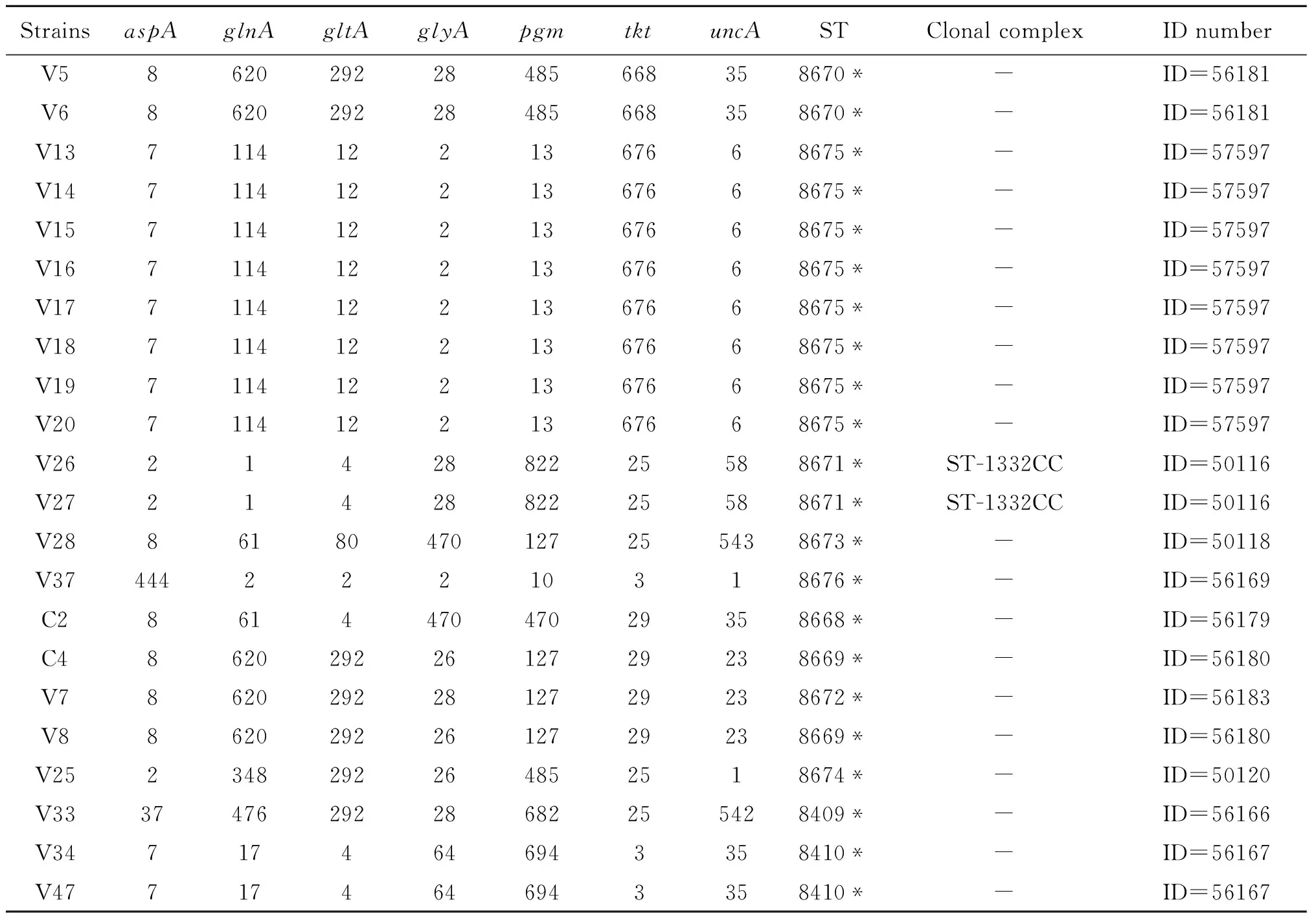
StrainsaspAglnAgltAglyApgmtktuncASTClonal complexID numberV5862029228485668358670*-ID=56181V6862029228485668358670*-ID=56181V1371141221367668675*-ID=57597V1471141221367668675*-ID=57597V1571141221367668675*-ID=57597V1671141221367668675*-ID=57597V1771141221367668675*-ID=57597V1871141221367668675*-ID=57597V1971141221367668675*-ID=57597V2071141221367668675*-ID=57597V262142882225588671*ST-1332CCID=50116V272142882225588671*ST-1332CCID=50116V2886180470127255438673*-ID=50118V3744422210318676*-ID=56169C2861447047029358668*-ID=56179C486202922612729238669*-ID=56180V786202922812729238672*-ID=56183V886202922612729238669*-ID=56180V252348292264852518674*-ID=50120V333747629228682255428409*-ID=56166V347174646943358410*-ID=56167V477174646943358410*-ID=56167
Note: The bolded portion are the new allele value;* New sequence type;- No corresponding clones; ID Website:https://pubmlst.org/bigsdb?db=pubmlst_campylobacter_isolates&page=query
2.2.248株分離株所屬不同序列型和克隆系的數量 經序列比對后,48株分離株共含有24種不同的序列型,分屬于9種克隆系和11種獨特型。其中11種獨特型共包含21株分離株,占總分離株的43.75%。CC-21、CC-353和CC-1332均包含2種以上序列型,分別占菌株總數的12.5%、8.33%、6.25%,其余克隆系只包含一種序列型。所有的不同序列型中,獨特型ST-8675包含的菌株數最多,共包含8株分離株,占總菌株數的16.67%,ST-760、ST-464、ST-161和ST-354均包含3株以上的分離株,其余序列型只包含1-2株分離株,具體信息(表2)。
表2 48株分離株所屬不同序列型和克隆系的數量
Tab.2 Different STs and clonal complex contain 48 isolates
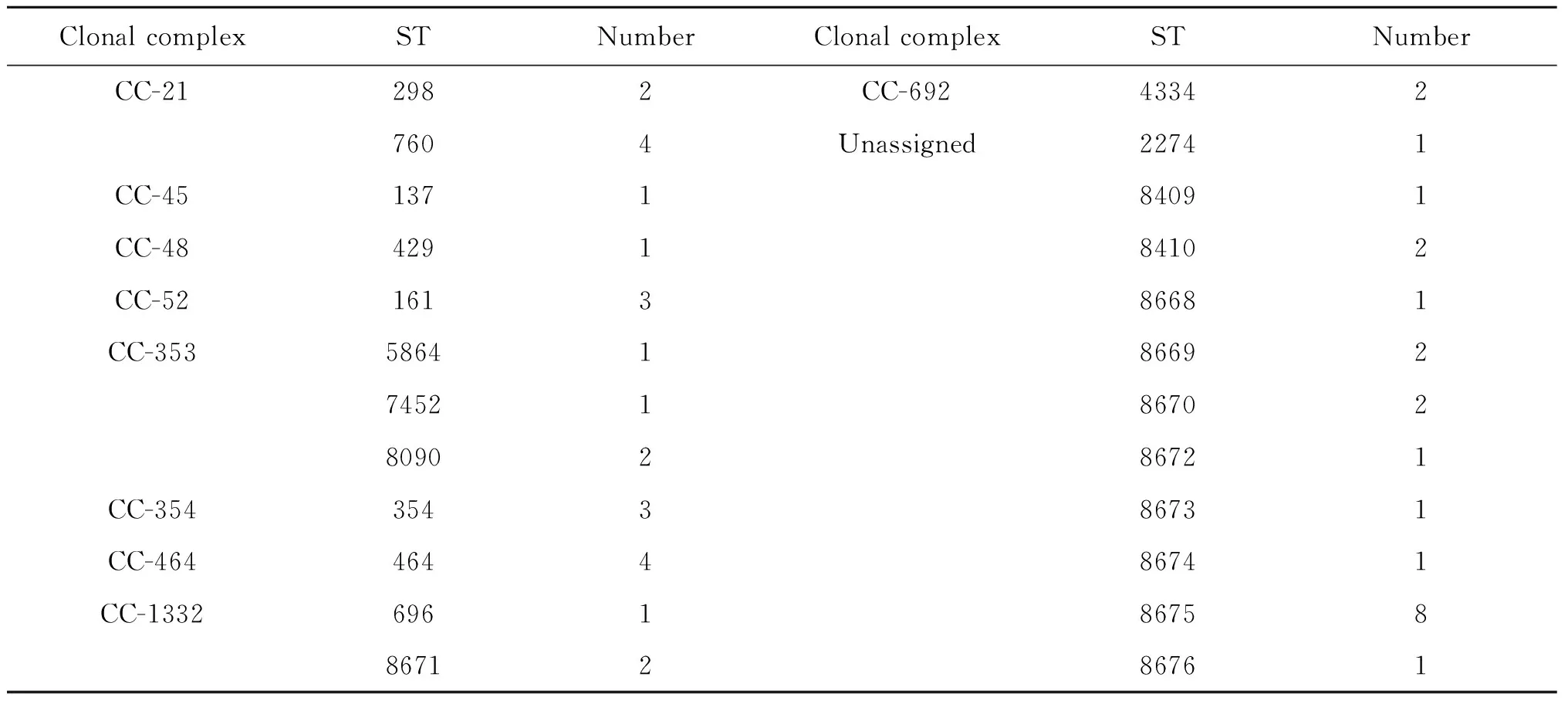
Clonal complexSTNumberClonal complexSTNumberCC-212982CC-692433427604Unassigned22741CC-45137184091CC-48429184102CC-52161386681CC-353586418669274521867028090286721CC-354354386731CC-464464486741CC-13326961867588671286761
2.348分離株MLST結果統計與分析 將分離株等位基因擴增結果統計后發現48株分離株中,等位基因值的數量最多的是pgm,為14個;其次glnA,為12個;gltA、tkt、uncA、aspA均為9個;最少的glyA,為8個。等位基因值數量的多少在某種程度上反應了該位點基因的多樣性,依此或可推知pgm基因最不穩定,突變最多,而glyA基因較穩定。依據本研究中出現的所有等位基因數值及其各數值所對應菌株的數量,結果顯示,glyA基因值為2的細菌數量最多,為21株;uncA基因值為6的細菌數量次之,為17株;而出現的各管家基因值為1的菌株分布最為廣泛。
2.4不同克隆系、序列型包含的不同地區來源菌株的數量 將不同克隆系、序列型包含的不同地區來源的菌株的數量進行統計后,結果顯示,源于(金堂養雞場)的9株分離株率屬于6種序列型;源于(溫江活禽市場2、3)地區的分離株均有8株率屬于6種序列型;源于(眉山鵪鶉養殖場)的8株分離株均歸屬于ST-8675序列型,而源于(溫江養雞場2)的分離株僅有2株率屬于2種序列型。針對克隆系而言,以CC-21包含的分離株最多為6株,以CC-45包含的分離株最少為1株。除此之外可發現同一個序列型包含2株及以上菌株的,菌株多為同一地區分離株,如ST-298包含2株(溫江活禽市場1)的分離株,ST-760包含3株(溫江養雞場1)的分離株,表明菌株的流行與地域特征有關。
2.5分離株的遺傳進化分析 使用MEGA6.0軟件對48株分離株所含的24種序列型及國內2株腹瀉病人空腸彎曲菌分離株、2株雞源分離株、1株鴨源分離株,以及英國、丹麥、波蘭、新西蘭、阿根廷、韓國、泰國、肯尼亞、印度雞源分離株各1株,共38株空腸彎曲菌的序列型進行遺傳進化樹的構建(圖2)。結果顯示,泰國和丹麥的ST-8659和ST-8661型別分離株與其余36個型別分離株處于不同分支,遺傳距離較遠;眉山鵪鶉源的ST-8675型的8株分離株(V13-20)與其同屬于一大分支的其余35個型別分離株遺傳距離較遠;屬于同一克隆系的2種型別菌株遺傳距離近,如同屬于CC-21克隆系的ST-298與ST-760處于同一小分支、屬于CC-1332克隆系的ST-8671與ST-696處于同一小分支;來源于同一地區不同場的分離株遺傳距離較近,如來源于(溫江活禽市場1)ST-5864序列型的V26分離株和(溫江養雞場2、活禽市場3)ST-8090序列型的V40、V43分離株處于同一小分支,親緣關系較近。除此之外,本研究中序列型為ST-354的V24、V42、V46分離株與英國ST-354序列型分離株處于同一小分支并與印度ST-8657序列型菌株處于同一分支,暗示著不同克隆系間的序列也存在密切相關性;國內腹瀉病人ST-8727序列型分離株與本研究中雞源分離株C1親緣關系最近,或可推之雞與人類感染有重要關系。

圖2 本研究24種序列型和國內外14株分離株序列型的遺傳進化樹Fig.2 Phylogenetic tree of 24 kinds of sequence types in this study and 14 sequence types in home and abroad
3 討 論
MLST作為一種常用且準確的分子分型方法,具有分辨率高、可積累、重復性強等優勢得到了研究者的廣泛關注[9-10]。本研究對四川部分地區分離的48株禽源空腸彎曲菌進行MLST分型,共獲得24種不同的序列型,其中有22株分離株屬于11種新序列型,新序列型占比為45.83%,發現的新序列型數量所占比率與薛峰[11]、董俊[12]等從禽類腸道分離的菌株、王新等[13]從禽肉分離的菌株以及Petra等[14]自肉雞群分離的菌株新序列型所占比率均有差異,或與不同地區分離株的遺傳進化有關。在發現的新序列型中,眉山鵪鶉源的8株分離株均為新序列型且變異度較小,或與菌株間的基因水平轉移、代謝類型相似等因素有關,是否如此機制還有待于進一步的研究證實后以便為更好地揭示空腸彎曲菌致病性與分子分型之間的相關性。
空腸彎曲菌可以在多種不同宿主間進行傳播,但某一類宿主所攜帶的空腸彎曲菌則大多歸為某一序列型或克隆系,這可能與生態適應、地理隔離、宿主相關等因素均有關[15]。本研究中的不同源分離株均存在獨特型序列,不同源分離株中的獨特型序列可反映出該源分離株基因間的多樣性。對分離株所含的序列型進行遺傳進化分析發現,鵪鶉源分離株V13-20與鴨源、雞源分離株的親緣關系較遠,表明不同家禽中的空腸彎曲菌序列型存在差異具有多樣性,分離株中出現新序列型或與空腸彎曲菌MLST數據庫中的數據量缺乏有關;分離株所含的CC-21、CC-45、CC-353、CC-354克隆系與Ragimbeau等[16]自腹瀉病人中分離到的空腸彎曲菌所含的克隆系有交叉重疊,暗示著禽類對于人類致病有潛在相關性。綜上所述,本研究結果為MLST數據庫的擴充及基于該方法探究不同地域、宿主來源的空腸彎曲菌致病性與分子分型的相關性提供了參考依據。
利益沖突:無
