隆頭魚(yú)科分類學(xué)研究進(jìn)展
2019-03-08 02:46:26黃新春唐文喬
海洋漁業(yè) 2019年1期
劉 東,黃新春,唐文喬
(1.海洋動(dòng)物系統(tǒng)分類與進(jìn)化上海高校重點(diǎn)實(shí)驗(yàn)室,上海 201306; 2.水產(chǎn)種質(zhì)資源發(fā)掘與利用教育部重點(diǎn)實(shí)驗(yàn)室,上海 201306; 3.上海海洋大學(xué),水產(chǎn)科學(xué)國(guó)家級(jí)實(shí)驗(yàn)教學(xué)示范中心,上海 201306)
隆頭魚(yú)科魚(yú)類約5 000萬(wàn)年前開(kāi)始生活在海洋中,目前已知該科魚(yú)類約71屬519余種,是海洋魚(yú)類中物種數(shù)量第2大類群,也是體形最為分化的一個(gè)類群,體長(zhǎng)最小僅幾厘米,最大可達(dá)2.3 m[1]。該類群對(duì)維持海洋珊瑚礁生態(tài)系統(tǒng)的穩(wěn)定具有重要作用。
LINNAEUS[2]于1758年在《自然系統(tǒng)》中創(chuàng)立了隆頭魚(yú)屬(Labrus)。DUMéRIL[3]依據(jù)物種命名詞性規(guī)則,把隆頭魚(yú)屬的拼寫(xiě)改為“Labre”,建立了Leiopomes科。RISSO[4]于1810年在LINNAEUS的分類系統(tǒng)基礎(chǔ)上,以隆頭魚(yú)屬為模式屬建立了隆頭魚(yú)科Labroidae,CUVIER等[5]采用了RISSO的分類體系。GUNTHER[6]于1861年最早正式確定隆頭魚(yú)科作為分類單元。自此,隆頭魚(yú)科被廣泛接受,或被寫(xiě)成“Labridae”。
隆頭魚(yú)科的研究雖然已歷經(jīng)260余年,但有關(guān)它的分類學(xué)知識(shí),包括物種鑒定、系統(tǒng)發(fā)育關(guān)系等仍然十分有限。本文梳理了隆頭魚(yú)科的分類學(xué)研究歷史、系統(tǒng)發(fā)育學(xué)研究進(jìn)展以及我國(guó)隆頭魚(yú)科的分類現(xiàn)狀,并在此基礎(chǔ)上展望了隆頭魚(yú)科研究方向,以期為隆頭魚(yú)科魚(yú)類的種質(zhì)資源保護(hù)和利用提供基礎(chǔ)數(shù)據(jù)。
1 分類學(xué)地位
1.1 隆頭魚(yú)科分類學(xué)地位
早期分類研究中,GUNTHER[7]依據(jù)下咽骨和咽齒的特征,把鸚嘴魚(yú)科(Scaridae)和巖鱚科(Odacidae)歸并至隆頭魚(yú)科,但BLEEKER[8]認(rèn)為鸚嘴魚(yú)科是一個(gè)有效類群。GILL[9]根據(jù)左、右下咽骨愈合這一特征(圖1-A),建立了咽顎區(qū)(pharyngognathi)及棘鰭上目(acanthopterygii)的分類階元,棘鰭上目包括現(xiàn)生的頜針魚(yú)目(Beloniforms)、麗魚(yú)科(Cichlidae)、雀鯛科(Pomacentridae)、隆頭魚(yú)科、鸚嘴魚(yú)科等5類群。JORDAN[10-11]研究發(fā)現(xiàn),熱帶水域隆頭魚(yú)科種類的脊椎骨數(shù)(25)較少,而溫帶水域種類的脊椎骨數(shù)(27~41)較多,因此把隆頭魚(yú)科劃分為2個(gè)地理類群。同時(shí)他認(rèn)為僅分布于澳大利亞和新西蘭的巖鱚科在頜骨形態(tài)方面與隆頭魚(yú)科存在差異,應(yīng)為一個(gè)有效類群。NORMAN[12]在1966年將鸚嘴魚(yú)科、巖鱚科和隆頭魚(yú)科組成隆頭魚(yú)目(Labriformes);隨后,GREENWOOD等[13]把NORMAN的隆頭魚(yú)目降級(jí)為隆頭魚(yú)亞目(Labroidei)。尼科里斯基[14]在《分門魚(yú)類學(xué)》中把隆頭魚(yú)作為鱸形目(Perciformes)鱸亞目(Percoidei)隆頭魚(yú)科。NELSON[15]在1976年的《世界魚(yú)類》(Fishes of the world)中將鸚嘴魚(yú)科、巖鱚科和隆頭魚(yú)科共同形成隆頭魚(yú)亞目。自隆頭魚(yú)科確立以來(lái),之所以與鸚嘴魚(yú)科和巖鱚科歸并的爭(zhēng)議持續(xù)不斷,關(guān)鍵是因?yàn)闆](méi)有發(fā)現(xiàn)類群獨(dú)有特征[16-17]。咽顎魚(yú)類特有的下頜提肌直接與腦顱相連的特征,也曾被作為隆頭魚(yú)科特有的共源性狀[18],由此與鰭棘魚(yú)類相區(qū)分,但這種特征也存在于海鯽科(Embiotocidae)、麗魚(yú)科和鸚嘴魚(yú)科中(圖1-B和圖1-C)[19]。
雖然鸚嘴魚(yú)科、巖鱚科和隆頭魚(yú)科存在形態(tài)差異,但早期基于形態(tài)學(xué)特征的系統(tǒng)學(xué)研究支持3科合并為單系群[20],或鸚嘴魚(yú)科及巖鱚科各為單系群[17,21]。WILEY等[22]在其2010年的分類系統(tǒng)中,將隆頭魚(yú)目分為6科,即隆頭魚(yú)科、鸚嘴魚(yú)科、巖鱚科、海鯽科、麗魚(yú)科和雀鯛科。WAINWRIGHT等[19]利用核基因標(biāo)記結(jié)合咽骨功能形態(tài)學(xué)分析,未將海鯽科、麗魚(yú)科和雀鯛科歸為隆頭魚(yú)目分支,而是與其它40科共同形成一個(gè)新的類群,即絨毛卵亞系,但該亞系缺乏形態(tài)特征共性支持。BETANCURR等[23]利用基因標(biāo)記(線粒體基因和核基因)進(jìn)行硬骨魚(yú)類的分子系統(tǒng)發(fā)育的研究,發(fā)現(xiàn)鼬鳚目、蟾魚(yú)目、蝦虎魚(yú)超目、海龍目、鯖目、攀鱸亞系、鲹亞系、絨毛卵亞系和鱸形魚(yú)超目共同構(gòu)成了鱸形魚(yú)系,而隆頭魚(yú)目成為鱸形魚(yú)超目的分支群,與擬鲉鱸科互為姐妹群(圖2)。

圖1 隆頭魚(yú)的咽骨與一般鰭棘魚(yú)類的比較Fig.1 Comparative diagram of the pharyngeal jaw bones between wrasses and generalized spiny-rayed fishes注:A:隆頭魚(yú)咽骨圖;B:一般鰭棘魚(yú)類的咽頜后部觀圖;C:咽顎魚(yú)類的咽頜后部觀圖。MS-提肌;UPJ-上咽骨;LPJ-下咽骨;AD4-第四鰓收肌;LE4-第四外提肌;EB-上鰓骨;NC-腦顱。分圖B、C引自WAINWRIGHT等[19]Notes: A: Diagram of the pharngeal jaw bones in wrasses; B: Diagram of the posterior view of the pharngeal jaw apparatus in generalized spiny-rayed fishes; C: Diagram of the posterior view of the pharyngognathous pharyngeal jaw. MS: muscular sling; UPJ: upper pharngeal jaw; LPJ: lower pharngeal jaw; AD4: fourth branchial adductor muscle; LE4: fourth levator externus muscle; EB: epibranchial bone; NC: neurocranium; B and C are cited from WAINWRIGHT et al[19]

圖2 隆頭魚(yú)目在鱸形魚(yú)系中的進(jìn)化關(guān)系Fig.2 Phylogenetic relationship of Labriformesin Percomorphaceae
現(xiàn)有分子系統(tǒng)支持鸚嘴魚(yú)科、巖鱚科和隆頭魚(yú)科為共同單系起源[19],但這一類群缺乏鱸形目魚(yú)類普遍存在的咽咬式攝食機(jī)制,即上頜第4上鰓骨向下擠壓下咽骨咬碎食物(圖1-B)。隆頭魚(yú)則是單一的下咽骨兩端由提肌與腦顱相連,提肌收縮向上擠壓上咽骨破碎食物(圖1-C)。這種攝食機(jī)制能增強(qiáng)捕食能力及擴(kuò)大食物來(lái)源,攝食適應(yīng)性的進(jìn)化促進(jìn)了物種的多樣性。
NELSON等[6]在2016年的《世界魚(yú)類》(第五版)(Fishes of the world)(5th Edition)中將隆頭魚(yú)亞目提升為隆頭魚(yú)目。他們根據(jù)咽喉部與咽骨的共源特征,例如單一的下咽骨以及上咽骨通過(guò)關(guān)節(jié)與頭蓋骨相連等特征,認(rèn)為隆頭魚(yú)目是單一系群,將該目分為隆頭魚(yú)科、鸚嘴魚(yú)科和巖鱚科。
1.2 隆頭魚(yú)科分類和系統(tǒng)發(fā)育關(guān)系
GUNTHER[5]在1861年正式確定隆頭魚(yú)科時(shí)包括6個(gè)亞科群:Labrina、Hypsigenina、Julidina、Pseudodacina、Scarina和Odacina,鸚嘴魚(yú)科和巖鱚科分別作為隆頭魚(yú)科的亞科群。隨后,GUNTHER[1]使用Choeropina代替了Hypsigenyini,隆頭魚(yú)科包含6群46屬,其中Labrichthys、Pseudolabrus、Anampses、Gomphosus和Coris等5屬隸屬于Julidina群。同一時(shí)期,BLEEKER[8]提出不同的分類系統(tǒng),隆頭魚(yú)科(未含鸚嘴魚(yú)科)劃分為10群:Cheilinina、Pseudodacina、Cheilionina、 Pseudolabrina、Novaculaena、Labrichthyna、Cossyphina、Labrina、Odacina和Clepticina。
GILL[24]將隆頭魚(yú)科新建1亞科Xyrichthyinae,并把GüNTHER分類中的Julidina群內(nèi)4屬提升為亞科,結(jié)合BLEEKER的群,將隆頭魚(yú)科劃分為11個(gè)亞科,即Labrinae、 Cheilioninae、Choeropinae、Xyrichthyinae、Coridinae、Gomphosinae、Anampsinae、 Cheilininae、Pseudolabrinae、Clepticinae及Pseudodacinae。JORDAN等[25]于1902年新建了普提魚(yú)亞科Bodianinae和錦魚(yú)亞科Thalassominae。 REGAN[26]于1913年建立9個(gè)亞科,即Julidinae、Xyrichthyinae、Cheilininae、Epibulinae、Clepticinae、Harpinae、 Pseudodacinae、Labrinae及Malacopterinae,并把Pseudolabrus歸入Julidinae亞科。NORMAN[12]在GüNTHER分類群基礎(chǔ)上,把隆頭魚(yú)科劃分為9亞科,即Hemipteronotinae、Epibulinae、 Clepticinae、Pseudodacinae、Neolabrinae、Cheilininae、Julidinae、Labrinae及Bodianinae,使用普提魚(yú)亞科替代了GüNTHER分類中的亞科群Choeropina。NELSON[15]把鸚嘴魚(yú)科和巖鱚科從隆頭魚(yú)科分離之后,包括幾個(gè)亞科和58屬。RUSSELL[27]通過(guò)形態(tài)特征,將Austrolabrus、Dotalabrus、Eupetrichthys、Notolabrus、Pictilabrus及Pseudolabrus等6屬組成Pseudolabrini族,與Julidini族互為姐妹群,隸屬于NORMAN分類中的Julidinae亞科。
GOMON[16]把REGAN和NORMAN分類中的亞科降為族,重新使用GüNTHER分類中的“Hypsigenyini”族名,由于未有亞科水平上聚類數(shù)據(jù)支持,因此把隆頭魚(yú)科劃分為10族,即Hypsigenyini、Scarini、Labrini、Pseudocheilini、Cheilini、 Odacini、Pseudolabrini、Labrichthyini、Julidini及Novaculini,鸚嘴魚(yú)科與隆頭魚(yú)科Hypsigenyini族的Pseudodax屬互為姐妹群。 WESTNEAT[28]認(rèn)為Cheilinini族為單系群,包括Cheilinus、Doratonotus、Epibulus、Oxycheilinus及Wetmorella等5屬,其與Pseudocheilini族互為姐妹群。CLEMENTS等[17]認(rèn)為巖鱚族Haletta、Neoodax、Odax及Siphonognathus等4屬形成的單系群應(yīng)歸于Hypsigenyini族,而歸于巖鱚科的Cheilio屬[16,27]并未與該科其它屬互為姐妹群,也不聚于Hypsigenyini族,其分類地位有待商榷[29]。
WESTNEAT等[29]利用線粒體和核基因標(biāo)記,構(gòu)建了隆頭魚(yú)科的進(jìn)化拓?fù)浣Y(jié)構(gòu),形成幾個(gè)主要族(圖3),其中普提魚(yú)族居于樹(shù)根,與隆頭魚(yú)科其它族互為姐妹群;唇魚(yú)族和鸚嘴魚(yú)族互為姐妹群而與隆頭魚(yú)族親緣關(guān)系最近;離鰭魚(yú)族和擬唇魚(yú)族互為姐妹群;盔魚(yú)族是包括突唇魚(yú)族和擬隆頭魚(yú)族在內(nèi)最為分化的一群;盔魚(yú)族中幾個(gè)大屬?zèng)]有分別聚群成單系群,表明了隆頭魚(yú)頜骨的攝食功能差異,導(dǎo)致屬內(nèi)種間分化和族內(nèi)屬間聚集的結(jié)果。頜骨攝食功能形態(tài)學(xué)的演化是隆頭魚(yú)科進(jìn)化的典型特征[30]。

圖3 基于分子標(biāo)記的隆頭魚(yú)各族的系統(tǒng)演化Fig.3 Phylogenetic relationship of tribes in Labridae based on molecular markers
2 隆頭魚(yú)科分類特征和主要屬分類研究
2.1 隆頭魚(yú)科分類特征
隆頭魚(yú)科多數(shù)種類嘴唇肥厚,最初稱之為唇魚(yú)(lip-fish)。體形側(cè)偏,具有單個(gè)背鰭,鰭棘和鰭條部連續(xù),體被圓形鱗。側(cè)線鱗一般25~80片,少數(shù)種類超過(guò)100片,側(cè)線分3種類型:1)連續(xù)型,側(cè)線連續(xù)、平緩向后伸至尾鰭基部;2)彎折型,側(cè)線向后延伸至背鰭后部下方時(shí),急劇向下彎折,然后水平伸至尾鰭基部;3)中斷型,側(cè)線在背鰭后部下方中斷,分為前后兩部分(圖4)。口前位,能向前方伸出。兩頜齒分離或在基部愈合成骨脊,前方數(shù)齒多呈犬齒狀。犁骨與腭骨上無(wú)齒。左右下咽骨完全愈合成一體,多數(shù)呈T或Y形。咽骨齒呈錐形、豆粒形或臼齒形。唇厚,內(nèi)側(cè)有縱褶。頭部每側(cè)有2鼻孔。鰓蓋膜與峽部相連或不相連。背鰭棘8~15根,鰭條6~21根;臀鰭棘2~3根,鰭條7~18根;脊椎骨23~42塊。因?yàn)榉N類不一,體型差異很大。大多數(shù)種體色艷麗,但不同發(fā)育時(shí)期及雌雄之間或有差異。
2.2 主要屬分類研究
NELSON等[6]在《世界魚(yú)類》(第五版)(Fishes of the world)(5th Edition)中列出的隆頭魚(yú)科71屬中,具單種的屬有29個(gè),約50%的種類集中在海豬魚(yú)屬(79種)、絲隆頭魚(yú)屬(48種)、普提魚(yú)屬(43種)、錦魚(yú)屬(28種)、盔魚(yú)屬(27種)、頸鰭魚(yú)屬(21種)和副唇魚(yú)屬(17種)等7屬。本文就分類較為復(fù)雜的屬進(jìn)行分析。
以雙斑海豬魚(yú)(HalichoeresbimaculatusRüppell)為模式種建立的海豬魚(yú)屬,RANDALL等[31]認(rèn)為該屬的種類、雌雄差異以及幼魚(yú)期的不同體色均會(huì)造成物種鑒定困難。海豬魚(yú)物種豐富,廣泛分布于印度-太平洋-加勒比海,為熱帶珊瑚礁海洋生物多樣性的重要組成物種。以線粒體基因分子標(biāo)記的系統(tǒng)發(fā)育研究表明,海豬魚(yú)屬為復(fù)系,具有多個(gè)起源中心[32]。有學(xué)者提議海豬魚(yú)屬的分子系統(tǒng)各分支群應(yīng)確立為新屬,但缺乏新屬鑒定的外部形態(tài)特征[33]。盡管海豬魚(yú)屬包括了近80個(gè)物種,分子系統(tǒng)研究表明有幾個(gè)明確的譜系,但多數(shù)學(xué)者傾向于維持該屬的完整性。

圖4 隆頭魚(yú)背鰭后下方側(cè)線形態(tài)Fig.4 Patterns of lateral lines situated below posterior portion of the soft dorsal fin in wrasses注:A:連續(xù)型;B:彎折型;C:中斷型Note:A: continuous pattern; B: abruptly curved pattern; C: discontinuous pattern
以紅普提魚(yú)(BodianusbodianusBloch)為模式種建立的普提魚(yú)屬,分布于大西洋、印度洋和太平洋的熱帶及溫?zé)釒ШS颉OMON[34]利用支序系統(tǒng)學(xué)的方法,結(jié)合外部形態(tài)和內(nèi)部解剖結(jié)構(gòu)特征,構(gòu)建了普提魚(yú)屬的系統(tǒng)發(fā)育,支持該屬劃分為10亞屬,但有幾點(diǎn)缺陷:一是亞屬的種類分布太廣,超出傳統(tǒng)的物種地理分布格局;二是亞屬的鑒別特征單一,多數(shù)僅具1~2個(gè)鑒別性狀,實(shí)際操作困難。這種劃分沒(méi)有被多數(shù)學(xué)者采用[6,33]。SANTINI等[35]使用線粒體基因和核基因標(biāo)記的研究支持該屬為復(fù)系群,表明因?yàn)閿z食分化導(dǎo)致該屬物種約在早中新世時(shí)期的二千萬(wàn)年開(kāi)始了擴(kuò)張,起點(diǎn)為印度-西太平洋和南-西太平洋。
以紫錦魚(yú)(ScaruspurpureusForssk?l)為模式種建立的錦魚(yú)屬,是一類中小型魚(yú)類,體長(zhǎng)一般13~46 cm,廣泛分布于世界大洋亞熱帶珊瑚叢和礁巖環(huán)境,約為28種[6]。依據(jù)分子系統(tǒng)發(fā)育[36],該屬物種形成2個(gè)分支,即大西洋分支和印度-太平洋分支。分支形成的歷史因素,歸結(jié)于古大西洋地區(qū)終極特提斯隆起和巴拿馬海峽閉合形成的地理障礙。尖嘴魚(yú)屬(Gomphosus)的2個(gè)物種雜色尖嘴魚(yú)(G.varius)和雀尖嘴魚(yú)(G.caerulaeus)為錦魚(yú)屬的印度-太平洋分支,表明這兩屬起源于同一祖先,隨后因食性差異導(dǎo)致攝食器官分化:尖嘴魚(yú)屬吻延長(zhǎng)呈管狀,錦魚(yú)屬吻正常,但屬間兩種雜色尖嘴魚(yú)和新月錦魚(yú)(Thalassomalunare)會(huì)形成自然雜交種,盡管如此,尖嘴魚(yú)屬仍為有效屬[33]。
頸鰭魚(yú)屬(Iniistius)的分類歷史最為復(fù)雜,該屬物種早期多歸入離鰭魚(yú)屬(HemipteronotusLacepède),以Naucratesdoctor(Linnaeus 1758)為模式種建立的離鰭魚(yú)屬的目的是歸類短背鰭物種[37]。以CoryphaenanovaculaLinnaeus為模式種建立的連鰭唇魚(yú)屬(XyrichtysCuvier),隨后被取消,部分種歸入離鰭魚(yú)屬。GILL[9]于1872年以孔雀連鰭唇魚(yú)(XyrichtyspavoValenciennes)為模式種建立了頸鰭魚(yú)屬。JORDAN[10]把離鰭魚(yú)屬、連鰭唇魚(yú)屬和頸鰭魚(yú)屬歸入盔魚(yú)科Coridae。后來(lái)研究發(fā)現(xiàn)離鰭魚(yú)屬的模式種隸屬于鲹科Carangidae 而非隆頭魚(yú)科,由此該屬失效。FOWLER[38]于1957年以鰓斑離鰭魚(yú)(HemipteronotusevidesJordan & Richardson)為模式種建立了Duohemipteronotus,但PARENTI等[33]認(rèn)為該屬與連鰭唇魚(yú)屬同名。RANDALL[37]最初把頸鰭魚(yú)屬作為連鰭唇魚(yú)屬的同名,但由于發(fā)現(xiàn)這兩屬的鄂骨、翼狀骨及額骨等具形態(tài)差異,由此確定這為2個(gè)屬有效,可根據(jù)背鰭起點(diǎn)和第1、2背鰭棘距離區(qū)分,而連鰭唇魚(yú)屬的物種在中國(guó)海域沒(méi)有分布。
3 隆頭魚(yú)科物種生態(tài)習(xí)性
隆頭魚(yú)主要生活在熱帶和溫帶海域近海岸至200 m深的珊瑚礁、巖礁、海草、海藻和泥沙等環(huán)境中,形成3類捕食方式:1)珊瑚礁類捕食,以螺、蚌、貝、小魚(yú)和小蝦為食,用咽喉齒咬碎貝殼;2)濾食性捕食,包括浮游生物和海藻;3)高度特化的捕食,或如小型的裂唇魚(yú)(Labroides)會(huì)充當(dāng)大型魚(yú)的清潔工,以其它魚(yú)體表或口腔內(nèi)附著物為食,或如伸口魚(yú)(Epibulusinsidiator)的長(zhǎng)吻藏于口內(nèi),捕食時(shí)從口中伸出,可達(dá)0.7倍于頭長(zhǎng),依靠長(zhǎng)吻捕獲獵物。
隆頭魚(yú)多數(shù)種類的體色需經(jīng)歷幼魚(yú)期、兩性中雌性先熟的成熟期及雌性性逆轉(zhuǎn)的超雄魚(yú)期等3個(gè)發(fā)育時(shí)期的變化。在幼魚(yú)階段,魚(yú)類體色十分絢麗,因年齡和雌雄的差異,體色和色斑也有不同,如盔魚(yú)屬、尖嘴魚(yú)屬和海豬魚(yú)屬的種類因其體色艷麗或體形怪異而為著名的觀賞魚(yú);多數(shù)種類具有雄性異型、雌性先熟的特性。少數(shù)種類初次性成熟的雄性個(gè)體小,小雄魚(yú)以群體形式排出精液與雌魚(yú)的卵子受精。或雌魚(yú)性成熟后逆轉(zhuǎn)而成的超雄魚(yú),個(gè)體較大,超雄魚(yú)實(shí)行一對(duì)一的繁殖方式與雌魚(yú)進(jìn)行交配,產(chǎn)卵受精后,隨后超雄魚(yú)保護(hù)產(chǎn)卵場(chǎng)[39]。
4 中國(guó)隆頭魚(yú)科分類研究
中國(guó)隆頭魚(yú)的分類研究,最早可追溯到博物學(xué)家OSBECK[40]在1750—1752年間在中國(guó)旅行中記載的2種隆頭魚(yú):Labrusopercularis和L.Chinensis。1846年,RICHARDSONS報(bào)道了廣東沿海隆頭魚(yú)1新種,即遠(yuǎn)東隆頭魚(yú)(Pseudolabruseoethinus)[41]。1931年,朱元鼎[42]發(fā)表的我國(guó)第一部《中國(guó)魚(yú)類索引》收錄了2種隆頭魚(yú)。20世紀(jì)50年代中期陸續(xù)開(kāi)展的海域區(qū)系普查,促進(jìn)了我國(guó)隆頭魚(yú)分類的系統(tǒng)研究。
《南海魚(yú)類志》[43]、《南海諸島海域魚(yú)類志》[44]及《北部灣魚(yú)類圖鑒》[45]等記錄南海隆頭魚(yú)科28屬76種;《中國(guó)南海西南中沙群島珊瑚礁魚(yú)類圖譜》[46]中記錄19屬54種,經(jīng)統(tǒng)計(jì),南海隆頭魚(yú)科達(dá)29屬99種。《東海魚(yú)類志》[47]、《福建魚(yú)類志》[48]及《浙江海洋魚(yú)類志》[49]等記錄東海有10屬18種,黃海和渤海未調(diào)查到[50]。20世紀(jì)80年代中期,《中國(guó)魚(yú)類系統(tǒng)檢索》[51]記錄中國(guó)隆頭魚(yú)科29屬99種。中國(guó)臺(tái)灣學(xué)者對(duì)隆頭魚(yú)的分類研究較為深入,于名振[52]在1968年報(bào)道了臺(tái)灣海域隆頭魚(yú)科3亞科22屬85種,沈世杰[53]進(jìn)一步完善了臺(tái)灣海域隆頭魚(yú)科的分類研究,回顧了臺(tái)灣海域絲隆頭魚(yú)屬的分類[54],隨后編著的《臺(tái)灣近海魚(yú)類圖鑒》[55]、《臺(tái)灣魚(yú)類志》[56]和《臺(tái)灣魚(yú)類圖鑒》[57]等記錄臺(tái)灣海域隆頭魚(yú)科為32屬130種。劉靜在劉瑞玉編《中國(guó)海洋生物名錄》[58]列出中國(guó)隆頭魚(yú)科有40屬166種,伍漢霖在黃宗國(guó)、林茂編著的《中國(guó)海洋物種多樣性》[59]所列名錄為35屬157種。
對(duì)于中國(guó)隆頭魚(yú)科的科下分類,最早FOWLER[38]依據(jù)側(cè)線的形態(tài)和背鰭棘數(shù)把隆頭魚(yú)科(不包括鸚嘴魚(yú)科)分為3亞科(普提魚(yú)亞科、盔魚(yú)亞科、唇魚(yú)亞科),14族(Choerodontidi、Bodianidi、Epibulidi、Duymaeriidi、Labroididi、Thalassomidi、Gomphosidi、Coridi、Hemigymnidi、Pseudolabridi、Cheiliidi、Anampsidi、 Cheilinidi、Hemipteronotidi),30屬。FOWLER的工作奠定了中國(guó)隆頭魚(yú)科的分類基礎(chǔ)。SHEN等[53]在FOWLER的基礎(chǔ)上增加了擬巖喜亞科,取消族的階元;成慶泰等[51]結(jié)合FOWLER[38]和JORDAN等[25]的分類成果,劃分中國(guó)隆頭魚(yú)科為4亞科(普提魚(yú)亞科、盔魚(yú)亞科、唇魚(yú)亞科、錦魚(yú)亞科),29屬。于名振[60]在2001年取消亞科階元,分為7族(隆頭魚(yú)族、普提魚(yú)族、擬巖喜族、盔魚(yú)族、突唇魚(yú)族、唇魚(yú)族、離鰭魚(yú)族),64屬。在整理我國(guó)主要館藏的隆頭魚(yú)標(biāo)本后,依據(jù)MIKAMI[61]于2013年的最新研究成果,筆者以側(cè)線形態(tài)(圖4)為分類特征,將中國(guó)隆頭魚(yú)科分為普提魚(yú)亞科、唇魚(yú)亞科和盔魚(yú)亞科。
5 中國(guó)隆頭魚(yú)科物種厘定和展望
5.1 物種厘定
我國(guó)對(duì)隆頭魚(yú)科的研究起步較晚,至今仍停留在物種鑒定層面。成慶泰等[62]于1979年報(bào)道中國(guó)隆頭魚(yú)科第1個(gè)新種:尖尾新絲隆頭魚(yú)(Neocirrhilabrusoxyurus),但與RANDALL等[54]報(bào)道黑緣絲隆頭魚(yú)(C.melanomarginatus)為同物異名。CHEN等[63]于1995年發(fā)表了我國(guó)第一個(gè)產(chǎn)自臺(tái)灣的新種:側(cè)斑擬盔魚(yú)(Pseudocorisocellatus)。最近20年的研究,也僅有零星的報(bào)道新記錄種[64-66]。中國(guó)隆頭魚(yú)科魚(yú)類更缺乏分子系統(tǒng)學(xué)和動(dòng)物地理學(xué)方面的研究,這與我國(guó)具有豐富的隆頭魚(yú)科種類資源極不相稱。
參考1750—2017年期間公開(kāi)發(fā)表的文獻(xiàn)資料、Fishbase 和臺(tái)灣魚(yú)類資料庫(kù)(http:// fishdb. sinica.edu.tw)以及魚(yú)類分類數(shù)據(jù)庫(kù)(Catalog of fishes),經(jīng)同物異名和錯(cuò)誤命名的考證和修訂,厘定中國(guó)隆頭魚(yú)科(不包括鸚嘴魚(yú)科)有38屬150種(數(shù)據(jù)未發(fā)表)。中國(guó)各海域的屬種具體分布見(jiàn)表1,東海分布的隆頭魚(yú)科的屬種均與南海重疊,而臺(tái)灣的屬種的分布來(lái)源于臺(tái)灣島海域、南海太平島海域和其它島海域,臺(tái)灣分布非特指臺(tái)灣島海域。
5.2 展望
隆頭魚(yú)科自建立以來(lái),科內(nèi)屬種不斷的歸并和增加。在世界范圍內(nèi),2000年記錄454種,2010年記錄504種[33],2016年記錄519種[6],而我國(guó)已知隆頭魚(yú)科種類約占世界總數(shù)的28.9%。中國(guó)這些種類中,鑒定所依據(jù)的模式標(biāo)本記錄大多來(lái)自紅海、印度洋和南、東太平洋,與我國(guó)所處的西太平洋相隔甚遠(yuǎn),因此我國(guó)海域所產(chǎn)標(biāo)本與模式種具一定的形態(tài)差異,時(shí)有誤鑒發(fā)生。為此,需要廣泛深入的開(kāi)展我國(guó)物種普查和鑒定研究,摸清我國(guó)隆頭魚(yú)種質(zhì)資源狀況。
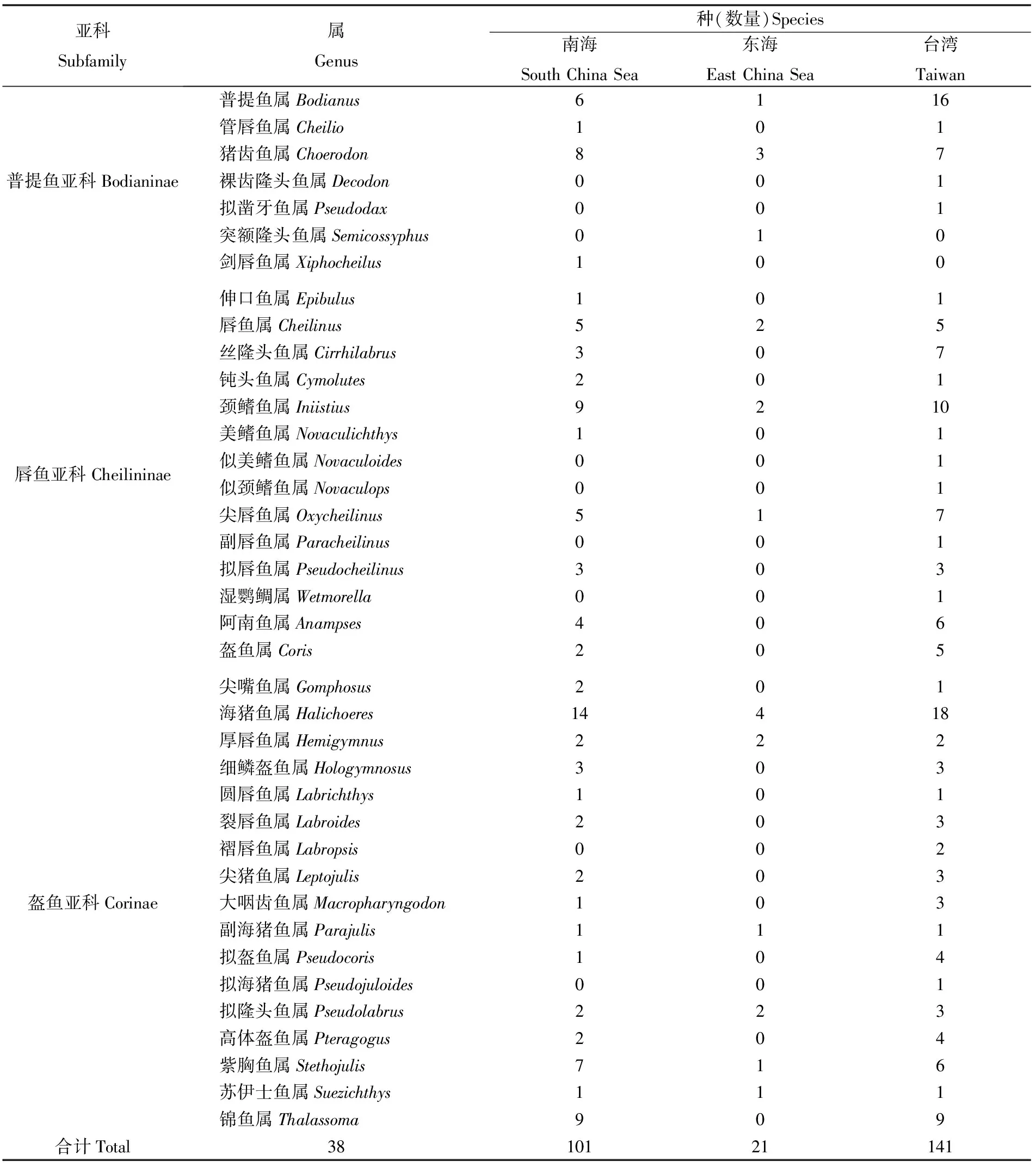
表1 中國(guó)海域隆頭魚(yú)科屬種的分布Tab.1 Distribution of species and genera of Labridae in China sea areas
物種鑒定層面上,由于隆頭魚(yú)科魚(yú)類為世界性分布,多數(shù)種存在幼魚(yú)期、成魚(yú)期及超雄期之間以及雌雄之間的體色和形態(tài)的差異,或物種之間產(chǎn)生的自然雜交種等問(wèn)題,為物種分類鑒定造成了困難。此外,分布于我國(guó)海域的標(biāo)本是否為已知種類或所具形態(tài)是否為種內(nèi)差異等問(wèn)題,有必要在收集標(biāo)本的基礎(chǔ)上,采用先進(jìn)的分子生物學(xué)技術(shù)如DNA條形碼等予以解決。
分子系統(tǒng)和親緣地理學(xué)研究方面,中國(guó)隆頭魚(yú)科的研究尚屬空白,今后可以通過(guò)線粒體基因和核基因作為分子標(biāo)記,構(gòu)建系統(tǒng)發(fā)育樹(shù),以解決中國(guó)隆頭魚(yú)科屬種進(jìn)化中的單復(fù)性問(wèn)題以及進(jìn)化關(guān)系,從而推動(dòng)隆頭魚(yú)科系統(tǒng)發(fā)育的研究。隨著測(cè)序技術(shù)的發(fā)展,測(cè)序成本進(jìn)一步降低,將加速產(chǎn)生隆頭魚(yú)科物種基因數(shù)據(jù),如果結(jié)合新方法與新工具,以大數(shù)據(jù)分析中國(guó)隆頭魚(yú)科的分布成因及其發(fā)生過(guò)程,將有助于研究世界分布的隆頭魚(yú)科不同分類階元的地理分布格局,探討隆頭魚(yú)科魚(yú)類的起源與分化,從而深入理解多樣性的物種對(duì)海洋生態(tài)穩(wěn)定的作用機(jī)制。
猜你喜歡
課堂內(nèi)外·初中版(科學(xué)少年)(2025年1期)2025-02-28 00:00:00
課堂內(nèi)外·初中版(科學(xué)少年)(2025年2期)2025-02-28 00:00:00
英語(yǔ)世界(2023年10期)2023-11-17 09:18:18
數(shù)學(xué)小靈通(1-2年級(jí))(2021年4期)2021-06-09 06:25:56
大眾健康(2021年6期)2021-06-08 19:30:06
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2019年4期)2019-05-20 10:06:32
科學(xué)大眾(中學(xué))(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2018年6期)2018-06-26 08:36:06
初中生世界·七年級(jí)(2017年9期)2017-10-13 22:27:46
