鹽脅迫對寒地粳稻籽粒淀粉形成積累及產量的影響
2019-03-08 02:00:38胡博文谷嬌嬌沙漢景張君顏黃書勤趙宏偉
華北農學報 2019年1期
關鍵詞:差異
胡博文,谷嬌嬌,賈 琰,沙漢景,張君顏,黃書勤,趙宏偉
(東北農業大學 水稻研究所,黑龍江 哈爾濱 150030)
黑龍江省是寒地粳稻的重要栽植區域,其種植總面積及產量穩步持續增長[1]。鹽脅迫對水稻有諸多影響,其中包括:造成發育延遲、抑制水稻組織與器官生長分化,最終影響產量及品質[2-4]。淀粉是水稻籽粒中的重要組成成分,其比例大多為80%~90%,因此,淀粉的形成積聚過程即為水稻籽粒的充實過程[5],而該過程受鹽脅迫的影響較大。
籽粒中的淀粉是由源器官所產生的光合產物,以蔗糖的方式運輸到籽粒后在多個酶的共同作用下轉化而成[6-7]。這一過程中,ADPG焦磷酸化酶、可溶性淀粉合成酶(SSS)以及淀粉分支酶(Q酶)發揮主要調控作用[8-9]。ADPG焦磷酸化酶的功能是產生直接前體ADPG,催化葡萄糖-1-磷酸與ATP的反應,該酶是淀粉合成過程中的限速酶[10]。在SSS的催化作用下,生成的ADPG與淀粉引物(葡聚糖)發生相互作用從而使葡萄糖分子轉運至淀粉引物,致使淀粉鏈的延伸[11]。Q酶決定淀粉組分與結構,是支鏈淀粉合成的關鍵酶[12]。
當前,國內外已有許多學者對鹽脅迫下水稻的響應機制進行了深入研究。陳海燕等[13]研究表明,在鹽脅迫下水稻根系中超氧化物歧化酶(SOD)和抗壞血酸過氧化物酶(APX)活性無明顯變化,而耐鹽品種的過氧化氫酶(CAT)和過氧化物酶(POD)活性高于鹽敏感品種。晏斌等[14]發現,在86 mmol/L的NaCl培養液中,適當增加外界 Ca2+濃度,會使水稻體內Na+濃度下降,并保證水稻植株有較高相對生長率。Wutipraditkul等[15]研究發現,施用外源物質脯氨酸可增強水稻的耐鹽性。前人多從抗氧化機制、滲透調節以及離子分布等角度解析水稻對鹽脅迫的響應機制,但鹽脅迫對灌漿結實期寒地粳稻籽粒淀粉合成關鍵酶活性、淀粉含量及最終產量形成的影響的相關研究仍缺乏系統性報道。
本試驗采用寒地粳稻品種龍稻5(耐鹽)、牡丹江30(鹽敏感)為試驗材料,研究不同濃度鹽脅迫下寒地粳稻籽粒淀粉合成關鍵酶活性變化規律及其與淀粉含量間關系,揭示寒地粳稻籽粒淀粉合成代謝對鹽脅迫的響應機制,明確鹽脅迫對寒地粳稻產量及產量構成因素的影響,以期豐富寒地粳稻耐鹽研究的生理基礎,并為水稻選擇適宜的鹽堿地種植和鹽漬土水稻栽培提供理論依據。
1 材料和方法
1.1 試驗材料
經東北農業大學水稻研究所前人篩選所得的耐鹽性不同的2個寒地粳稻品種:牡丹江30(鹽敏感)、龍稻5(耐鹽)。
1.2 試驗設計
試驗在東北農業大學盆栽場進行。于2017年4月20日播種,5月30日移栽。本試驗采用完全隨機試驗設計,選擇無滲漏的塑料盆體,土壤風干后過篩,每盆裝10 kg土壤。施鹽量設置5個梯度,以施入土壤NaCl占土壤總量的百分比表示,分別為:0,0.075%,0.150%,0.225%,0.300% 。其中,牡丹江30各處理依次記為M1(對照)、M2、M3、M4、M5,龍稻5則記為L1(對照)、L2、L3、L4、L5。共計10個處理。于返青后開始鹽處理,分5次處理完畢。盆栽密度設置為4穴/盆,每穴3株,每個處理80盆。田間管理同一般生產田。
1.3 試驗方法
1.3.1 籽粒淀粉合成代謝關鍵酶活性及淀粉的取樣方法 記錄各處理到達齊穗日期,并于齊穗期標記每處理生長整齊的植株120穴。在齊穗后的7,14,21,28,35 d的9:30-11:00(晴天)選取每處理所標記植株6穴,其中,3穴取稻穗中部籽粒用液氮冷凍處理并移至-80 ℃冰柜,用于測定籽粒的ADPG焦磷酸化酶、SSS和Q酶活性。剩余3穴籽粒用于淀粉含量的測定。
1.3.2 測定方法
1.3.2.1 籽粒淀粉合成代謝關鍵酶的測定方法 參照李太貴等[16]測定Q酶活性的方法。以吸光值下降一個百分點為一個酶活性單位U。
參照Doehlert等[17]、程方民等[18]方法,采用相應試劑盒測定ADPG焦磷酸化酶、SSS活性。酶活性測定重復3次。
1.3.2.2 籽粒淀粉及其組分含量測定 稱取各處理籽粒粉碎樣品0.1 g用于淀粉含量的測定,3次重復。采用雙波長法測定籽粒淀粉含量[19]。以每1 000 g籽粒中淀粉量的克數表示,即g/kg。
1.3.2.3 產量及產量構成因素測定 在成熟期每處理選取9穴測定有效穗數、穗粒數、千粒質量及結實率并計算理論產量。
1.4 數據分析
采用Microsoft Excel 2010和SPSS 19.0統計分析數據。
2 結果與分析
2.1 鹽脅迫下寒地粳稻籽粒淀粉組分的動態變化
2.1.1 直鏈淀粉積累動態 由圖1可知,寒地粳稻籽粒直鏈淀粉含量隨齊穗后天數的遞增呈持續增長狀態,直鏈淀粉含量的增長趨勢由快速增長趨于平緩增長。同對照相比(M1、L1),鹽處理下籽粒直鏈淀粉含量處于較高水平。牡丹江30的M1及M5表現為:齊穗后7 d差異不顯著,其余天數均表現為差異顯著;龍稻5的L1及L5表現為:齊穗后7,14,21 d差異不顯著,其余天數差異顯著。
在齊穗后的7~14 d這一快速增長階段,鹽敏感品種牡丹江30各處理增長速率為 M4>M5>M1>M2>M3,耐鹽品種龍稻5為L3>L5>L1>L4>L2,當土壤含鹽量達到0.300%時,2個品種的直鏈淀粉增長速率均高于對照水平。2個寒地粳稻品種在高鹽脅迫下的反應不同,具體表現為增長速率M4>L4、M5>L5。
齊穗后21~35 d籽粒直鏈淀粉含量增長趨于平緩。牡丹江30籽粒直鏈淀粉含量在齊穗后35 d表現為M4>M5>M3>M2>M1;龍稻5為L5>L4>L3>L2>L1。
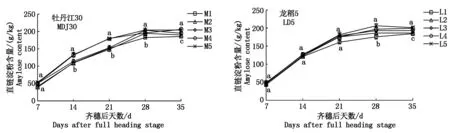
圖1 鹽脅迫下籽粒直鏈淀粉含量動態變化Fig.1 Dynamic changes of amylose content in kernels under salt stress
2.1.2 支鏈淀粉積累動態 由圖2可知,寒地粳稻籽粒支鏈淀粉的積累規律為持續增長趨勢。鹽脅迫降低寒地粳稻籽粒支鏈淀粉含量,具體表現是土壤含鹽量越高,籽粒支鏈淀粉含量越低。牡丹江30的M1及M5表現為:齊穗后7 d差異不顯著,其余天數均表現為差異顯著;龍稻5的L1及L5表現為:除齊穗后7,21 d差異不顯著外,其余天數均差異顯著。
可以發現,在齊穗后的7~21 d這一階段,2個寒地粳稻品種的各處理在積累支鏈淀粉含量的速率上表現不盡相同,其中M1、M2、L1、L2、L3處理表現為先快速增長后緩慢增長,其余處理則是先緩慢增長后快速增長。可見,當土壤含鹽量達到0.150%時,牡丹江30受影響較大,而龍稻5則是當土壤含量達到0.225%時才受較大影響。
當灌漿過程進行到齊穗后21 d后,各處理增長速率趨于平緩。對照及低濃度(0.075%)鹽處理下寒地粳稻籽粒支鏈淀粉含量一直保持相對較高水平,且差異較小。
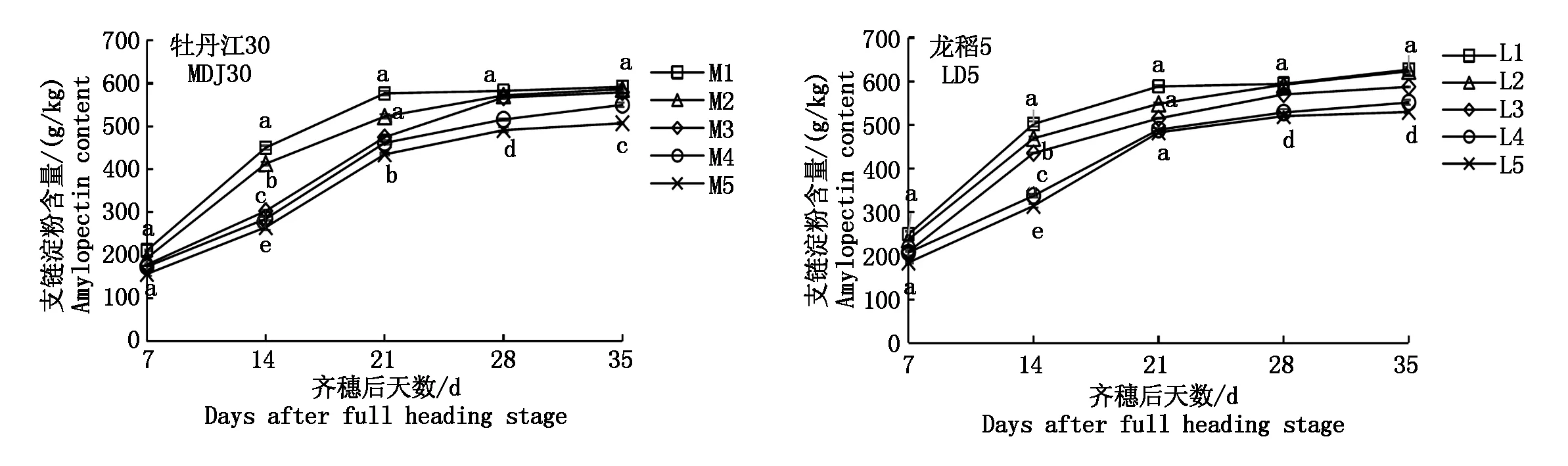
圖2 鹽脅迫下籽粒支鏈淀粉含量動態變化Fig.2 Dynamic changes of amylopectin content in kernels under salt stress
2.2 鹽脅迫下籽粒淀粉及其組分含量分析
寒地粳稻籽粒淀粉及其組分含量(齊穗后35 d)分析如表1所示。M4處理和L5處理下寒地粳稻籽粒直鏈淀粉含量最高,且均顯著高于對照水平。 2個供試品種在不同濃度鹽脅迫下的應激能力不同,牡丹江30各處理籽粒直鏈淀粉含量較對照的增幅較大,為2.43%~9.72%,龍稻5增幅較小,為1.42%~9.00%。
與對照水平相比,鹽脅迫下寒地粳稻籽粒支鏈淀粉含量下降,除M2、L2處理與對照差異不顯著外,其余各處理均顯著低于對照水平。總淀粉含量變化趨勢同支鏈淀粉。鹽脅迫下牡丹江30籽粒支鏈淀粉、總淀粉含量較對照的下降幅度分別為1.09%~18.62%和0.25%~12.09%,龍稻5分別為0.58%~15.46%和0.12%~9.9%。可以發現,大量的NaCl堆積嚴重影響籽粒淀粉形成,且品種對鹽脅迫的反應有所不同,耐鹽品種龍稻5的表現好于鹽敏感品種牡丹江30。從M1與M2、L1與L2可以看出,總淀粉含量以及淀粉組分含量都表現為差異不顯著,這充分說明寒地粳稻可抵御來自低濃度鹽脅迫所帶來的毒害。
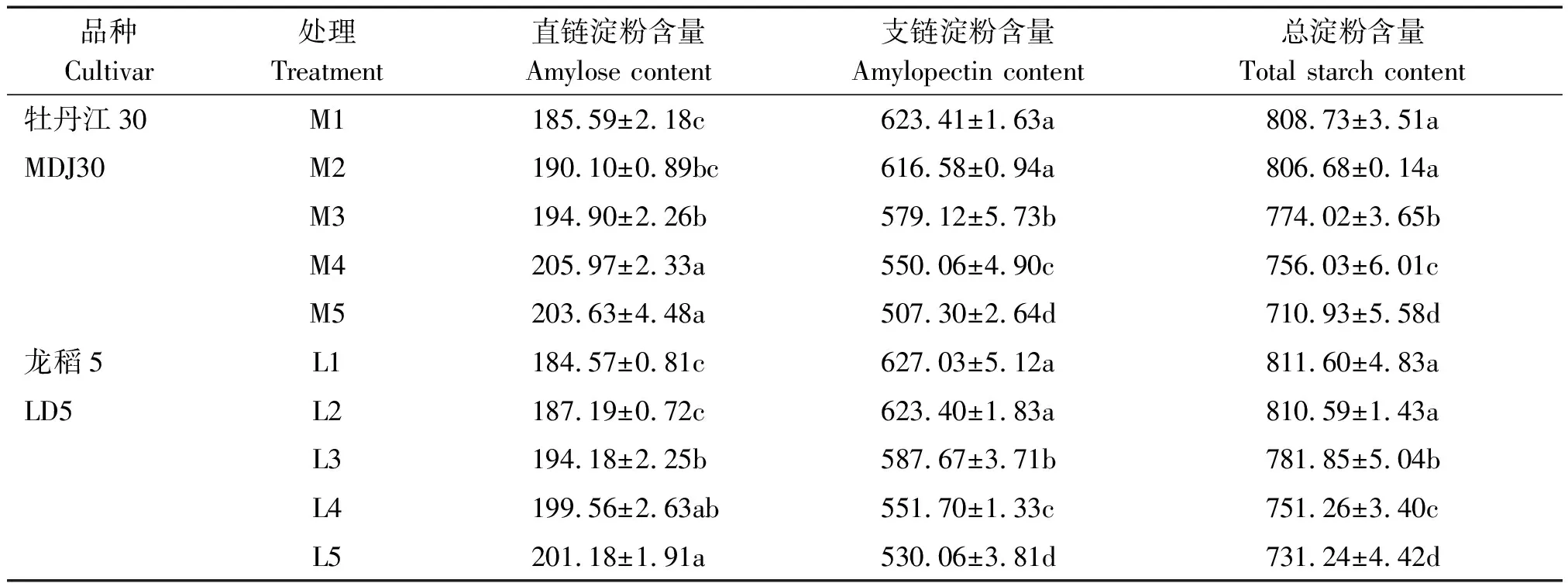
表1 鹽脅迫下籽粒淀粉及其組分含量Tab.1 Starch component contents in kernels under salt stress g/kg
注:表中不同的小寫字母表示5%水平的差異顯著。表3同。
Note: Values followed by different small letters are significantly different at the 5% probability levels. The same as Tab.3.
2.3 鹽脅迫對淀粉合成關鍵酶活性的影響
2.3.1 淀粉分支酶(Q酶)的動態變化 如圖3可知,隨著齊穗后天數的增加,寒地粳稻籽粒Q酶活性(以鮮質量計)體現為單峰曲線變化趨勢。但各處理Q酶活性抵達峰值的日期不同,于齊穗后21 d抵達峰值的處理有:M1、M2、L1、L2、L3,其中M1同M2差異不顯著,L1同L2、L3差異顯著;其余處理于齊穗后28 d達到峰值,其中M3、M4、M5差異不顯著,L4、L5差異不顯著。2個品種籽粒Q酶活性對鹽脅迫的響應不同,鹽敏感品種牡丹江30在土壤含鹽量達到0.150%時,Q酶活性嚴重受抑,出現峰值后移現象,而耐鹽品種龍稻5則是在土壤含鹽量達到0.225%才出現以上現象。同時,隨著土壤含鹽量的增高,酶活性峰值下降明顯,其具體表現為M1>M2>M4>M3>M5、L1>L2>L3>L4>L5。

圖3 鹽脅迫下寒地粳稻籽粒Q酶活性的變化規律Fig.3 Changes of Q activity in kernels of Japonica rice in cold-region under salt stress
2.3.2 可溶性淀粉合成酶(SSS)的動態變化 如圖4可知,寒地粳稻籽粒SSS活性(以鮮質量計)隨齊穗后天數的增加呈單峰曲線變化,但各處理峰值出現的日期不盡相同,具體表現為:M1、M3、L1、L2、L3處理于齊穗后14 d達峰值,其中,M1同M3差異不顯著,L1同L2、L3差異不顯著;其余處理均在齊穗后21 d達峰值,且在齊穗后21 d達峰值的品種內各處理均表現為差異不顯著。通過對比2個品種可以看出,當土壤含鹽量達到0.075%時,牡丹江30受影響較大,峰值日期推遲(除M3處理);龍稻5表現較好,當土壤含鹽量達到0.225%時,才出現峰值后移現象,其峰值具體表現為M1>M4>M2>M5>M3、L2>L1>L4>L5>L3。
2.3.3 ADPG焦磷酸化酶的動態變化 由圖5可知,寒地粳稻籽粒ADPG焦磷酸化酶活性(以鮮質量計)隨生育進程的推進表現為單峰曲線,但各處理到達峰值日期存在差別。M1、M2、M3、L1、L2、L4在齊穗后14 d達峰值,其中M1同M2、M3差異顯著,L1同L2、L4差異顯著;其余處理在齊穗后21 d達峰值,其中M4、M5差異顯著,L3、L5差異不顯著。鹽脅迫下籽粒ADPG焦磷酸化酶活性峰值變化趨勢具體表現為M1>M2>M3>M4>M5、L1>L2>L3>L5>L4。且耐鹽品種龍稻5各處理峰值酶活性高于鹽敏感品種牡丹江30各處理(除M4、L4外)。
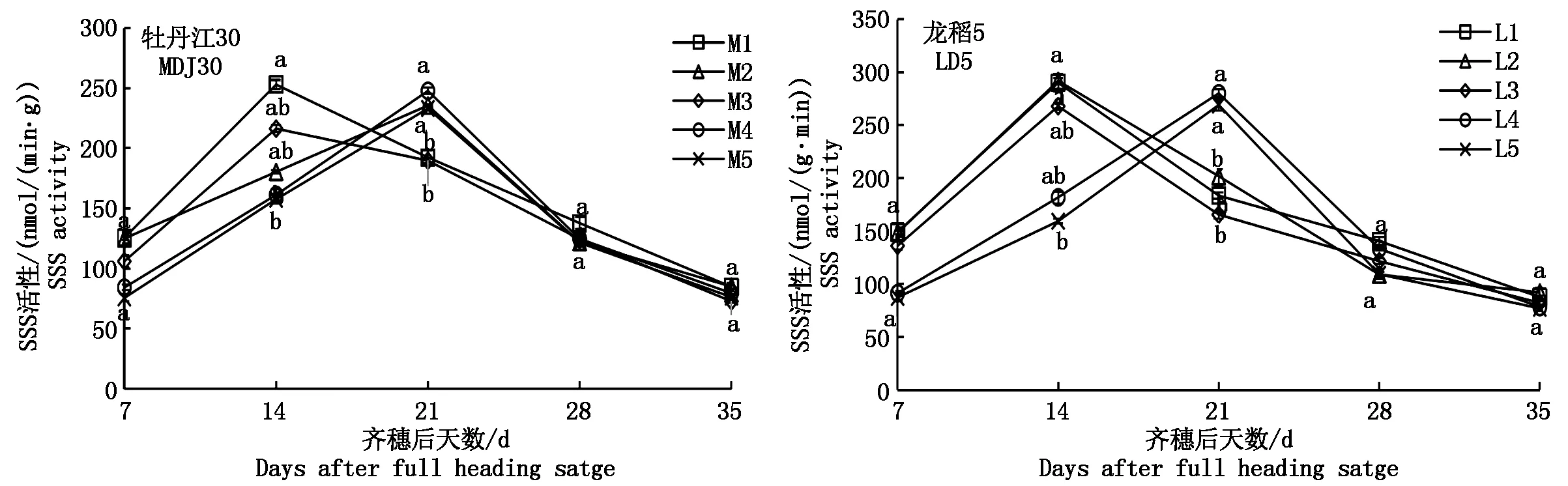
圖4 鹽脅迫下寒地粳稻籽粒SSS活性的變化規律Fig.4 Changes of SSS activity in kernels of Japonica rice in cold-region under salt stress
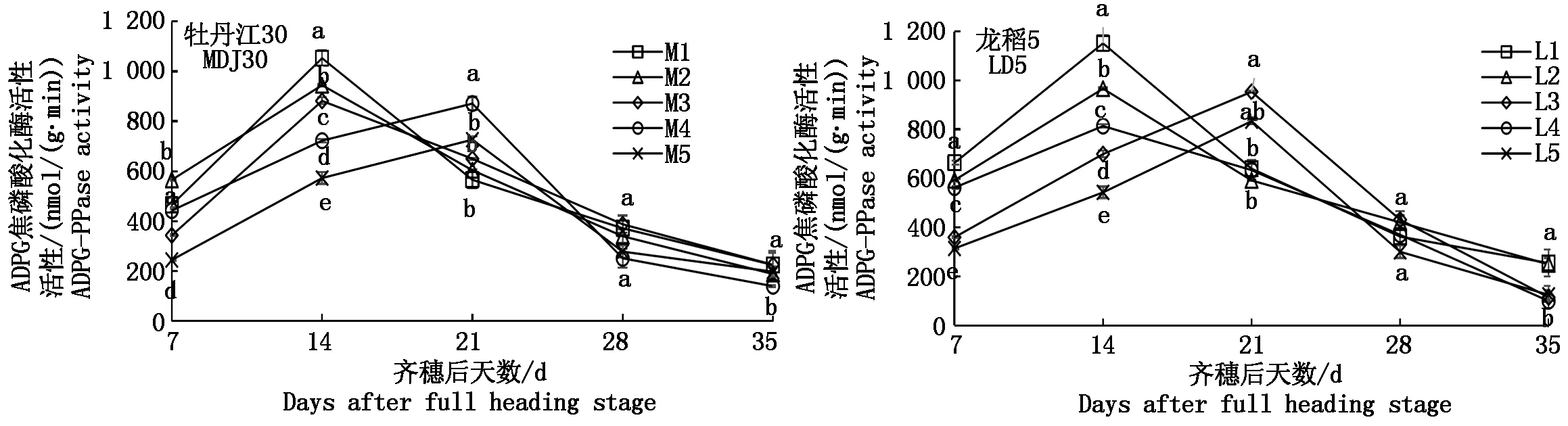
圖5 鹽脅迫下寒地粳稻籽粒ADPG焦磷酸化酶活性的變化規律Fig.5 Changes of ADPG-PPase activity in kernels of Japonica rice in cold-region under salt stress
2.4 淀粉合成關鍵酶活性與淀粉及淀粉組分之間的相關分析
如表2所示,寒地粳稻籽粒支鏈淀粉含量以及總淀粉含量與齊穗后14,21,28,35 d的Q酶活性呈顯著或極顯著正相關,與齊穗后14,21,35 d的SSS活性呈顯著或極顯著正相關,可以發現,籽粒Q酶與SSS共同完成支鏈淀粉的合成,通過影響支鏈的淀粉含量最終決定總淀粉含量。籽粒總淀粉含量與ADPG焦磷酸化酶在齊穗后7,35 d呈極顯著正相關,這說明ADPG焦磷酸化酶主要作用于籽粒灌漿過程的前期與后期。
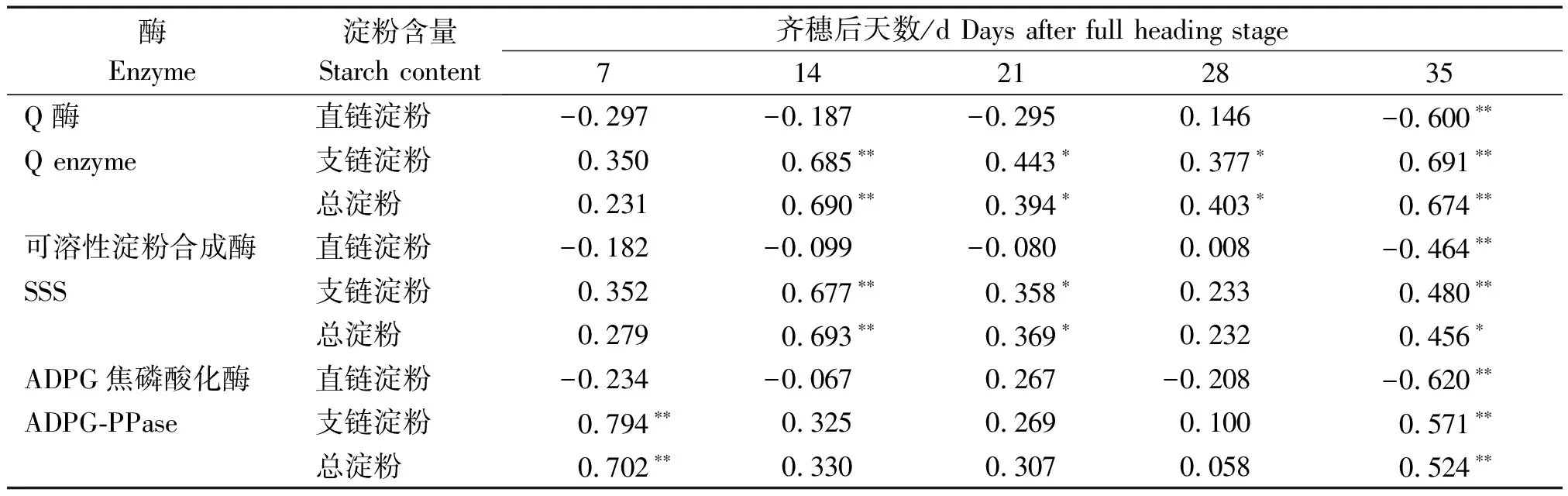
表2 鹽脅迫下寒地粳稻籽粒 ADPG 焦磷酸化酶、SSS、Q 酶活性與淀粉含量之間的關系Tab.2 Relationship of ADPG-PPase, SSS, Q enzyme activities, and starch contents in kernels of Japonica rice in cold-region under salt stress
注*、**分別表示在0.05和0.01水平上顯著相關。
Note*,**indicate significant correlation at the 0.05 and 0.01 levels,respectively.
2.5 鹽脅迫對寒地粳稻產量及產量構成因素的影響
由表3可見,鹽脅迫下2個品種的各產量構成因素及理論產量較對照(M1、L1)均有不同程度下降。牡丹江30各處理有效穗數的降幅為0~27.08%,龍稻5為4.65%~41.86%,具體表現為:M1同M2、M3差異不顯著,但顯著高于M4、M5;L1同L2差異不顯著,但顯著高于L3、L4、L5。牡丹江30穗粒數的降幅為7.32%~39.46%,龍稻5為4.69%~25.51%。其中,M1的穗粒數顯著高于各處理;L1與L2、L3處理差異不顯著,但顯著高于其余處理。
牡丹江30的結實率,各處理較對照下降0.02~14.33百分點,而龍稻5則為1.20~11.45百分點。同時,牡丹江30的千粒質量、理論產量降幅依次下降為:7.97%~22.98%,14.41%~71.30%;龍稻5為1.24%~15.08%,10.92%~67.49%。2個品種的處理間表現相同,即當土壤含鹽量達到0.150%時,結實率及理論產量顯著下降;當土壤含鹽量達到0.225%時,千粒質量顯著下降。
通過分析可知,M1和M2、L1和L2處理各指標均表現為差異不顯著(除牡丹江30的穗粒數),說明水稻可承受較低濃度的鹽脅迫。同時可以發現,牡丹江30的穗粒數及結實率在處理間差異較大,而龍稻5則是有效穗數及結實率差異較大,說明耐鹽性不同的品種在鹽脅迫下產量構成因素的變化存在差異。通過對比2個品種,除有效穗數外,牡丹江30的各產量構成因素降幅均高于龍稻5,有效穗數降幅較小的原因可能是牡丹江30具有相對較好的分蘗能力。

表3 鹽脅迫對寒地粳稻產量構成因素的影響Tab.3 Effects of salt stress on yield components of Japonica rice in cold-region
3 結論與討論
在一系列淀粉合成相關酶的調控作用下,籽粒中淀粉開始形成并逐步積累,這一動態變化即為水稻籽粒充實的過程。已有大量研究指出,ADPG焦磷酸化酶、SSS、Q酶在高等植物的光合及非光合器官中都發揮著至關重要的調控功能,同時,由NaCl引起的不良影響并不會隨NaCl的消失而減弱[20]。本研究發現,隨土壤含鹽量的提升,峰值酶活性波動較大。這可能是在高濃度鹽脅迫下(0.150%~0.300%),植物體內由于累積了大量Na+使得原有的離子平衡被破壞,在一定程度上損傷了植物的基礎蛋白質結構,從而使酶活性大幅下降[21]。通過淀粉合成關鍵酶動態變化可看出,鹽脅迫延遲酶活性到達峰值的日期,分析原因可能是作物本身需要時間來適應鹽脅迫。有研究指出耐鹽品種較鹽敏感品種更能有效地控制外界Na+向地上部的轉運,可在一定范圍避免地上部遭受鹽害[22-23]。通過導致淀粉合成關鍵酶活性峰值推遲的土壤含鹽量來看,耐鹽性品種龍稻5更能有效避免Na+所帶來毒害,在鹽脅迫下籽粒仍能保持相對較高的淀粉合成關鍵酶活性。
稻米品質取決于淀粉的組分與含量。步金寶[24]研究發現,鹽處理顯著抑制粳稻籽粒淀粉的最終形成,且淀粉組分積累速率受到影響。本研究發現,隨著鹽濃度的升高,2個供試品種籽粒的總淀粉及支鏈淀粉含量降幅逐漸增大,低濃度鹽處理(M2與 L2)與對照差異不顯著,而中高濃度鹽處理導致總淀粉及支鏈淀粉含量下降明顯,但具體降幅因品種的耐鹽性而異。本研究同時還發現,鹽脅迫可導致籽粒直鏈淀粉含量顯著增加,直鏈淀粉含量的增幅隨著鹽濃度的升高而增大,這與羅成科等[25]的研究結果基本一致。分析原因可能是由于植株在遭受鹽脅迫時可促進直鏈淀粉合成,從而改變直鏈淀粉與支鏈淀粉的比例,以此來抵御不良環境帶來的影響,但這一現象是否為水稻的一種應激反應還有待進一步研究。
通過試驗結果分析可知,籽粒支鏈淀粉含量與Q酶活性呈顯著或極顯著正相關(除齊穗后7 d),并與 SSS活性表現為灌漿中后期顯著或極顯著正相關,說明鹽脅迫下寒地粳稻籽粒支鏈淀粉的合成與 SSS和Q酶關系密切,這在前人的研究中已有體現[26-27]。籽粒淀粉含量與灌漿前期、后期ADPG焦磷酸化酶活性呈極顯著正相關,淀粉及其組分在灌漿前期都有一個快速增長的過程,說明 ADPG焦磷酸化酶的活性與灌漿速率密切相關,這和前人的研究結果基本一致[28-29],后期呈極顯著正相關可能是由于當灌漿過程進行到后期,植株趨于成熟啟動衰老機制,ADPG焦磷酸化酶通過抑制淀粉合成速率來減緩代謝。
產量形成依靠籽粒干物質的積累,又取決于籽粒淀粉合成積累的多寡[30]。本研究發現,2個供試品種的理論產量在土壤含鹽量為0.075%時與對照差異不明顯,當土壤含鹽量達到0.150%時抑制效果明顯。關于鹽脅迫對水稻產量構成因素影響的說法不一:楊福等[31]研究得出鹽堿脅迫下產量的下降是由穗粒數減少及千粒質量減輕所引起;余為仆[32]通過對產量構成因素的分析發現,鹽脅迫導致的降產在多方面因素的共同作用下完成。本研究結果表明,產量構成因素各指標隨土壤含鹽量的提高而下降,但低濃度的鹽處理(M2、L2)與對照差異不顯著(除牡丹江30的穗粒數),當土壤含鹽量超過0.075%時,各產量構成因素受影響顯著,可以發現,結實率的變化最為突出。但寒地粳稻耐鹽臨界值是否為0.075%還需進一步探討。此外,本研究結果為盆栽試驗所得,可對大田生產有一定指導,但具體應用還有待進一步驗證。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:19:16
汽車實用技術(2022年10期)2022-06-09 11:16:58
音樂探索(2022年2期)2022-05-30 21:01:37
收藏界(2019年3期)2019-10-10 03:16:40
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
中國非營利評論(2017年1期)2017-11-09 03:09:10
海外華文教育(2017年8期)2017-11-07 04:42:02
現代語文(2016年21期)2016-05-25 13:13:50
