硼促進缺鐵條件下擬南芥根系細胞壁鐵的再利用
2019-04-01 06:28:46朱曉芳沈仁芳
植物營養與肥料學報 2019年2期
關鍵詞:植物
吳 啟,朱曉芳,沈仁芳*
(1 土壤與農業可持續發展國家重點實驗室/中國科學院南京土壤研究所,南京 210008;2 中國科學院大學,北京 100049)
鐵在地殼中含量十分豐富,約占地殼總重量的5%左右。同時鐵也是動植物生長發育所必需的微量元素,作為許多功能蛋白的重要組成部分,鐵能參與高等植物的光合作用和氮同化等生理生化過程[1-2]。雖然在土壤中鐵的含量很高,但其主要是以Fe3+復合物的形式存在,而植物可以直接吸收利用的Fe2+含量卻是很低的。尤其是在堿性和石灰性土壤中,由于高pH和強氧化作用使得土壤中的Fe3+溶解度極低,從而抑制了土壤中游離鐵的有效性,最終限制植物的生長發育[3-4]。據統計,全世界約有30%的土壤處于鐵饑餓狀態[5],地中海地區、北美大陸、南美的部分地區都嚴重缺鐵。而我國西南地區、淮北平原以及西北等地均有缺鐵現象的發生[6]。缺鐵導致的癥狀首先是新葉出現黃化現象,鐵元素由地下部向上運輸的過程受阻,從而造成營養元素的分配不均勻,最終導致作物營養不良,品質和產量嚴重受限,進而影響人類的飲食健康[7]。
高等植物在漫長的進化過程中逐漸形成了一整套完善而復雜的應答缺鐵的策略。根據植物對缺鐵表現的形態學差異和生理變化的不同,人為地將這些植物劃分為非禾本科植物和雙子葉植物所共有的機理Ⅰ和禾本科所特有的機理Ⅱ。在受到缺鐵脅迫時,機理Ⅰ植物主要有以下三個步驟響應缺鐵信號:首先,植物根系表面的ATP酶 (H+-ATPase) 會首先活化,向根系外分泌H+來酸化土壤,增加Fe3+的溶解度來促進植物對鐵的吸收[8]。如擬南芥中的AHA(Arabidopsis H+-ATPase isoform) 家族基因就是在該過程起著重要的作用[9]。其次,根系表面溶解的Fe3+需要轉化成Fe2+形式才能被植物所吸收,位于根系細胞膜表面的鐵離子螯合還原酶 (ferric-chelate reductase oxidase, FRO) 可以將根表面的三價鐵還原成二價鐵以供植物吸收利用[10]。最后,還原后的Fe2+會在一系列鐵轉運蛋白 (iron-regulated transport, IRT)的運輸下轉運到根皮層細胞[11],之后再進一步通過各類轉運蛋白運輸到細胞的各個組織器官供植物體吸收。機理Ⅱ植物包括水稻、玉米、小麥和大麥等多種農作物。研究發現,在嚴苛的缺鐵條件下,禾本科植物往往能比雙子葉或非禾本科植物表現出更高的抗性[12],其主要原因是這些植物在受到缺鐵信號脅迫時往往能夠分泌麥根酸類 (mugineic acid, MA) 物質[13],這類物質能夠與根系微環境中的Fe3+螯合形成Fe3+-PS復合體,最終被根表皮細胞的細胞膜上的YS/YSL (yellow stripe/yellow stripe like) 家族轉運蛋白吸收[14]。
作為植物接觸和響應外界信號的第一道屏障,細胞壁在植物抵御鋁毒和鎘毒的過程中發揮了重要的作用[15-16]。Chang等[17]研究表明,在鋁毒脅迫下,植物細胞中的鋁主要集中在細胞壁的果膠中。為了進一步明確果膠在植物耐鋁中所發揮的作用,Zheng等[18]利用果膠酶可以降解果膠的原理,發現去除細胞壁果膠后,細胞壁可結合的Al含量減少了約50%,說明細胞壁果膠成分在結合Al方面確實起著重要的作用。而對于細胞壁緩解鎘毒的相關研究,Zhu等[16,19]研究發現,細胞壁組分中的半纖維素可以結合鎘,外源添加生長素可增強半纖維素對鎘的結合能力,減少鎘由地下部向上運輸的過程,從而緩解植物鎘毒的危害。除了可以緩解鎘毒和鋁毒,最近的研究表明,細胞壁還參與植物抵抗缺鐵逆境的過程[20]。Zhu等[21]研究發現,NaCl可以通過激發根系細胞壁鐵的再利用和再分配機制來緩解擬南芥的缺鐵癥狀,進一步研究顯示,脫落酸作為一類植物逆境激素,可以通過參與調解細胞壁組分的合成來響應植物缺鐵的信號。
硼也是植物生長發育所必需的微量元素,在植物中主要以硼-糖復合物的形式存在于細胞壁中[22],可以用來維持植物細胞壁的穩定性[23]。同時,硼還能夠參與碳水化合物的運輸、花粉萌發和花粉管生長等一系列生理生化過程,對作物的產量和品質都有極其重要的影響[24]。一般認為,細胞中 80%以上的硼可以和細胞壁的果膠成分相結合,形成大小合適的細胞壁孔徑,從而調節其他養分的吸收和轉運[25]。最近研究發現,硼可以通過膠聯細胞壁果膠成分來緩解植物體內的鋁毒[26-27],硼還可以和鈣協同作用緩解植物鋁毒的癥狀[28],鋅硼之間的交互作用對植物生長、光合作用和水分的吸收都起著至關重要的作用[29]。但是,外源添加硼是否能夠緩解植物的缺鐵癥狀還鮮有研究,本試驗以模式作物擬南芥為研究材料,試圖揭示在缺鐵的條件下,微量元素硼能否可以通過調節細胞壁組分來提高植株體內有效鐵的含量,從而達到緩解植物缺鐵的目的。
1 材料與方法
1.1 植物材料和培養條件
供試材料為野生型擬南芥Col-0 (Columbia ecotype)。挑選無病蟲害的擬南芥種子,首先用75%乙醇處理 5 min,對種子表面進行消毒,再用無菌水清洗3次;然后將用全營養液浸泡過的海綿塞入1.5 mL去底eppendorf離心管中;再將消毒完畢的種子用牙簽點在濕潤的海綿上,隨后放入光照培養箱中。培養室內植物生長的條件:溫度為24°C,光照 16 h,黑夜 8 h,光照強度為 140 μmol/(m2·s)。
全營養液成分:大量元素濃度 (mmol/L) 為KNO36.0、Ca (NO3)24.0、MgSO41.0、NH4H2PO40.1;微量元素濃度 (μmol/L) 為 Fe (III) -EDTA 50、H3BO312.5、MnSO41.0、CuSO40.5、ZnSO41.0、H2MoO40.1、NiSO40.1。用1 mol/L HCl或NaOH調節溶液pH為 5.6。
在光照培養箱培養7 d后,將幼苗移至全營養液(pH 5.6) 生長2周。最后挑選長勢一致的擬南芥移入1.25 L用黑色塑料袋包裹的罐子里,每個罐子種 9 棵苗。對以上樣品進行如下處理:+Fe (pH 5.6的全營養液);+Fe + 100 μmol/L H3BO3;+Fe + 1000 μmol/L H3BO3;-Fe (pH 5.6不含鐵的營養液);-Fe + 100 μmol/L H3BO3;-Fe + 1000 μmol/L H3BO3。處理中每隔3 d換一次營養液,處理共進行了7 d。
1.2 葉綠素含量的測定
利用便攜式葉綠素儀 (Konica Minolta SPAD-502,Tokyo, Japan) 測定表征植株新葉葉綠素含量的SPAD值。
1.3 總鐵含量的測定
切除處理后擬南芥的根系和地上部,用蒸餾水清洗3次,在70°C烘箱中放置2 d后,分別稱重,記錄干重。再把樣品放入20 mL用酸浸泡過的消煮管中,加入2 mL HNO3/HClO4 (4∶1, v/v),然后將消煮管放在消煮爐中130°C加熱,直至樣品澄清透亮,最后加入8 mL蒸餾水定容至10 mL,濾紙過濾后采用電感耦合等離子體原子發射光譜法 (ICP-AES)測定溶液中的鐵含量[30]。
1.4 有效鐵含量的測定
參照Lei等的方法[31],具體操作步驟如下:將處理后的植株根系和地上部分別用去離子水清洗后稱取鮮重,加入液氮后在研缽中充分研磨,最后加入8 mL去離子水充分浸提。24 h后對浸提液13200 rpm離心10 min,取上清液采用電感耦合等離子體原子發射光譜法 (ICP-AES) 測定溶液中的鐵含量。
1.5 細胞壁成分的分級提取及細胞壁組分鐵的測定
細胞壁的提取:將擬南芥根系在去離子水下沖洗3遍后,用吸水紙吸干多余的水分。用鑷子將根系轉移到干凈的研缽中,加入過量的液氮充分研磨,然后加入8 mL 75%乙醇將研磨液轉移至10 mL離心管中,放在漩渦震蕩儀上使之充分混勻,靜置20 min后5000 rpm離心10 min后倒掉上清液。然后依次加入丙酮、1∶1甲醇、氯仿的混合液以及甲醇,每次均在漩渦震蕩儀上震蕩數分鐘使提取液充分混勻, 然后靜置20 min后離心棄上清液。最后細胞壁粗提物放入冷凍干燥機干燥,4°C保存備用[32]。
果膠的提取:稱取已經提取好約2 mg的細胞壁放在1.5 mL離心管中,加入1 mL去離子水后在100°C沸水中水浴1 h,然后在12000 rpm下離心10 min,用移液槍吸取上清液至5 mL離心管中,再重復以上步驟2遍,最后用去離子水定容至3 mL即為所提果膠溶液[33]。
半纖維素的提取:在提取果膠后的殘渣中加入1 mL 24% KOH,室溫靜置12 h后,12000 rpm下離心 10 min,取上清液于5 mL離心管中,再重復一次上述步驟,將兩次所得的上清液合并在一起,用去離子水定容至2 mL即為半纖維素溶液。
細胞壁鐵含量的測定:稱取約2 mg的粗提細胞壁于1.5 mL離心管中,加入1 mL 2 mol/L HCl后放入搖床室溫震蕩3 d,然后利用電感耦合等離子體原子發射光譜法 (ICP-AES) 測定細胞壁中的鐵含量。
細胞壁組分鐵含量的測定:分別取果膠和半纖維素溶液利用電感耦合等離子體原子發射光譜法(ICP-AES) 測定其鐵含量。
1.6 半纖維素含量的測定
半纖維素含量一般用總糖的含量來表示。具體操作如下:取200 μL半纖維素于1.5 mL離心管中,隨后在離心管中加入10 μL 80%的苯酚和1 mL濃硫酸,室溫靜置15分鐘后再放入100°C沸水中水浴15分鐘,快速取出離心管置于冰上冷卻。然后在490 nm波長下比色。標準曲線的制作參照Dubios等方法[34]。
1.7 RNA提取和基因表達分析
擬南芥幼苗在+Fe + 100 μmol/L H3BO3、+Fe +1000 μmol/L H3BO3、-Fe+100 μmol/L H3BO3、-Fe +1000 μmol/L H3BO3溶液中處理7 d后,收集根系放入液氮中,隨后在研缽中加入液氮充分研磨,然后用植物RNA提取試劑盒 (TIANGEN) 并按照相關說明要求提取RNA,提取后的RNA用nanodrop檢測樣品濃度和質量。先通過RNA濃度測定換算成含有1 μg的樣品,然后用PrimeScript RT試劑盒 (Takara)進行反轉錄合成cDNA。反轉錄后的cDNA放入-20°C冰箱保存。
實時熒光定量PCR采用TAKARA公司生產的SYBR Premix ExTaq體系進行 (Takara Bio,Inc,Japan)。首先將反轉得到的cDNA稀釋10倍用于定量PCR的模板。PCR循環體系為10 μL體系,其中1 μL 稀釋后的 cDNA,5 μLSYBR Premix ExTaq,0.5 μL的正向和反向引物,3 μL的無菌蒸餾水。PCR反應的程序為: 95°C初預熱1 min,95°C變性15 s,55°C退火15 s,72°C延伸20 s,整個程序共45個循環。以tublin作為內參基因,采用公式2-ΔΔCT計算不同處理間基因的相對表達量。上述試驗每個均有4個生物學重復和3個機械重復。
相關基因引物如下:
TUBLIN
上游引物 5’-AAGTTCTGGGAAGTGGTT-3’
下游引物 5’-CTCCCAATGAGTGACAAA-3’
FRD3
上游引物 5’-TTTTGTCGGGCGTTTAGG-3’
下游引物 5’-TTGCTGTGGCTGGTTGGT-3’
YSL2
上游引物5’-GGATACTTATTCTTCTCCCTT GTC-3’
下游引物 5’-CCATCGTTTTTTCCTGCC-3’
NAS1
上游引物5’-CATGATCTTCCACACAACGG AC-3’
下游引物 5’-CGACGTCATATTGGTCAA GGC-3’
1.8 一氧化氮 (NO) 含量的測定
利用3-氨基-4-甲氨基-2′-7′二氟熒光素測定擬南芥根系NO含量。具體操作如下,取根尖1 cm處在20 mmol/L的HEPE緩沖液中沖洗20 min,然后用鑷子將根尖剪出放在裝有10 μmol/L熒光染料的離心管中避光染色30 min,最后再用HEPE緩沖液沖洗根系3次,每次15 min。制片,在熒光顯微鏡下觀察,拍照,利用photoshop 軟件計算熒光度。
1.9 數據處理
數據采用Excel2013、Sigmaplot13.0、Statistix8.0進行分析整理和繪圖。
2 結果與分析
2.1 外源硼對擬南芥缺鐵癥狀的緩解作用
為了探究外源添加硼在缺鐵脅迫中的作用,本試驗以模式植物擬南芥為材料,在正常和缺鐵的條件下采用不同濃度的硼處理后觀察植株表型并測定不同部位有效鐵含量。結果顯示,在正常全營養液條件下,添加外源硼不能改變植株的生長情況。當受到缺鐵脅迫時,植株新葉首先出現黃化現象,外源施加硼 (1000 μmol/L H3BO3) 可以顯著改善缺鐵癥狀 (圖1-A);利用便攜式葉綠素儀測定植株新葉的葉綠素含量 (SPAD值),結果表明,缺鐵情況下植株葉片的葉綠素含量隨著硼濃度的增加而顯著增加 (圖1-B)。值得關注的是,在缺鐵脅迫下,外源添加硼后可以顯著增加植株根系和地上部有效鐵的含量 (圖1-C、D),表明硼緩解缺鐵癥狀的主要原因可能是植株體內鐵的再利用機制。
2.2 外源硼對細胞壁鐵再利用的促進作用
在缺鐵的條件下根系仍含有一定的有效鐵含量,筆者猜測可能與植株體內鐵的再利用有關。因此為了探究其內在的鐵再利用機制,筆者用不同濃度梯度的硼酸處理擬南芥幼苗7 d,分別測定了加鐵和不加鐵情況下根系細胞壁鐵的含量和其組分上結合的鐵含量。結果顯示,與單獨缺鐵環境相比,外源添加硼后細胞壁鐵和半纖維素鐵含量都顯著降低 (圖2-A、B),且隨著硼酸濃度的增加,細胞壁組分結合的鐵含量降低。進一步測定了細胞壁的半纖維素含量發現,隨著外源硼酸濃度的增加,細胞壁半纖維素含量也逐漸減少 (圖2-C)。綜上分析,推測硼可以通過提高細胞壁鐵含量釋放來增加植物體內有效鐵的含量,從而確保植株在缺鐵的條件下正常生長。
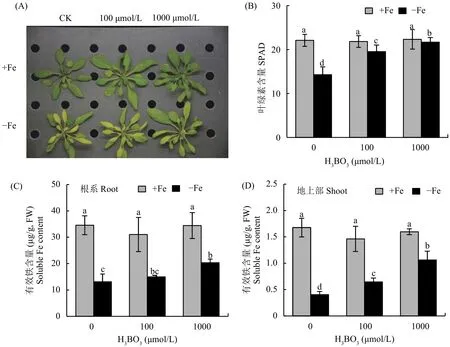
圖1 不同濃度硼處理后葉片表型、葉綠素含量以及根系和地上部有效鐵含量Fig. 1 Phenotype and chlorophyll content of leaves, and soluble Fe contents in root and shoot after different concentrations of boron treatments
2.3 外源硼對植株體內鐵由根系向地上部轉運的促進作用
外源添加硼可以通過增加地上部有效鐵的含量從而緩解植株的缺鐵癥狀,為了進一步探討其內在的生理機制,測定了植株根系和地上部的總鐵含量。試驗結果顯示,在鐵含量充足的條件下,硼不影響根系和地上部總鐵含量的積累;但是在缺鐵的條件下,與不施加硼相比,外源施加硼酸,尤其是施加 1000 μmol/L H3BO3,根系和地上部單位質量的總鐵含量都得到顯著增加 (圖3-A、B)。由圖3-C可以得出外源添加硼可以參與擬南芥體內的鐵由根系向地上部轉運的過程。
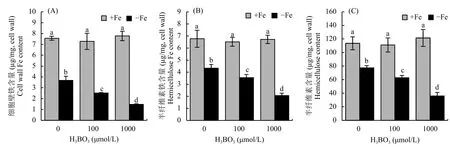
圖2 不同處理擬南芥細胞壁鐵含量及半纖維素和半纖維素鐵含量Fig. 2 Iron contents in cell wall, hemicellulose and hemicellulose content after different boron treatments
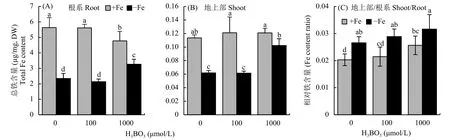
圖3 不同濃度硼處理后根系和地上部鐵含量Fig. 3 Total iron content in root and shoot after different concentrations of boron treatments
2.4 擬南芥體內鐵轉運相關基因表達的變化
前面的結果表明,外源添加硼后可以促進根系鐵向地上部的轉運,因此本試驗主要測定了外源添加硼前后這 3 個轉運蛋白的表達變化。結果顯示,鐵充足的情況下外源硼對這 3 個基因的影響不大。在缺鐵處理 7 d后,AtFRD3和AtNAS1基因表達顯著上調,卻抑制AtYSL2的表達 (圖4-A、B、C);然而再添加外源硼,特別是 1000 μmol/L硼酸后,這3 個基因的表達量顯著高于單純的缺鐵條件 (圖4),暗示硼可能主要是通過調節AtFRD3、AtNAS1以及AtYSL2基因的表達來增加其有效鐵從根部向地上轉運的過程。
2.5 擬南芥根系一氧化氮 (NO) 含量的變化及其作用
為了明確一氧化氮 (NO) 和硼對缺鐵信號的影響,我們首先研究了缺鐵脅迫下外源加入硼后對內源NO代謝的影響。結果顯示,在加鐵條件下,外源施加硼對根系NO含量沒有顯著影響 (圖5-A、B),但是在缺鐵情況下,外源施加硼后根系NO含量是不施加硼情況的 1.5 倍 (圖5-C、D),暗示信號分子NO可能參與了調控細胞壁鐵的再釋放過程。為了進一步確定NO的作用,我們又對以上4個處理分別外源添加NO清除劑c-PTIO,結果顯示在加入c-PTIO后,以上4個處理根系NO含量皆顯著降低,且加入c-PTIO后上述 4 個處理的NO含量均處于同一水平上,說明c-PTIO對擬南芥根系NO的清除作用十分明顯 (圖5-E、F、G、H)。探究缺鐵條件下在營養液中加入NO清除劑c-PTIO后,外源添加硼緩解擬南芥缺鐵的癥狀可以被逆轉,這進一步說明硼緩解缺鐵的癥狀是由信號分子NO所介導的(表1)。
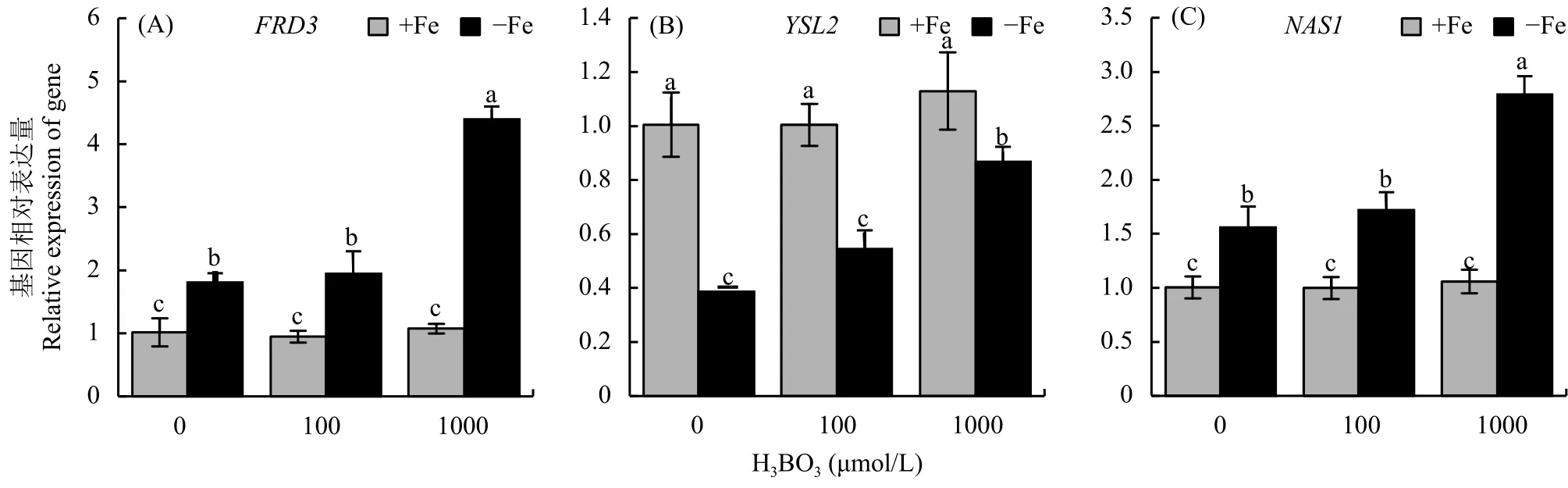
圖4 不同濃度硼處理后鐵轉運相關基因相對表達量Fig. 4 The relative expression of iron transporter after different boron treatments
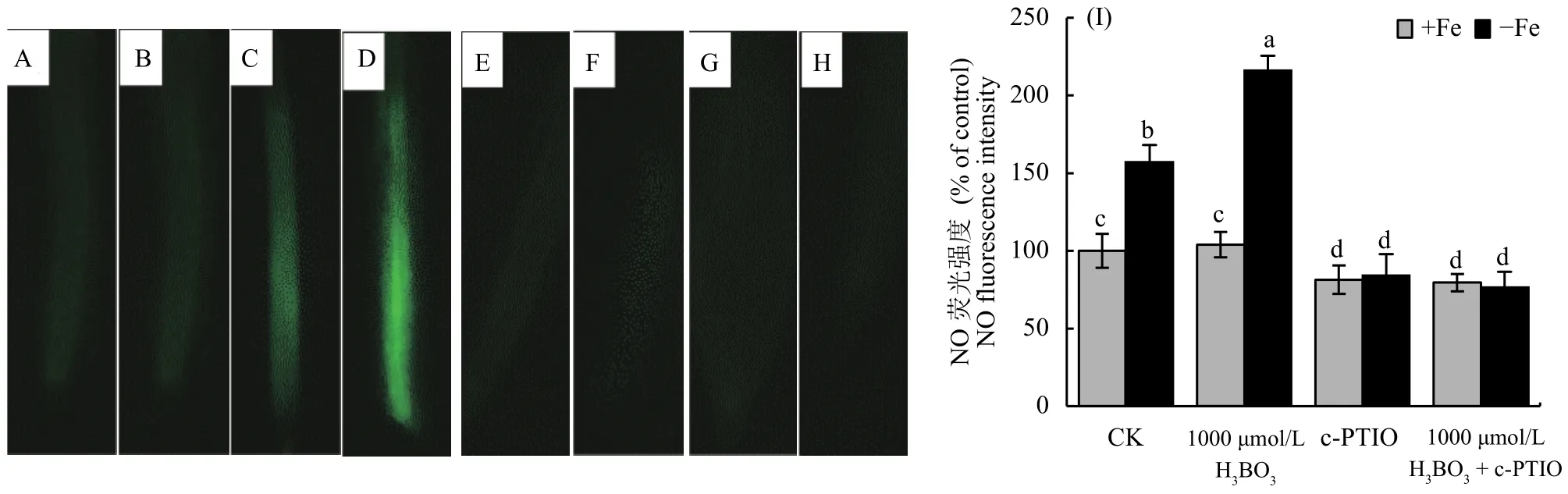
圖5 不同濃度硼處理后對根系NO含量的影響Fig. 5 Effect of different boron treatments on NO production in root
3 討論
近年來,微量元素硼在植物逆境中發揮的重要作用也越來越受到人們的重視,比如可以在豌豆(Pisum sativum) 中抵御鋁毒。研究表明,硼可以促進鋁在細胞壁果膠中的累積,并且通過抑制鋁的解吸作用來降低鋁在細胞中的可移動性。進一步結果顯示,硼還可以通過降低果膠的甲酯化程度來增加細胞表層的負電荷,最終達到固定Al3+的作用[27]。不僅如此,硼還在植物鹽害中起關鍵的作用,Bastias等[35-36]發現硼可以通過維持細胞壁的延展性來調節植物葉片的水勢和滲透壓力,遺傳學證據顯示,水孔通道蛋白也能夠參與這種硼緩解鹽害的過程。在本試驗中發現,隨著外源添加1000 μmol/L H3BO3時,在缺鐵的條件下,擬南芥根系和地上部的有效鐵含量都能夠顯著提高 (圖1)。很明顯在營養液缺鐵的情況下,仍然能夠檢測到植物根系含有可溶性鐵。根據前人的研究發現,細胞壁,尤其是細胞壁的組分半纖維素,是擬南芥根系鐵的重要結合位點。因此我們猜測在缺鐵的條件下,硼增加擬南芥體內有效鐵的含量是由于細胞壁鐵的再利用機制導致的。
同時,一氧化氮 (NO) 是植物體內重要的信號分子,它在調節植物生長發育以及幫助植物抵御逆境脅迫中發揮著重要的作用。本試驗中發現,外源添加硼酸可以促進擬南芥根系細胞壁鐵的釋放來增加體內的有效鐵含量,那么硼又是通過什么信號途徑介導缺鐵反應的?因為之前的相關鐵含量和基因表達量數據均顯示1000 μmol/L H3BO3可以顯著緩解缺鐵癥狀,因此本試驗選取了+Fe、+Fe + 1000 μmol/L H3BO3、-Fe、-Fe + 1000 μmol/L H3BO34 個處理分別測定其根系NO含量,檢測到缺鐵環境中加硼后也能促進根系NO的釋放,暗示信號分子NO確實也能夠參與調控植物體內鐵的再利用過程,而在加入NO清除劑c-PTIO后,外源添加硼緩解擬南芥缺鐵的癥狀可以被逆轉 (表1),這也就進一步解釋了硼緩解缺鐵的癥狀是由信號分子NO所介導的。
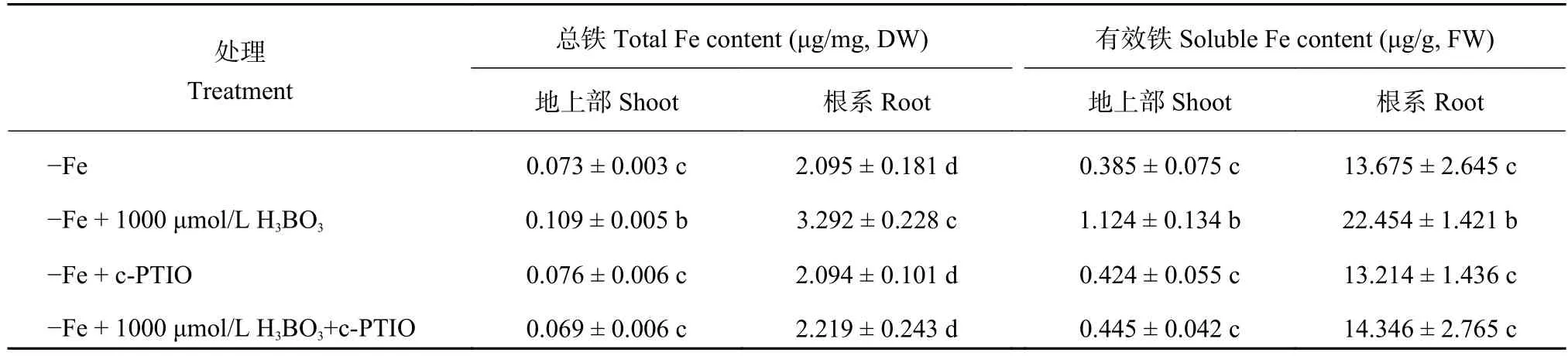
表1 NO清除劑c-PTIO對缺鐵情況下根系和地上部鐵含量的影響Table 1 Effect of NO scavenger c-PTIO on iron contents in root and shoot
擬南芥根系細胞壁在提高植物體內鐵的再利用方面起著十分重要的作用。Andersen等[37]發現,在藻類中細胞壁的不同結構可以影響鐵在體內的分布。進一步研究顯示,細胞壁木質纖維素的結構可以影響水稻和擬南芥種子中鐵含量的累積,具體表現為將位于細胞壁纖維素表面的鐵結合肽和碳水化合物結合肽的融合多肽轉入擬南芥和水稻后可以明顯看到種子中鐵含量的積累和生物量的增加,對于提高糧食作物的產量和解決全世界范圍內鐵缺乏的問題具有積極的指導意義[38]。Zhu等[21]研究發現,NaCl可以緩解擬南芥的缺鐵癥狀,其主要原因是缺鐵時外源添加NaCl后細胞壁組分半纖維素含量發生變化,從而導致細胞壁半纖維素上結合的鐵游離到組織中供植物吸收,最終達到緩解植物缺鐵的效果;其后續的研究還發現,植物激素脫落酸 (ABA)可以作為一種信號分子,參與植物體內細胞壁鐵的再轉運機制。植物細胞壁的半纖維素主要是在高爾基體中合成,在擬南芥中半纖維素最主要的成分是木葡聚糖。傳統的觀點認為,半纖維素的主要成分不帶電或者帶有中性電荷,因此很難像細胞壁果膠等成分一樣結合金屬陽離子。但是由于果膠與半纖維素之間能以共價鍵相結合,所以半纖維素上的金屬陽離子結合位點有可能會被果膠中不同形式的多糖所占據[39]。最新的研究結果顯示,細胞壁的半纖維素可能通過與鋁離子形成某種形式的復合物來參與調控植物體內鋁的吸收與釋放過程[33]。但是有關細胞壁半纖維素上鐵的結合位點的研究仍未有報道。本試驗中,在缺鐵條件下外源添加硼后,擬南芥根系細胞壁鐵的含量顯著降低,同時細胞壁組分半纖維素含量和半纖維素結合的鐵含量也相應減少,因此筆者認為外源添加硼可以通過改變細胞壁組分半纖維素的含量,解吸半纖維素上吸附的鐵來提高體內的有效鐵含量。
擬南芥體內的一系列轉運蛋白不僅負責根系對土壤中鐵的吸收,還和體內鐵的長距離運輸密切相關。鐵在木質部運輸主要以鐵—檸檬酸復合物的形式存在,前人已經報道的AtFRD3家族蛋白可以參與這一過程,遺傳學證據分析,Atfrd3突變體在鐵充足情況下葉片黃化,地上部有效鐵含量顯著降低但是根系卻積累大量的鐵,過表達Atfrd3后能夠回補這種缺鐵癥狀,說明AtFRD3蛋白確實參與了鐵由根系向地上部運輸的過程[40]。關于韌皮部運輸,AtNAS1和AtYSL2轉運蛋白被認為是在鐵韌皮部運輸中起著重要作用。當在擬南芥中敲除AtNAS1和AtYSL2后,地上部鐵含量大幅度下降,并且伴隨著幼葉出現脈間失綠的現象,暗示著AtNAS1和AtYSL2可以調控擬南芥體內鐵的動態平衡過程[41-42]。在本次試驗中發現,在缺鐵的條件下外源添加硼后,尤其是添加1000 μmol/L H3BO3后,擬南芥根系中AtFRD3、AtNAS1和AtYSL2的基因表達量都顯著高于正常的缺鐵條件,說明硼可以通過調控以上3個基因的表達來促進體內的鐵從根系向地上部運輸的過程。
4 結論
在缺鐵的條件下,外源添加硼可以通過改變擬南芥體內細胞壁半纖維素含量來促進細胞壁鐵的解吸,從而增加根系和地上部的有效鐵含量。通過相關基因表達量分析,調控鐵運輸過程的幾個關鍵轉運蛋白AtFRD3、AtNAS1和AtYSL2參與并能促進鐵從根系向地上部轉運的過程,而信號分子NO能夠參與這一過程并最終緩解植物的缺鐵癥狀。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
