冷誘導金黃色葡萄球菌環境脅迫耐受性及生物被膜控制
2019-04-01 06:31:10,,,,,,
食品工業科技 2019年5期
關鍵詞:生物
,,,,, ,
(南京農業大學食品科技學院,江蘇南京 210095)
近年來,我國速凍食品微生物安全受到廣泛關注,有餡食品如速凍餃子在生產、儲藏、銷售過程中受微生物污染的可能性最大,而金黃色葡萄球菌(Staphylococcusaureus)是速凍食品中常檢出的食源性致病菌[1-2]。研究表明,金黃色葡萄球菌在-18 ℃冷藏96 d過程中,亞致死率增加但細胞死亡率卻逐漸變緩,說明亞致死細胞對冷脅迫耐受性較強[3-5]。金黃色葡萄球菌可以通過冷休克蛋白(CSP)的表達而有效地適應溫度的降低,這種冷休克反應對金黃色葡萄球菌在低溫下生存起了重要作用[6]。Onyango等[7]研究發現,金黃色葡萄球菌在4 ℃可形成小菌落突變體,其生長形態和超微結構發生改變且對氨基糖苷類藥物耐受性增強。冷脅迫環境可使金黃色葡萄球菌蛋白組代謝組發生變化[8]。
雖然關于食源性致病菌對多種環境脅迫產生交叉抗性的報道已有很多。但對于金黃色葡萄球菌經冷誘導后對其他環境耐受性的研究較少。腸炎沙門氏菌在5 ℃脅迫5 h后,其對酸性環境的耐受性增強[9]。Cebrián等人研究發現在低濃度的酸、堿、過氧化氫以及45 ℃溫度條件下處理一定時間的金黃色葡萄球菌,對更極端的同源環境耐受性增強[10]。Singh等[11]發現對酸具有強適應性的沙門氏菌能在60 ℃脅迫條件下保持較高存活率。金黃色葡萄球菌能在工作臺和器皿表面形成生物被膜,會對外界刺激因素如高溫高鹽、強酸強堿、紫外線、抗生素、消毒劑、殺菌劑等產生抗性[12-13]。
為了控制金黃色葡萄球菌在冷凍食品中的污染傳播,本文研究了低溫敏感性不同的金黃色葡萄球菌經冷誘導后對其他環境脅迫(高溫、酸、堿、高滲、消毒劑)的抗性變化,同時研究了冷凍食品分離株生物被膜的形成和有效控制方法,為冷凍食品工業安全控制提供理論支持。
1 材料與方法
1.1 材料與儀器
金黃色葡萄球菌(S.aureus,CICC10201) 中國工業微生物菌種保藏管理中心;金黃色葡萄球菌(S.aureus,W3) 分離自市售冷凍魚丸樣品;胰蛋白胨大豆肉湯培養基(TSB)、營養肉湯培養基(NB) 北京陸橋技術有限公司;氯化鈉、30%過氧化氫、乙醇 分析純,南京化學試劑有限公司;84消毒液、雕牌洗潔精 南京蘇果超市。
1300-A2生物安全柜 賽默飛世爾科技公司;AR512電子精密天平 奧豪斯(上海)公司;LDZX-50 KBS立式壓力蒸汽滅菌鍋 上海申安醫療器械廠;WH-3微型旋渦混合儀 上海滬西分析儀器廠;HYL-A全溫搖瓶柜 太倉市強樂實驗設備廠;5418高速離心機 Eppendorf;DK-8D型電熱恒溫水槽 上海森信實驗儀器有限公司;隔水式電熱恒溫培養箱 上海躍進醫療器械廠;BCD-215YD海爾冰箱 海爾集團。
1.2 實驗方法
1.2.1S.aureus耐冷凍能力的研究 分別取10 μLS.aureusCICC10201及S.aureusW3于4 mL離心管37 ℃ 180 r/min過夜培養,調整菌液終濃度為OD595=0.5后,以1%(v/v)接種量接種到新鮮培養基中,37 ℃ 180 r/min培養5 h。將所得菌液分裝于1.5 mL/管離心管中,凍藏于-20 ℃冰箱內。每隔7 d取一個凍藏的離心管測定其菌落數,測定8周。
1.2.2S.aureus的冷誘導S.aureusCICC10201及S.aureusW3經TSB液體培養基過夜培養后,調整菌液終濃度為OD595=0.5,按1%(v/v)接種量接種至50 mL新鮮培養基中,37 ℃ 180 r/min振蕩培養5 h后,6000 r/min離心5 min,用2 mL滅菌生理鹽水重懸菌液后,分別置于4 ℃冰箱中,冷脅迫5 h。
1.2.3 冷誘導S.aureus環境脅迫抗性研究
存活率(%)=脅迫前細菌數/脅迫后細菌數×100
式(1)
降低的細菌數(lg CFU/mL)=lg(脅迫后細菌總數)-lg(脅迫前細菌總數)
式(2)
1.2.3.1 高溫脅迫 將2 mL冷誘導的S.aureusCICC10201及S.aureusW3菌體細胞懸液分別置于45、50、55、60 ℃水浴處理30 min后,快速置于冰上冷卻30 s,平板稀釋涂布,測定細菌數計算存活率,計算方法見式(2)。
1.2.3.2 酸脅迫 取2 mL未經誘導及冷誘導后菌株S.aureusCICC10201及S.aureusW3菌懸液,6000 r/min離心5 min,去上清,得到的菌體分別重懸于2 mL高壓滅菌的pH為2.5、3.5、4.5、5.5的HCl溶液中室溫靜置1 h[11]。平板稀釋涂布,統計活菌數并計算存活率。
1.2.3.3 堿脅迫 取2 mL未經誘導菌株及冷誘導后菌株的菌懸液(冷誘導時間增加至24 h),6000 r/min離心5 min,去上清,得到的菌體分別重懸于2 mL高壓滅菌的pH為8.5、9.5、10.5、11.5的NaOH溶液中室溫靜置1 h。平板稀釋涂布,統計活菌數并計算存活率。
1.2.3.4 高滲透壓脅迫 取2 mL未經誘導菌株及冷誘導后菌株的菌懸液,6000 r/min離心5 min,去上清,得到的菌體分別重懸于2 mL高壓滅菌的25%、30%、35%氯化鈉溶液中,模擬S.aureus可能遇到的高滲環境,室溫靜置1 h,平板稀釋涂布,統計活菌數并計算存活率。
1.2.3.5 酒精脅迫 取2 mL未經誘導菌株及冷誘導后菌株的菌懸液,6000 r/min離心5 min,去上清,得到的菌體分別重懸于2 mL濃度為10%、20%、30%、40%的酒精溶液中處理5 min。平板稀釋涂布,測定細菌數并計算存活率。
1.2.3.6 過氧化氫脅迫 取2 mL未經誘導菌株及冷誘導后菌株的菌懸液,6000 r/min離心5 min,去上清,得到的菌體分別重懸于2 mL用30%過氧化氫 與無菌水配制的10、15、20、25 mmol/L 過氧化氫溶液中室溫靜置30 min,后平板稀釋涂布,統計活菌數并計算存活率。
1.2.4 生物被膜形成能力的變化及控制
1.2.4.1 菌株的挑選 以NB培養基為基礎,使用96孔板結晶紫染色法[14]衡量兩株分離株S.aureusCICC10201及S.aureusW3的生物被膜形成能力。菌株按1%(v/v)接種量接種到96孔微量細胞培養板中,每孔含菌懸液200 μL。每個樣品設6個重復,同時12孔不接菌的原始培養基為對照。37 ℃培養24 h后棄去培養基,無菌水洗3次,加入200 μL結晶紫染色30 min。倒去染色液,使用無菌水洗去浮色,晾干后每孔加200 μL 95%乙醇,室溫放置15 min[13]。用酶標儀在595 nm處測得樣品OD值,選取OD值最大的菌株進行下一步實驗。
1.2.4.2 消毒劑和清潔劑對S.aureus生物被膜的清除作用 取培養24 h的96孔板,棄去培養基,無菌水洗3次后,在不同孔中分別加入無菌水(對照),不同濃度的84消毒液(1.00%、0.50%、0.25%),洗潔精(2%、5%、10%),過氧化氫(0.5%、1.5%、3.0%),酒精(20%、40%、80%)浸泡處理30 min,倒出浸泡液,用無菌水清洗3次,后續實驗同1.2.4.1。
1.3 數據處理
每個實驗重復3次,實驗數據經SPSS軟件進行單因素方差分析,結果用Excel 2007軟件繪圖。
2 結果與討論
2.1 冷凍對S. aureus存活率的影響
如圖1,隨著冷凍時間的增長,標準菌株存活率顯著低于分離株(p<0.05)。S.aureusW3經-20 ℃ 8周冷藏后,細菌數發生變化不明顯,對于冷脅迫具有較強的耐受性。而對于標準菌株S.aureusCICC10210,對-20 ℃耐受能力,較冷脅迫不同時間的分離株均差,凍藏8周后,菌體數下降2.27 lg CFU/mL,存活率不到1%。S.aureus已被證明能在多種環境條件下生存,在極端環境中通過改變細胞代謝和結構維持生存狀態[15]。冷凍食品分離株S.aureusW3較標準菌株S.aureusCICC10201在-20 ℃環境下的存活率更高,體現了其極強的抗冷凍脅迫調節能力。這意味著一旦速凍食品污染了耐冷菌株,發生食品安全事故的概率較高。

圖1 -20 ℃長期冷藏對S. aureus菌體數的影響Fig.1 Effect of low temperature(-20 ℃)on the cell number of S. aureus
2.2 冷誘導對菌株環境脅迫抗性的影響
2.2.1 冷誘導后S.aureus對高溫耐受性的變化 如圖2A,45 ℃下耐冷凍菌株S.aureusW3具有較高存活率,未經冷誘導前存活率高達98.12%,顯著高于對照標準菌株S.aureusCICC10201(58.93%)(p<0.05)。而經冷誘導后,兩株菌的存活率都有所上升,分別達到114.5%(S.aureusW3)及77.35%(S.aureusCICC10201)。50 ℃水浴處理,冷誘導前S.aureusW3存活率依然高達88.36%,而標準菌S.aureusCICC10201僅為37.11%。經冷誘導后,存活率增長,分別達94.14%和55.61%。隨著溫度升高,不同處理組的兩株菌的存活率都快速下降,冷誘導前S.aureusW3在55 ℃高溫下,存活率達9.2%,而標準菌存活率為5.23%。由于在60 ℃處理0.5 h后,各組菌存活率均不足1%,細菌數降低量都在3 lg CFU/mL左右,無顯著差異(p>0.05)(圖2B)。由此可見,冷誘導可使菌株對高溫的耐受性增強,而耐冷菌株較標準株的高溫耐受性更強。已被證實暴露在亞致死溫度(冷誘導)中對高溫具有更強耐受性的菌種有大腸桿菌[16]、單核細胞增生李斯特氏菌[17]、小腸結腸炎耶爾森菌[18]和植物乳桿菌[19]。因此在冷凍食品工業中,應考慮冷誘產生的細菌高溫抗性,建立高效的殺菌措施,保證耐冷菌株完全滅活。

圖2 冷誘導前后S. aureus在高溫脅迫下的存活率Fig.2 Survival rate of S. aureus at high temperature before and after cold induction注:A:兩株菌冷誘導前后對高溫存活率變化;B:60 ℃處理后各組降低的細菌數。*代表差異顯著,p<0.05,圖3~圖8同。
2.2.2 冷誘導S.aureus對酸耐受性的變化 酸對去除蔬菜、肉制品表面食源性致病微生物有良好的效果[20-21]。由圖3可知在pH3.5~5.5條件下,四組存活率極高,均超過80%,分離株與標準株存活率未有顯著差異(p>0.05)。在室溫下pH4.5、pH5.5處理1 h后,未經冷誘導的兩株菌均有增殖現象,存活率超過100%,經冷誘導后,兩株菌在弱酸條件下的存活率都有所下降。在pH5.5條件下,冷誘導后的兩組菌存活率分別下降20.0%和21.4%。在pH4.5條件下,冷誘導后S.aureusW3存活率下降不顯著(p>0.05),但標準株S.aureusCICC10201存活率下降16.7%。在pH3.5條件下,冷誘導后兩組菌存活率分別下降18.0%和16.8%。在較低pH條件下(pH=2.5),標準菌株的存活率(17.7%)顯著高于分離株(3.22%)(p<0.05)。以上結果表明,冷誘導S.aureus對弱酸環境的敏感性增強,同時分離株對pH2.5環境下的耐受性低于標準菌株。有研究表明,不同溫度條件下乙酸和檸檬酸對芝麻醬中S.aureus存活率有影響。在低溫(10 ℃)條件下比37 ℃條件下S.aureus對酸的耐受能力更強[22],這可能與酸種類、處理方式不同有關。本研究采用的酸處理為無機酸,而文獻報道中所用的是有機酸。另有研究證實[9],5 ℃ 5 h誘導的鼠傷寒沙門氏菌產生酸抗性,使其在pH4.0條件下存活率提高。這可能與陰性菌和陽性菌對酸反應不同有關,實際生產過程中也需注意可能存在交叉抗性的情況。

圖3 冷誘導前后S. aureus在酸性脅迫下的存活率Fig.3 Survival rate of S. aureus before and after cold induction under acid stress
2.2.3 冷誘導S.aureus對堿耐受性的變化 如圖4,冷誘導5 h后,pH9.5S.aureusW3與pH8.5S.aureusCICC10201的存活率無顯著差異(p>0.05)。通過增加冷誘導時間至24 h,觀察冷誘導后兩株菌對堿脅迫耐受性變化。隨著誘導時間的加長,兩株菌的存活率均有所提高。S.aureusW3在誘導5、24 h后在pH8.5的條件下存活率分別提高了12.44%和16.84%。在pH9.5條件下存活率分別提高了3.9%和18.4%。且在較高pH條件下(pH10.5),存活率提高更為顯著(p<0.05),為18.6%和33.8%,冷誘導前在該pH條件下不能生長。對于標準菌株S.aureusCICC10201,在冷誘導后其對堿耐受性也有略微增長。雖然分離株在pH8.5及9.5的條件下存活率較標準株低,但在pH10.5的條件下存活率顯著高于標準株(p<0.05)。國內外對于冷誘導后S.aureus堿耐受性提高的報道較少。僅有研究表明,S.aureus在pH9.5的環境中暴露0.5 h會使部分細菌產生堿適應性,這與蛋白質及RNA的合成有關[23]。

圖4 冷誘導前后S. aureus在堿脅迫下的存活率Fig.4 Survival rate of S. aureus beforeand after cold induction under alkaline stress注:A為S. aureus W3冷誘導前后對堿脅迫存活率變化;B為S. aureus CICC10201冷誘導前后對堿脅迫存活率變化。
2.2.4 冷誘導S.aureus對氯化鈉耐受性的變化 食品環境及S.aureus侵襲感染過程中都可能遇到高滲環境。S.aureus對氯化鈉耐受性較高,可能與其細胞壁合成和鹽應激蛋白有關[24-25]。隨著氯化鈉濃度的增加,各組存活率逐漸降低(圖5),但冷誘導前耐冷菌S.aureusW3與標準菌的存活率無顯著差異(p>0.05)。25%(w/v)氯化鈉處理后,經冷誘導的兩株菌存活率下降,分別降低了9.2%和20.3%。在30%(w/v)氯化鈉處理后,經冷誘導的兩株菌存活率分別降低24.09%和22.1%。在35%(w/v)氯化鈉處理后,經冷誘導后的兩株菌存活率分別下降13.08%和9.1%。冷誘導后高滲環境的存活率顯著下降(p<0.05),這可能與冷誘導造成的部分S.aureus細胞亞致死有關,使其對氯化鈉的耐受性降低。

圖5 冷誘導前后S. aureus在氯化鈉脅迫下的存活率Fig.5 The survival rate of S. aureus before and after cold induction under hypertonic condition
2.2.5 冷誘導S.aureus對酒精耐受性的變化 如圖6A,經10%、20%酒精脅迫,兩株菌的存活率均超過50%,且誘導前后無顯著差異(p>0.05)。10%酒精處理后,各組存活率均達80%以上。但經20%酒精脅迫后,標準株較分離株S.aureusW3存活率高。而在超過30%酒精濃度下,菌體大量死亡,特別是在40%酒精濃度下,已檢測不到活菌存在。根據30%酒精處理菌株的降低的細菌數(圖6B)可知,未經誘導的兩株菌菌體數分別下降了4.95和4.86 lg CFU/mL,而經誘導后的兩株菌菌體數下降了3.03和2.79 lg CFU/mL,存活率顯著提高(p<0.05)。由此可得,經冷誘導后的S.aureus對30%濃度的酒精產生交叉抗性。國內外對于冷誘導后S.aureus對酒精耐受性提高的報道較少。但有學者研究了其他環境因素誘導食源性致病菌產生酒精抗性。如Chiang等[26]研究發現,酸適應性副溶血性弧菌對8%的酒精耐受性提高。

圖6 冷誘導前后S. aureus在酒精脅迫下的存活率Fig.6 Survival rate of S. aureus before and after cold induction under alcohol stress注:A為經酒精處理后不同組的存活率;B為在30%酒精處理后各組降低的菌體數。
2.2.6 冷誘導S.aureus對過氧化氫耐受性的變化 過氧化氫是食品加工過程中常用的器械消毒劑,其殺菌機理是產生活性氧作用于細胞DNA、脂質和蛋白。如圖7所示,耐冷菌株S.aureusW3對雙氧水的耐受性高于標準菌株S.aureusCICC10201,在各濃度處理后,其存活率均高于50%。經冷誘導后,S.aureusW3對過氧化氫的耐受性變化不顯著(p>0.05),而標準菌株在冷誘導后存活率顯著(p<0.05)降低,對過氧化氫的敏感性增強。有研究表明,sigB基因與過氧化氫抗性的產生相關,特殊的調節子例如PerR也可能調控S.aureus應激作用[27]。
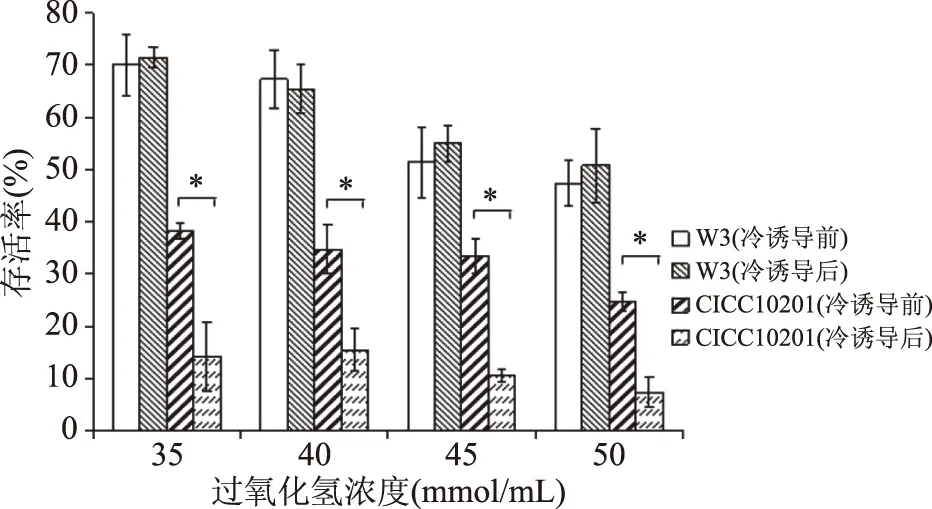
圖7 冷誘導前后S. aureus在過氧化氫脅迫下的存活率Fig.7 Survival rate of S. aureus under hydrogenperoxide stress before and after cold induction
2.3 生物被膜形成能力的變化及控制
生物被膜是微生物抵抗外界不良生活環境的一種特殊生長方式[28-29]。S.aureus具有一定粘附性,因此可在器物表面及食品表面[30]形成生物被膜,生物被膜較游離菌體更難被清除。結果表明S.aureusW3的生物被膜OD595值為2.7,而S.aureusCICC10201的值為0.5,因此S.aureusW3作為冷凍食品分離株比標準具有更強的生物被膜形成能力。
運用不同濃度的洗潔精(w/v)、84消毒水(v/v)、酒精(v/v)和雙氧水(v/v),對分離株S.aureusW3形成的生物被膜進行清除(圖8)。各梯度稀釋的洗潔精對S.aureusW3形成的生物被膜均有一定清除作用,與未添加洗潔精相比,其吸光度均有顯著下降(p<0.05)。不同濃度的洗潔精之間均有顯著差異(p<0.05)。84消毒液(主要成分為次氯酸鈉)對S.aureusW3形成的生物被膜也有一定清除作用,與對照相比,經30 min處理后各實驗組生物被膜吸光度均顯著下降(p<0.05)。1%的84消毒水對其生物被膜的清除作用好于10%的洗潔精。酒精對其生物被膜清除沒有明顯效果(p>0.05)。0.5%~3%的過氧化氫對其生物被膜也有一定清除作用,但效果較高濃度的洗潔精和84消毒水差。對于該菌,1%的84消毒水能夠有效地清除生物被膜,酒精幾乎沒有清除作用。
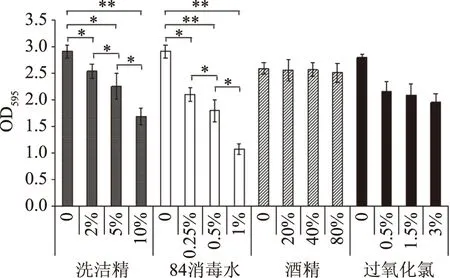
圖8 不同消毒劑對分離菌S.aureus W3生物被膜的清除作用Fig.8 Scavenging effect of different scavengers on S.aureus W3 biofilm of cold resistant bacteria注:** 代表差異極顯著,p<0.01。
3 結論
冷凍食品中分離得到的S.aureusW3較標準菌株S.aureusCICC10201在-20 ℃環境下的存活率更高,經8周冷凍后,細菌數幾乎沒有下降,體現了其極強的抗冷凍脅迫調節能力。
S.aureusW3對高溫(50 ℃)、堿(pH10.5)、過氧化氫的耐受性均較標準菌株S.aureusCICC10201更好,但對酸(pH2.5)敏感性較高。冷誘導會使S.aureusW3和S.aureusCICC10201對于高溫、堿、酒精產生交叉抗性。同時,冷誘導會使這兩株菌對某些壓力環境的敏感性增強。如經冷誘導后,對酸、氯化鈉及過氧化氫的生存能力顯著下降。1%的84消毒水可有效去除冷凍食品分離株S.aureusW3形成的生物被膜。以上研究表明,利用冷誘導后S.aureus對酸、高滲及過氧化氫敏感性增強的特點,可有效控制S.aureus在冷藏食品工業中的污染和傳播。
猜你喜歡
天天愛科學(2022年9期)2022-09-15 01:12:54
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
科學大眾(2021年9期)2021-07-16 07:02:54
軍事文摘(2020年20期)2020-11-28 11:42:50
航空世界(2020年10期)2020-01-19 14:36:20
小學科學(學生版)(2018年3期)2018-04-18 12:34:19
知識經濟·中國直銷(2017年10期)2017-11-07 02:39:52
知識經濟·中國直銷(2017年3期)2017-04-16 03:08:01
文學少年(有聲彩繪)(2017年2期)2017-03-06 08:10:54
