四川華鳊精子發生顯微及超微結構觀察
2019-04-08 07:39:14解崇友王志堅
淡水漁業 2019年2期
尹 敏,解崇友,金 麗,王志堅
(西南大學生命科學學院,淡水魚類資源與生殖發育教育部重點實驗室,水產科學重慶市重點實驗室,重慶 400715)
四川華鳊(Sinibramataeniatus)屬鯉形目(Cypriniformes)鯉科(Cyprinidae)鲌亞科(Cultrinae)華鯿屬(Sinibrama),僅分布于長江上游岷江、青衣江、嘉陵江以及大渡河等水域[1],是長江上游珍稀特有小型魚類。近年來,長江上游干流及主要支流大規模修建電站,攔河筑壩,致其生境尤其是繁殖環境遭到不同程度破壞,持續的高強度捕撈壓力也使得四川華鳊種群數量銳減,解崇友等[2]的研究發現,四川華鳊的種群已出現小型化趨勢,因此有必要開展四川華鳊種質資源保護相關工作。目前關于四川華鳊的研究資料主要集中在資源調查[3]、年齡生長[2]以及繁殖生物學[4]等,未見室內人工繁殖的相關研究。人工繁殖是實現漁業資源保護和自然種群恢復、增殖的重要技術手段,而生殖細胞發生作為繁殖生物學的重要組成部分,了解其在生殖周期中的動態變化,對于掌握物種繁殖規律具有重要意義。本研究以四川華鳊成魚為研究對象,通過性腺組織學切片,輔以透射電鏡技術,對其精子發生過程進行顯微及超微結構的觀察,豐富其繁殖生物學基礎資料,為人工繁殖提供科學指導。
1 材料與方法
1.1 實驗材料
岷江眉山段采集的性成熟四川華鳊,在實驗室條件下暫養兩周后開始取材,每月一次。養殖條件:自然光照,水溫(20±2) ℃,pH值7.5±0.5,溶解氧>7 mg/L,每日9∶00投喂浮性顆粒飼料。
1.2 實驗方法
解剖取四川華鳊成魚精巢,顯微觀察采用Bouin氏液固定24 h,梯度酒精脫水,二甲苯透明,石蠟包埋,Leica RM 2235切片機切片5 μm,H.E染色[5],Nikon Eclipse 80 i顯微鏡觀察并拍照;超微結構觀察采用預冷的4%戊二醛固定,經0.1 mol/L磷酸緩沖液(pH值7.4)沖洗3次,1%鋨酸后固定1~2 h,雙蒸水清洗3次(每次不超過3 min),梯度酒精脫水(50%乙醇,15 min;70%乙醇+醋酸雙氧鈾過飽和溶液,過夜;90%乙醇,10 min),丙酮置換(90%乙醇+90%丙酮1∶1,10 min;90%丙酮,10 min;100%丙酮,10 min,3次),環氧樹脂650聚合包埋,LBK-5型超薄切片機切片,醋酸超氧鈾和枸櫞酸鉛雙染,JEM-1400 Plus透射電鏡觀察并拍照。
1.3 數據處理
使用圖像分析軟件Image-Pro Plus 6.0測量生殖細胞直徑及核徑,SPSS 17.0對數據進行統計分析。
2 結果
2.1 生殖細胞形態學指標
光學顯微鏡及透射電鏡下隨機選取不同標本的不同區域進行拍照,統計結果見表1。

表1 四川華鳊雄性生殖細胞大小統計表Tab.1 Statistics of male germ cell sizes in S. taeniatus μm
2.2 生殖細胞發生的顯微及超微結構
2.2.1 精原細胞
精原細胞在成魚性腺的各個時期中都能找到,精原細胞沿精小葉分布,一般在精巢邊緣分布較多(圖版I-1)。精原細胞為精子發生過程中體積最大的細胞,H.E染色后,精原細胞著色最淺,可分為兩種類型,分別稱A型精原細胞和B型精原細胞。A型精原細胞為精原干細胞,細胞卵圓形,細胞長徑(9.80±0.62) μm,細胞核也為卵圓形,長徑(6.28±0.50) μm,核膜清晰,核仁明顯,位于細胞核中央或偏向一側,嗜堿性強,染成藍色。核質中有極少的嗜堿性物質也著藍色,細胞質與細胞膜均不著色,因此細胞邊界不清晰但高倍鏡下基本能分清細胞界限(圖版I-2)。A型精原細胞分裂產生B型精原細胞。B型精原細胞稍大,長橢圓形,細胞直徑的長徑為(11.02±1.13) μm,細胞核長徑為(7.42±0.82) μm,光學顯微鏡下B型精原細胞核仁不明顯,核內嗜堿性染色質分散于核膜邊緣或核質內各處(圖版I-2)。
兩種類型的精原細胞在顯微結構上只能根據核仁大小及核膜是否清晰加以區分,而超微結構顯示,二者除了在核仁大小、電子密度以及核膜上的區別外,其胞質中擬染色質小體的數量、線粒體的多少等也是區分兩種細胞的重要依據。A型精原細胞的細胞膜及核膜清晰,中央大核仁電子密度高,核內染色質均一,核膜外周有多個擬染色質小體,核內有與其電子密度相似的顆粒物質,胞質中線粒體豐富,內嵴清晰,透射電鏡下整體呈棒狀或橢圓形(圖版II-7)。B型精原細胞其核膜較前者模糊,核仁松散,電子密度較低,核質均一,核膜外周擬染色質小體較少,胞質中線粒體明顯減少。兩種類型的精原細胞在與相鄰細胞接觸的位置均出現了細胞膜的缺口,猜測為細胞質橋(圖版II-8)。
2.2.2 初級精母細胞
B型精原細胞經有絲分裂生長形成初級精母細胞,初級精母細胞主要存在于Ⅱ期和Ⅲ期精巢中,在較晚時期的精巢中數量不多。初級精母細胞長徑(8.08±0.46) μm,細胞核長徑(4.95±0.58) μm,光學顯微鏡下只能觀察到初級精母細胞的細胞核,細胞邊界幾乎不可見,與精原細胞相比,細胞核質內染色質凝縮,絲狀或團塊狀不均一分布,胞核嗜堿性增強,H.E染色后呈淺藍色,核膜清晰,細胞核形態不一,部分初級精母細胞有中央大核仁,呈深藍色,部分初級精母細胞無明顯的中央大核仁(圖版I-3)。
超微結構中觀察到的初級精母細胞為不規則長橢形,細胞核圓形,胞質中細胞器較多,核質比增大不明顯,仍能觀察到核膜的雙層膜結構,核膜開始出現不同程度波曲,核質電子密度較精原細胞大,核內染色質團塊狀不均勻分布,核膜外有擬染色質小體存在,膨大線粒體附著在擬染色質小體附近,胞質中也有許多線粒體分布,可觀察到內質網,未觀察到具中央大核仁的初級精母細胞,在初級精母細胞群附近觀察到了橋粒連接(圖版II-9)。
2.2.3 次級精母細胞
初級精母細胞在減數第一次分裂后形成次級精母細胞,次級精母細胞大量存在于III期、IV期及V期早期的精巢中,其體積進一步減小,細胞長徑(4.75±0.49) μm,約為初級精母細胞的一半,細胞核長徑(3.75±0.24) μm,核質比明顯增大,核內染色質進一步凝縮,高倍鏡下可觀察到核質線團狀散亂分布,核質嗜堿性增強,整個細胞核染成較均一的藍色(圖版I-4)。
超微結構中觀察到的次級精母細胞呈卵圓形,細胞核形態與初級精母細胞相似,近圓形,細胞膜的雙層膜結構不清晰,但可根據電子密度區分細胞核與胞質,核內染色質均一,電子密度增大,核膜附近未見擬染色質小體,胞質內仍分布有大量線粒體,但內嵴不明顯,此外,胞質內還有許多電子密度較大的顆粒狀物質。同樣,次級精母細胞間也有細胞質橋存在(圖版II-10)。
2.2.4 精子細胞
次級精母細胞通過減數第二次分裂產生精細胞,在Ⅳ期的精巢中較易觀察到精細胞,精細胞長徑(3.81±0.38) μm,較次級精母細胞進一步縮小,核長徑(2.94±0.36) μm,光學顯微鏡下觀察到精細胞的細胞核圓形,較精母細胞體積明顯減小,只比精子稍大,細胞整體呈深藍色(圖版I-5)。

圖版I 四川華鳊精子發生的顯微結構(H.E 染色)Plate.I Microstructure of Sinibrama taeniatus spermatogenesis
電鏡下觀察到的精細胞有多種形態,根據核質濃縮程度及細胞形態結構特征,可將精細胞產生到成熟精子形成之前的過程大致分為三個階段,早期的精細胞體積較次級精母細胞稍小,細胞間開始出現明顯縫隙,但仍有少量的細胞質橋連接,核質電子密度稍有增加,核質比增大,胞質減少并向一側集中,胞質內可見多個膨大的線粒體,高爾基體結構明顯,大部分精細胞在精子頭部正下方的胞質中有一空腔,內有形似微管的結構,猜測為基體的橫切面。在結構完整的早期精細胞中,觀察到互相垂直的基體和近端中心粒,已有鞭毛沿基體伸出(圖版II-11)。中期的精細胞體積進一步減小,沒有多余胞質,精子頭部基本形成,細胞邊緣可觀察到多層膜結構,由細胞膜及核膜緊貼形成,細胞核進一步濃縮,核質密度明顯增大,可觀察到核空泡,高爾基體和膨大的線粒體位于胞核后端兩側的胞質內,胞核正下方為內含基體結構的腔(圖版II-12)。晚期的精細胞中,由基體延伸出的鞭毛仍位于胞質下方的空腔內,未見結構清晰的近端中心粒,但在基體上方靠近核膜的位置有中心粒附屬物的形成(圖版II-13)。
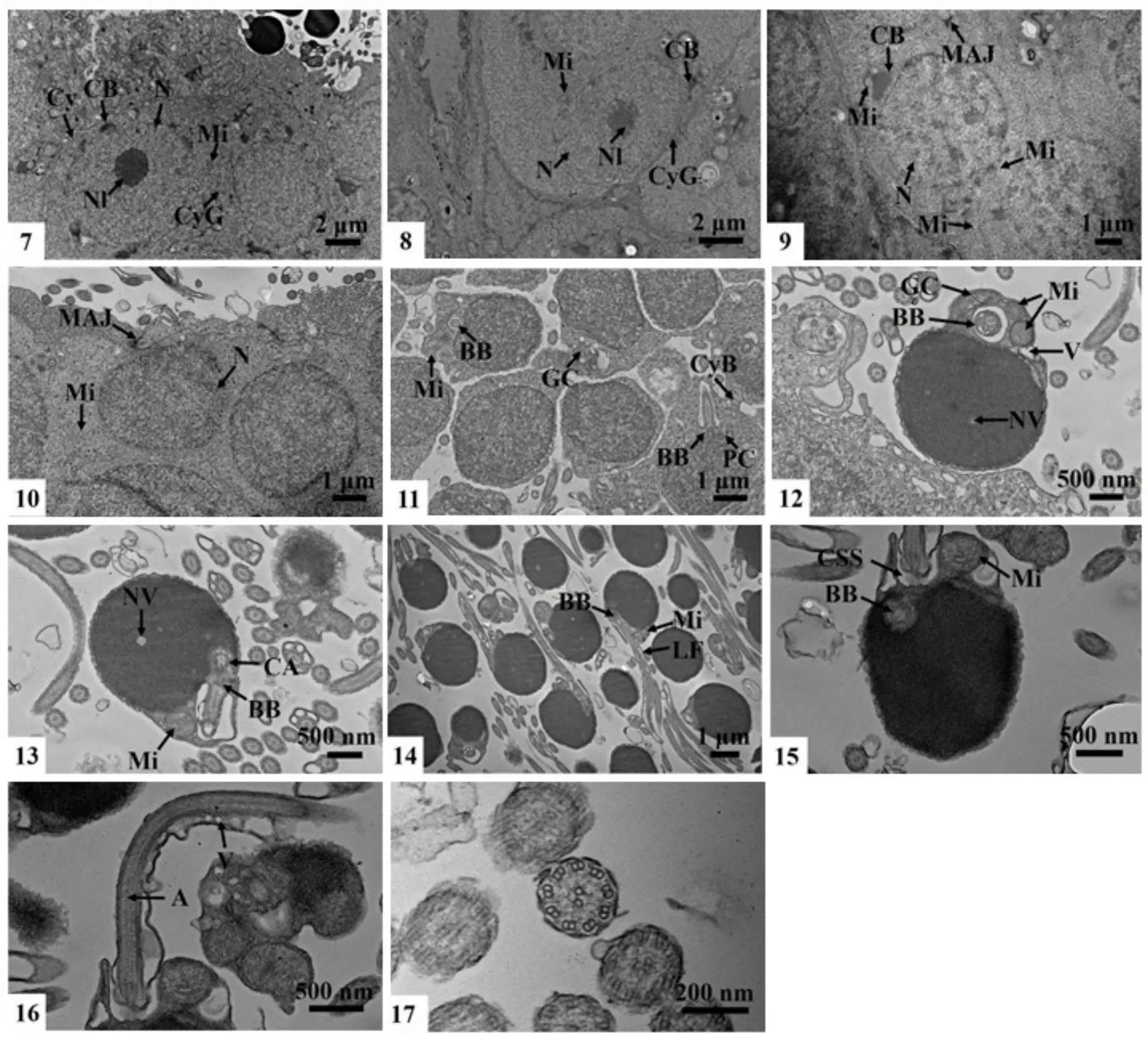
圖版II 四川華鳊精子發生的超微結構Plate.II Ultrastructure of Sinibrama taeniatus spermatogenesis
2.2.5 精子
精子細胞經變態發育為成熟精子,精子大量存在于Ⅳ期及Ⅴ期精巢的小葉腔中,在早期的精巢中也可觀察到少量的成熟精子。精子嗜堿性極強,為染色最深的生殖細胞,呈深藍色(圖版I-6)。
電鏡下觀察到的精子分三部分(圖版II-14):精子頭部近圓形或稍不規則,無頂體,長徑(2.42±0.02) μm,核長徑(2.28±0.05) μm,絕大部分為細胞核,電子密度極大,電鏡下呈黑色,精子頭部切面可觀察到核空泡,但較小,核凹窩不發達(圖版II-15);精子中片緊靠精子頭部下端核膜,由精子尾部著生位置的中心粒復合體和袖套共同構成,在成熟精子中,中心粒復合體中最明顯的結構為基體,未見中心粒及其附屬物,精子尾部著生位置胞質凹陷形成袖套腔,腔內有膨大的線粒體及一些囊泡化的細胞器結構,精子尾部由軸絲構成,貫穿精子細長尾部,形成精子運動的支撐結構,精子鞭毛橫切可觀察到軸絲由外圍9組二聯體微管及中央2根獨立微管共同組成,精子尾部兩側有側鰭,基本對稱,精子尾部后端的側鰭中有囊泡存在(圖版II-15,16,17)。
3 討論
四川華鳊精子發生過程經歷了精原細胞、初級精母細胞、次級精母細胞以及精子細胞變態的過程。A型精原細胞通過有絲分裂形成B型精原細胞,B型精原細胞不斷進行有絲分裂,產生能夠進入減數分裂的初級精母細胞,初級精母細胞經兩次減數分裂形成精細胞,再經變態形成成熟精子。在這個過程中,除兩次減數分裂以外,生精細胞的細胞核及細胞質結構均發生了一系列變化。
精原細胞核中最顯著的結構為中央大核仁,但B型精原細胞的核仁更為松散。核仁與蛋白質的旺盛合成、細胞的活躍生長有關[6],核仁一般在分裂前期消失,分裂末期重新出現。在四川華鳊的兩種精原細胞中均觀察到了核仁,而在顯微結構中只有部分初級精母細胞可觀察到明顯核仁。康現江等[7]發現半滑舌鰨的初級精母細胞核仁在減數第一次分裂前期的偶線期消失,與本研究的結果基本相符,但沒有說明核仁是否在減數第一次分裂末期重建,可能由于減數第一次分裂完成后停留時間太短,不易觀察到該階段的生精細胞。四川華鳊精子發生過程中,初級精母細胞之后的生精細胞中也未觀察到明顯核仁。
在超微結構中,從精原細胞分裂開始,處于同一精小囊內的生精細胞間就出現了細胞質橋,具體表現為相鄰細胞在接觸位置細胞膜出現了明顯缺口,這種現象直至發育到精細胞早期才結束。這與同一精小囊內生精細胞的同步發育有關,這與車靜[8]對大眼鱖精子發育的研究結果一致,而管汀鷺等[9]在對金魚精子的研究中,將這一結構稱為細胞間橋,并認為細胞間橋直至精子成熟前才消失。然而,關于細胞質橋的具體定義,不同的研究者持有不同觀點。洪萬樹等[10]在對鯔魚精子發生的超微結構觀察中發現,初級精母細胞有絲分裂過程為不完全胞質分裂,胞質之間出現合胞體,從而導致細胞界限不明顯,不同的是,他們僅將精細胞間的連通結構稱為細胞間橋。郭申明等[11]也提到,在孔雀魚的精子發生過程中,從精母細胞到精子細胞其胞質均有不完全分裂現象,但未將這樣的結構稱為細胞質橋或細胞間橋。不管稱為合胞體的胞質連接還是細胞質橋,這一現象的存在都是精小囊內生精細胞維持高效同步發育的前提。大量研究表明,細胞質橋在各種生物精子發生過程中具有普遍性,從低等動物如帶絳蟲[12]、人皮蠅[13]等到哺乳動物如大鼠[14]以及人的精子發生過程中,生精細胞的分裂過程始終伴隨著相鄰細胞胞質的溝通。
在四川華鳊精子發生過程中,兩種類型的精原細胞以及初級精母細胞的核膜外側均存在體積較大、數量不等的擬染色質小體,同時在精原細胞核內觀察到了與其電子密度相似的顆粒,猜測擬染色質小體的形成與核內的這些高密度顆粒物質有關。根據管汀鷺[15]對金魚精子發生中擬染色質小體的研究,細胞核內物質聚集形成致密體,排入胞質后形成擬染色質小體,在次級精母細胞以后,核內不再形成致密體,胞質內的擬染色質小體被消耗,數量和體積減小,這可以解釋為什么在次級精母細胞及以后的細胞中很難觀察到擬染色質小體。Walt等[16]認為擬染色質小體內存在肌動蛋白,與高爾基體共同作用參與頂體蛋白的形成,并參與RNA代謝。
精子變態過程中細胞質偏向一側,細胞核濃縮,在這一過程中,中心粒復合體起著至關重要的作用。四川華鳊精子細胞變態早期就已出現近端中心粒和基體,變態中期出現了中心粒附屬物,而成熟精子中只觀察到有基體的存在。根據林丹君等[17]對褐菖鲉精子的描述,中心粒復合體在精子細胞晚期發生變化,中心粒附屬物形成,在精子成熟前,中心粒附屬物萎縮直至消失,近端中心粒也在精子成熟時消失,這與我們的觀察結果基本一致。Fawcett等[18]認為,中心粒附屬物可能與哺乳類精子頸段的形成有關,但尚未有研究提到其在硬骨魚類精子形成過程中的作用。王志堅等[19]對長薄鰍精子發生的研究中,認為遠端中心粒(基體)向細胞后部移動導致胞質被拉長,近端中心粒向細胞核方向遷移導致核質進一步濃縮,形成核凹窩。四川華鳊精子的核凹窩極淺,核空泡小,側面說明其精子變態過程中近端中心粒向細胞核方向遷移程度不大,核質濃縮程度較小,這一特性在其精子頭部的形態上也有所體現。四川華鳊精子頭部與草魚[20]、刀鱭[21]、泥鰍[22]等相似,而與長吻鮠[23]、胭脂魚[24]等核凹窩極深的精子形態不同。研究顯示,同為鰍科的泥鰍[22]、大鱗副泥鰍[25]、寬體沙鰍[26]三種魚類精子頭部均為圓形,核凹窩淺,鲿科鮠屬的短尾鮠[27]、粗唇鮠[28]的精子頭部形態則與長吻鮠相似,核凹窩都較深,張濤等[29]對同屬四種羅非魚精子的對比研究顯示,其精子頭部結構大體相同,推測精子頭部的形態特征可能具有種屬特異性,或可作為魚類分類鑒定的一項指標及精子結構預測的依據,但這一猜想尚待進一步驗證。
與絕大多數體外受精魚類[30-32]精子一樣,四川華鳊精子為無頂體型,尾部具兩側對稱的側鰭。側鰭的有無、發達程度以及形態結構在不同硬骨魚中具有差異性。目前的研究表明,許多硬骨魚精子尾部均具側鰭,為胞質延伸形成。胭脂魚[24]、黃顙魚[33]精子尾部側鰭發達,兩側基本對稱,鯔魚[10]精子尾部側鰭亦為兩側對稱,但發達程度不如四川華鳊精子。江鱈[34]、銀鯧[35]精子尾部側鰭不對稱,泥鰍[22]、叉尾鯰[36]精子尾部無側鰭。關于精子尾部側鰭的生理意義,不同研究者持有不同意見:Stoss[37]認為側鰭的存在能提高精子的運動速度,而尹洪濱等[38]通過對不具側鰭的索氏六須鯰精子的研究后,認為精子運動速度主要取決于鞭毛,側鰭只起到部分作用。但目前尚未有研究者對不同類型精子做過對比研究,要明確側鰭的具體作用還需進一步研究。
