外源NO對重樓種子活力及超弱發光的影響
2019-04-09 08:32:50李海明李梅香孟艷艷宋發軍戴甲培李澤華
廣東農業科學 2019年2期
李海明,李梅香,孟艷艷,宋發軍,戴甲培,李澤華,耿 紅
(1.中南民族大學生命科學學院/生物技術國家民委重點實驗室/武陵山區特色資源植物種質保護與利用湖北省重點實驗室,湖北 武漢 430074;2.中南民族大學武漢神經科學與神經工程研究所,湖北 武漢 430074)
【研究意義】重樓(Paris)為百合科重樓屬(Trilliaceae)多年生草本植物,分布于云南、廣西、江西、浙江、湖南等地[1],其正品藥源包含七葉一枝花(Paris polyphyllavar.chinensis,PPC)和滇重樓(Paris polyphyllavar.yunnanensis,PPY)[2]。重樓的根狀莖經濟價值很高,是云南白藥等藥的重要組分之一,近年來受到了掠奪式采挖,導致重樓的野生資源銳減[3]。利用種子進行人工繁殖是緩解供需矛盾的有效途徑,已得到人們的廣泛關注。然而重樓種子的休眠期長且休眠程度高、萌發速度慢、萌發率較低。自然條件下,重樓種子一般需要兩冬一夏才能開始萌發,15個月的出苗率僅為46.2%,漫長的休眠期和萌發周期會使大量種子的生活力下降甚至喪失生命力[4]。因此,隨著市場對重樓種子需求的不斷增加,重樓種子質量的高低、種子活力的大小和種子處理方式直接影響人工栽培重樓的產量和質量。目前,國內外對重樓種子休眠原因和解除休眠方法、種子繁育技術等方面展開了系列研究[5-8],而關于重樓種子質量檢驗技術和生活力檢驗技術等方面的研究較少,重樓種子超弱發光方面的研究更是未見報道。生物的超弱發光是生物體進行新陳代謝和體內生理生化反應過程中細胞自發輻射的非常微弱的光子流,無論是動植物還是單細胞生物,其體內都會有超弱發光的現象[9]。其中,種子的超弱發光通常由自由基和有絲分裂產生。自由基主要是由于種子代謝活動產生,而有絲分裂與細胞生長分裂、細胞內核酸的含量變化有關[10-12]。因此,種子的超弱發光不僅可以反映種子的生命活動狀態、細胞內外的新陳代謝和信息交換的程度,還可以作為檢測種子活力的一個重要指標。【前人研究進展】目前,國內對種子萌發過程中自發超弱發光的研究較少,尤其是對于包含重樓在內的具有深度休眠特性的藥用植物種子而言,報道更是少見。現有研究表明,種子內部的生理生化反應、細胞活動等任何生成或利用ATP、NADPH、NADP的反應均與種子的自發超弱發光有著密切聯系[13]。其原因可能是在這些代謝反應中,細胞內的部分能量以光子的形式釋放出來,所以反應越劇烈,細胞活動越活躍,種子的自發超弱發光的強度就越高。種子的超弱發光可以借助不同的儀器設備測定。田茜等[14]研究了人工老化過程中超弱發光的變化及其與種子活力、ATP含量的關系。王春芳等[15]利用BPCL-2微弱發光測量儀完成了稻谷和小麥種子發芽率檢測。趙燕燕等[16]利用自制的測量系統對高粱種子萌發過程中的超弱發光特性進行了研究。而利用新型的超弱發光圖像檢測裝置可以達到更加直觀、準確和實時檢測種子的超弱發光變化情況的目的。【本研究切入點】選用本單位自行搭建的電子倍增式CCD(EMCCD)生物光子成像系統,對不同類型和不同處理的重樓種子活力進行檢測。該檢測系統具有超高靈敏性,在對極微弱光信號的實時快速探測方面,可真正做到單光子事件的檢測[17],并且此裝置可實時監測生物的不同部位及生物在不同代謝階段或不同環境下的超弱發光及其變化情況。已有研究表明,NO作為一種活性分子,在適宜濃度下可以促進多種植物種子的萌發,并提高其萌發率[18-25]。我們前期研究也發現,適宜濃度的外源NO供體硝普鈉(sodium nitroprusside,SNP)可以顯著縮短種子的萌發周期,且提高重樓種子的萌發率[26]。【擬解決的關鍵問題】分別以七葉一枝花和滇重樓的種子為研究對象,通過檢測SNP處理和對照的種子在萌發期間的超弱發光強度,建立重樓種子超弱發光值與其萌發率的關系,分析SNP對重樓種子超弱發光的影響,并建立通過超弱發光值預測種子生活力的模型,為重樓種子質量和發芽率的快速判定提供基礎。
1 材料與方法
1.1 試驗材料
供試材料為中南民族大學巴東實驗基地采摘的3年生七葉一枝花與滇重樓的成熟蒴果,經萬定榮教授鑒定。去穎殼和果皮,并以人工揉搓的方法去除紅色漿果的外種皮和果肉,得到乳白色較堅硬的種子。經自來水充分洗滌后室溫下晾干,存于4 ℃備用,具體方法參照文獻[27]。試劑為購自TIANDZ的亞硝基鐵氰化鈉(SNP)。
主要儀器:探測儀EMCCD(日本Hamamatsu公司,ImagEMCCDC9100-13);專用暗箱(定做,上海歐海化工機械設備有限公司);變焦透鏡(DO5095,美國Navitar公司,F stop值為1. 0)。
1.2 試驗方法
1.2.1 浸種處理 挑選健康飽滿的重樓種子,七葉一枝花與滇重樓各兩份,每份120顆,分裝于250 mL錐形瓶中,25 ℃條件下用蒸餾水浸種48 h,至種子發白為止。將分析純級別的SNP配成100 μmol/L溶液,用該溶液浸種為48 h,以蒸餾水浸種為對照。
1.2.2 接種處理 浸種完成后,在超凈工作臺將所有種子用蒸餾水清洗5遍,用無菌水清洗3遍,并用75%乙醇清洗30 s并不停搖晃,再用無菌水清洗3遍。接著用0.1%升汞處理種子12 min,處理過程中不停搖晃錐形瓶。最后將升汞處理過的種子用無菌水清洗5遍。并轉移至鋪有2層無菌濾紙的培養皿中進行暗培養,每皿120粒,溫度19(±2)℃。
1.2.3 觀察記錄 種子培養期間每隔7 d補1次水,補水量均為1 mL,保證培養皿內的種子濕潤。培養過程中記錄種子的形態變化。以胚根剛剛突破種皮為標準,統計種子培養0、10、20、30、40、50、60、70 d的發芽率(Gr),計算發芽指數(Gi):

式中,Gt培養t天的種子萌發數,Dt為萌發天數。
1.2.4 種子超弱發光值測定 重樓種子的自發超弱發光強度測定參照宋述燕[17]方法進行。不同處理重樓種子分別放置在專用暗箱中,種子超弱光子信號經連接在EMCCD的變焦透鏡成像在EMCCD探測器上,然后經光電效應轉換成電子圖像,再用計算機對圖像進行分析。培養后0、10、20、30、40、50、60、70 d,每份種子各取樣9粒,每3粒1組,共測定3組重樓種子樣品在萌發期間的自發超弱發光強度,3次重復,取平均值。光值采集參數設置均為:曝光時間60 ms,間隔0.3 s,增益1 200,持續采集90 min,試驗過程中不存在外部漏光的可能。數據處理時刪減前30 min的發光值,以除去自然延遲發光對實驗結果的影響。
利用SPSS16.0和Graphpad prism 7進行數據分析和作圖。
2 結果與分析
2.1 不同處理重樓種子萌發過程形態特征
由圖1可知,滇重樓種子經100 μ mol/L SNP處理30 d時即有胚根突破種皮,而對照種子培養50 d后才有胚根突破種皮。SNP處理的種子萌發時間比CK提前20 d,萌發培養70 d時SNP處理種子的胚根明顯長于CK。此外,SNP處理的七葉一枝花和滇重樓種子均比CK提前20 d萌發。

圖1 不同處理重樓種子萌發過程中的形態變化Fig.1 Morphological changes in Paris seed germination process of different treatment groups
2.2 不同處理重樓種子發芽率和發芽指數
從表1可以看出,SNP處理的七葉一枝花和滇重樓種子均在培養30 d發生萌發現象,即胚根開始突破種皮,萌發時間分別比CK提前20 d。萌發培養60 d,SNP處理的兩組種子萌發數均已超過50%,而對照種子的萌發率僅有25%左右。萌發培養70 d,兩種重樓種子在SNP處理的萌發率都達到90%,分別比對照高36.41%、47.5%。此外,SNP處理的滇重樓和七葉一枝花種子發芽指數也明顯高于CK。結合圖1與表1發現,外源NO具有促進種子萌發、提高種子活力和改善種質的作用,且這種作用在七葉一枝花中表現更為明顯。

表1 不同處理重樓種子的發芽率Table 1 The seeds germination rate of Paris with different treatments
2.3 不同處理重樓種子超弱發光強度變化
重樓種子萌發過程中體內自發超弱發光強度變化如圖2所示。由圖2A可知,種子培養0~30 d為萌發前期(種子培養開始到胚根開始突破種皮),處理和對照七葉一枝花種子的發光強度無顯著差異。在萌發中期(胚根突破種皮到胚根的長度達到1.5 cm,30~60 d),SNP處理中,七葉一枝花種子的自發超弱發光值迅速升高,萌發40~60 d,SNP處理種子的發光強度顯著高于對照,在萌發后期(胚根長度>1.5 cm,60~70 d)又下降,種子發光強度最高峰值出現在萌發40 d。而對照種子在不同萌發時間節點上發光強度變化不大;在萌發中期,種子發光強度略有降低;萌發后期,又有小幅度抬升。滇重樓種子和七葉一枝花種子的發光強度變化趨勢相似。其中SNP處理種子的發光值也在萌發40 d明顯升高且顯著高于對照,萌發50~60 d其發光強度也高于對照但差異不顯著;其他時間點的發光強度對比中,兩組差異均不顯著(圖2B)。未經SNP處理情況下,七葉一枝花和滇重樓的種子在不同時間段的發光強度無明顯差異。與此同時,SNP處理后七葉一枝花種子和滇重樓的種子發光強度的變化趨勢相同,但是在萌發50 d與萌發60 d時,七葉一枝花種子的發光強度均明顯高于滇重樓的。
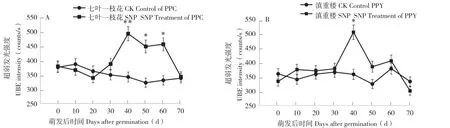
圖2 不同處理重樓種子的自發超弱發光強度Fig. 2 The spontaneous ultra-weak luminescence intensity of Paris seeds with different treatments
2.4 不同處理重樓種子的發光強度與發芽率變化趨勢
七葉一枝花對照在種子培養40 d后,發芽率為0,發光強度無明顯變化。培養40~60 d種子開始萌發,此時發光強度有輕微下降。培養70 d,CK種子大量萌發,與此同時該階段種子的發光強度有上升趨勢(圖3A)。SNP處理中,七葉一枝花種子在培養20 d時發芽率為0、發光強度變化波動較小,培養30~40 d種子開始萌發、發光強度明顯上升,隨著萌發種子數增加(50~60 d),發光強度保持較高水平,隨后在培養70 d時下降(圖3B)。滇重樓對照(圖3C)與七葉一枝花的對照變化趨勢相同,滇重樓的SNP處理(圖3D)的變化趨勢也與七葉一枝花SNP處理中種子的變化類似。
3 討論
NO作為細胞中的一種重要信號分子,參與植物體內如防御機制和抗逆反應等多個生理過程[18-25]。此外,有研究發現硝普鈉(SNP)對亞麻[28]、玉米[29]、九節木[30]和蒺藜苜蓿[31]等種子的萌發也有一定促進作用。我們前期研究也發現,適當濃度的外源NO可以大幅減短重樓種子的萌發周期,且提高種子的萌發率[26]。然而,外源NO對重樓種子活力及萌發期間種子超弱發光的影響還未見報道。因此我們從自然超弱發光的角度,探討了自然條件下和SNP處理后重樓種子萌發過程中發光強度的變化。
結果表明,SNP處理種子的萌發形態變化和萌發效果均明顯早于CK,表明SNP在一定程度上提升了種子活力,達到促萌作用。此外,不同試驗的重樓種子,在萌發后0~30 d內種子內部自發超弱發光無顯著差異,但在萌發后40~60 d SNP處理明顯高于CK,隨后差異不顯著。其原因可能是由于在種子萌發前期,重樓種子正在適應萌發條件,克服種子休眠的生理現象。因此該時間段內,種子內部的生理生化反應與細胞活動均較少。SNP處理后,短時間內可能不會對種子發光現象產生影響,存在滯后期。種子萌發40 d開始,SNP處理中種子內部的生理生化反應與細胞活動加劇,種子萌發數迅速增加,該時期種子的自發超弱發光強度也最大。隨著新增萌發種子數的減少,種子內部的有絲分裂等生理活動逐漸減弱,種子自發超弱發光強度也逐漸降低,與對照差別不大。對照重樓種子的自發超弱發光強度最大值的出現時間均與新增萌發種子數最高值的出現時間相吻合。
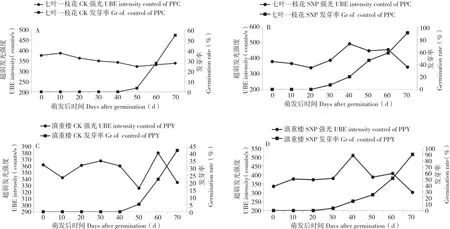
圖3 不同處理重樓種子發光強度與發芽率的關系Fig. 3 The relation between luminescence intensity and germination rate of Paris seeds with different treatments
已有研究表明,小麥種子的發芽率與發光強度呈線性相關關系,而水稻種子的發芽率與發光強度符合非線性相關關系[15]。白菜與蘿卜新陳種子的研究表明,新種子的發芽率與發光強度呈正相關,但陳種子的發芽率與發光強度呈負相關[32]。因此,了解種子發光強弱可為判定種子活力、預測種子發芽率提供理論依據。但從本試驗發芽率與超弱發光強度的比較可以看出,無論是對照還是處理的種子萌發率與發光強度沒有相同的變化趨勢,均不呈明顯的線性關系,這與小麥和蘿卜等的研究結果不同。原因可能是上述4種類型的種子均為淺休眠型,而重樓種子在萌發過程中是深度休眠,且具有形態休眠和生理休眠的雙重特性,從而使重樓種子發光強度的變化趨勢呈現峰狀變化。另外,不同試驗處理方式不同,也會造成較大差異。本試驗取樣時間為種子萌發進程的前70 d,在此時段內SNP處理組種子的萌發程度已達90%,但對照只有50%種子萌發。在自然條件下,種子萌發進程第70天后的發芽率與發光強度是否有相關性還未知,有待深入研究。
4 結論
綜上所述,本試驗采用的EMCCD能有效檢測種子萌發過程中的自發超弱發光強度,可準確得到待測種子二維光強分布的空間與時間信息,據此判斷種子活性的強度,并建立有效的通過超弱發光值預測種子生活力的模型,為解決重樓種子質量和發芽率的快速判定問題提供基礎。EMCCD對重樓種子二維光強進行檢測,表明自發超弱發光能給種子的萌發進程提供重要相關信息,預示對光子的研究技術在農業科研上有廣闊的利用前景。另外,我們從形態觀察、發芽率、發芽指數和發光強度等方面綜合得出,SNP對改善重樓種子萌發質量有明顯作用,在種子萌發40~60 d時效果尤為明顯。其改善機制是否因為提高了重樓種子體內的NO等信號分子的濃度,從而加快了種子內部的生理生化反應或促進了種子的萌發進程還有待進一步探索。
