雜草對ALS抑制劑抗藥性概述
2019-04-26 01:13:14黃兆峰王園園姜翠蘭周欣欣
農藥科學與管理 2019年2期
黃兆峰,劉 倩,王園園,姜翠蘭,周欣欣
(1.中國農業科學院植物保護研究所,北京 100193;2.北京市園林科學研究院,北京 100102;3.駐馬店市農業技術推廣站,河南 駐馬店 463000;4.農業農村部農藥檢定所,北京 100125)
雜草降低了作物產量和品質,增加了農民投入,制約了農業的持續發展,是農業生產中的一大類生物災害[1]。目前,化學除草是當前農業生產中最主要的除草手段。除草劑防除雜草相對于人工除草具有高效、省工等優點,為農業生產帶來了巨大的經濟效益,但是隨之帶來抗藥性雜草的發生與發展等負面問題日趨嚴重。近年來,抗藥性雜草生物型數量近乎呈直線上升的趨勢,若不加以控制,抗性雜草演變為農田優勢種,不僅加重農田雜草的危害,也會加大對農田雜草的防除難度。抗藥性雜草的發生和快速發展成為我國糧食增產的制約因素之一,這對以化學除草為主的雜草綜合治理體系發起了挑戰。目前,關于雜草抗藥性機制的研究已經成為近年來雜草科學研究熱點。從生態經濟學角度科學防控雜草,合理使用除草劑,延緩雜草抗藥性的發生與發展,保障雜草的有效防控以及農業生態系統的可持續發展。
1 雜草抗藥性現狀
第二次世界大戰期間,MCPA和2,4-D的發現和使用,拉開了人類化學除草的序幕。除草劑的使用顯著降低了田間工作量,并且提高了作物的產量。在保障農業增效、糧食增產、農民增收做出了積極的貢獻。同時,由于長期使用同一種或者多種但作用方式或代謝降解途徑相同的化學除草劑,導致抗藥性雜草不斷發生和發展。雜草抗藥性問題越來越嚴重,已經影響了糧食生產,受到了全球廣泛的關注[2]。1950年,在美國和加拿大發現抗2,4-D的鋪散鴨跖草(Commelinadiffusa)和野胡蘿卜(Daucuscarota)標志著抗藥性雜草的產生。隨后,歐洲千里光(Seneciovulgaris)對三氮苯類除草劑西瑪津和莠去津產生抗藥性也被報道[3]。抗藥性雜草不斷出現與發展,截止到2018年12月,全球范圍內,有255種雜草(單子葉雜草107種和雙子葉雜草148種)的496個雜草生物型對不同作用方式的除草劑產生了抗性(表1)[4]。我國田園雜草有1 400多種,已報道有27種共43個生物型的抗藥性雜草(表2)。

表1 全球抗藥性雜草生物型

續表

表2 中國所報道的抗藥性雜草
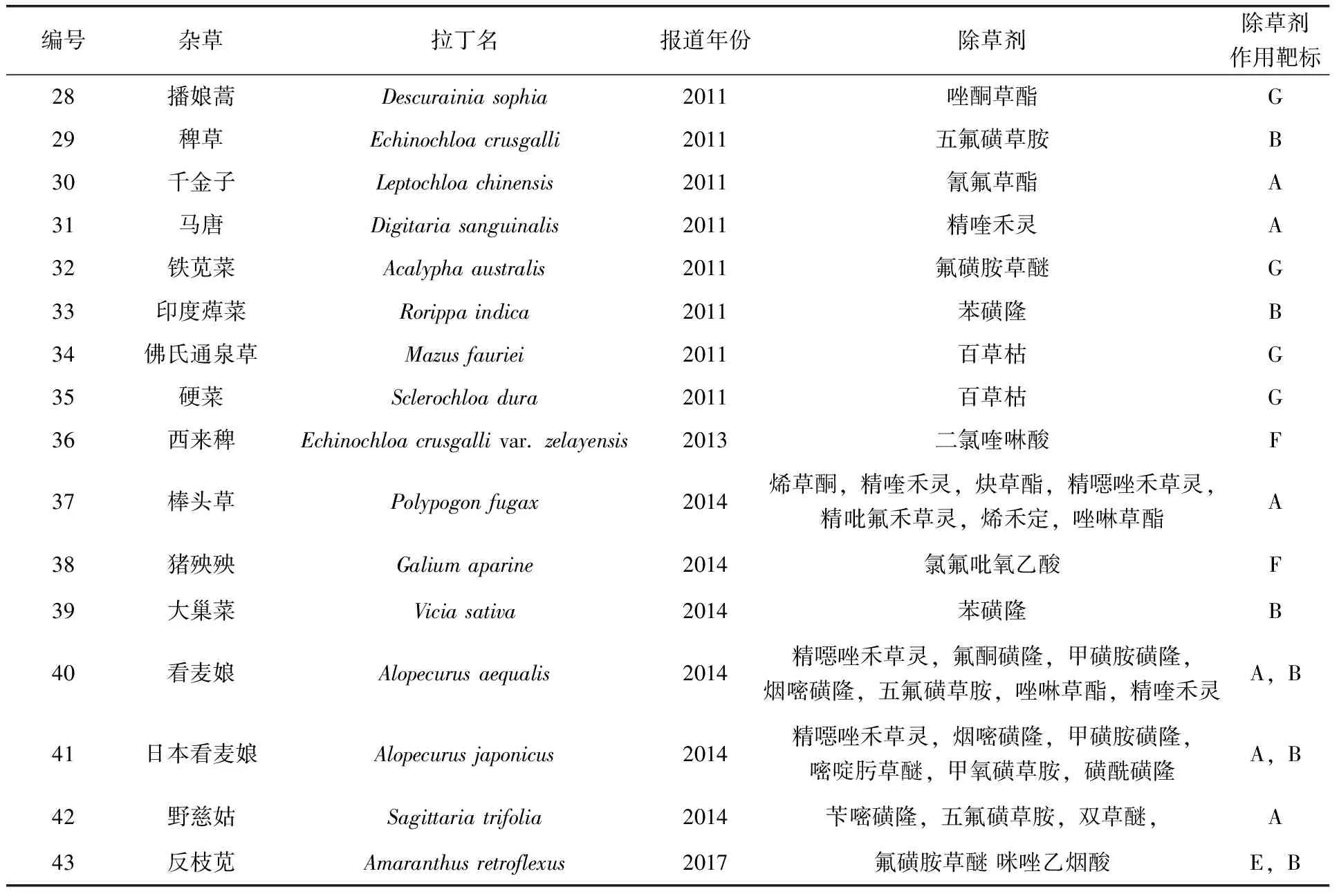
續表
注:A:乙酰輔酶A 羧化酶抑制劑;B:乙酰乳酸合成酶抑制劑;C:光合系統II抑制劑;D:5-烯醇丙酮莽草酸-3-磷酸合酶抑制劑;E:類脂合成抑制劑 ;F:合成激素類抑制劑;G:其他
雜草對ALS抑制劑抗藥性概況 乙酰乳酸合成酶(Acetolactate synthase,簡稱ALS,EC 2.2.1.6),廣泛存在于植物、真菌、細菌以及藻類等生物體中[5]。它是合成支鏈氨基酸,如亮氨酸、異亮氨酸、纈氨酸的關鍵酶[6]。ALS抑制劑是指以ALS酶為靶標的除草劑。現己開發的ALS抑制劑種類較多,主要包括磺酰脲類(sulfonylureas,SU)、咪唑啉酮類(imidazolinones,IMI)、嘧啶硫代苯甲酸酯類(pyrimidinylthiobenzoates,PTB)、三唑并嘧啶類(triazolopyrimidines,TP)和磺酰胺羰基三唑啉酮類(sulfonylaminocarbonyltriazolinone,SCT)。雜草受藥后,除草劑與ALS酶相結合使其活性被抑制,支鏈氨基酸合成被阻,最終導致死亡[7]。由于具有高效、低毒、對環境污染小等優點,該類除草劑迅速成為全球應用最為廣泛和最重要的除草劑之一。自1982年美國杜邦公司注冊登記首例ALS抑制劑除草劑氯磺隆以來,全球現已開發ALS抑制劑類除草劑50余種,其中SU類除草劑商品化品種近40種[8]。但由于作用位點單一,抗ALS抑制劑雜草發生速度極快。截止到2018年12月,對ALS抑制劑產生抗性的有160個雜草生物型[4],約占總數的32%,涉及禾本科、菊科、十字花科、莎草科、雨久花科等十幾個科。
2 雜草對ALS抑制劑的抗性機理
雜草對ALS抑制劑類除草劑的抗藥性可以分為靶標抗性和非靶標抗性兩方面。其中,靶標ALS基因突變或代謝作用增強導致抗藥性占主要部分。
2.1 靶標ALS基因突變 研究表明,高等植物的ALS氨基酸序列上存在5個不連續的高度保守區域,導致雜草抗藥性的產生的氨基酸突變位點主要有8個,且均在這5個保守區內。這些位點表達的氨基酸主要有:Ala122、Pro197、Ala205、Asp376、Arg377、Trp574、Ser653、Gly654(相對于擬南芥ALS)[9~10]。靶標基因保守區域發生錯義突變,改變靶標酶蛋白結構,致使靶標酶對除草劑的敏感性下降,使除草劑不能與靶標酶有效結合,除草活性不能很好的發揮,這是到目前為止報道最多的抗性機制。
一般情況下,靶標基因保守區域不同位點突變所造成的交互抗藥性類型不一[11](Yu & Powles,2014)。比如,Ala122和Ala205氨基酸突變一般會導致對IMI類除草劑有抗性;Pro197位氨基酸突變會導致對SU類除草劑有抗性,由于雜草種類和突變類型的差異,也會對TP類除草劑和SCT類除草劑有交互抗性,對IMI類除草劑敏感或者有較低水平抗性;Asp376位氨基酸突變一般會導致對五類ALS抑制劑有廣譜交互抗性;Trp574位氨基酸突變會導致對SU類和IMI類除草劑都有抗性,這是因為Trp574殘基決定著ALS活性位點通道的形狀,而且在ALS與SU類和IMI類除草劑結合中起到很重要的作用;Ser653位氨基酸突一般會造成對IMI類除草劑有抗性,也會對PTB類除草劑有抗性;Gly654位氨基酸突變一般會造成對IMI類除草劑有抗性(表3)[11~12]。

表3 部分雜草ALS突變位點與交互抗性
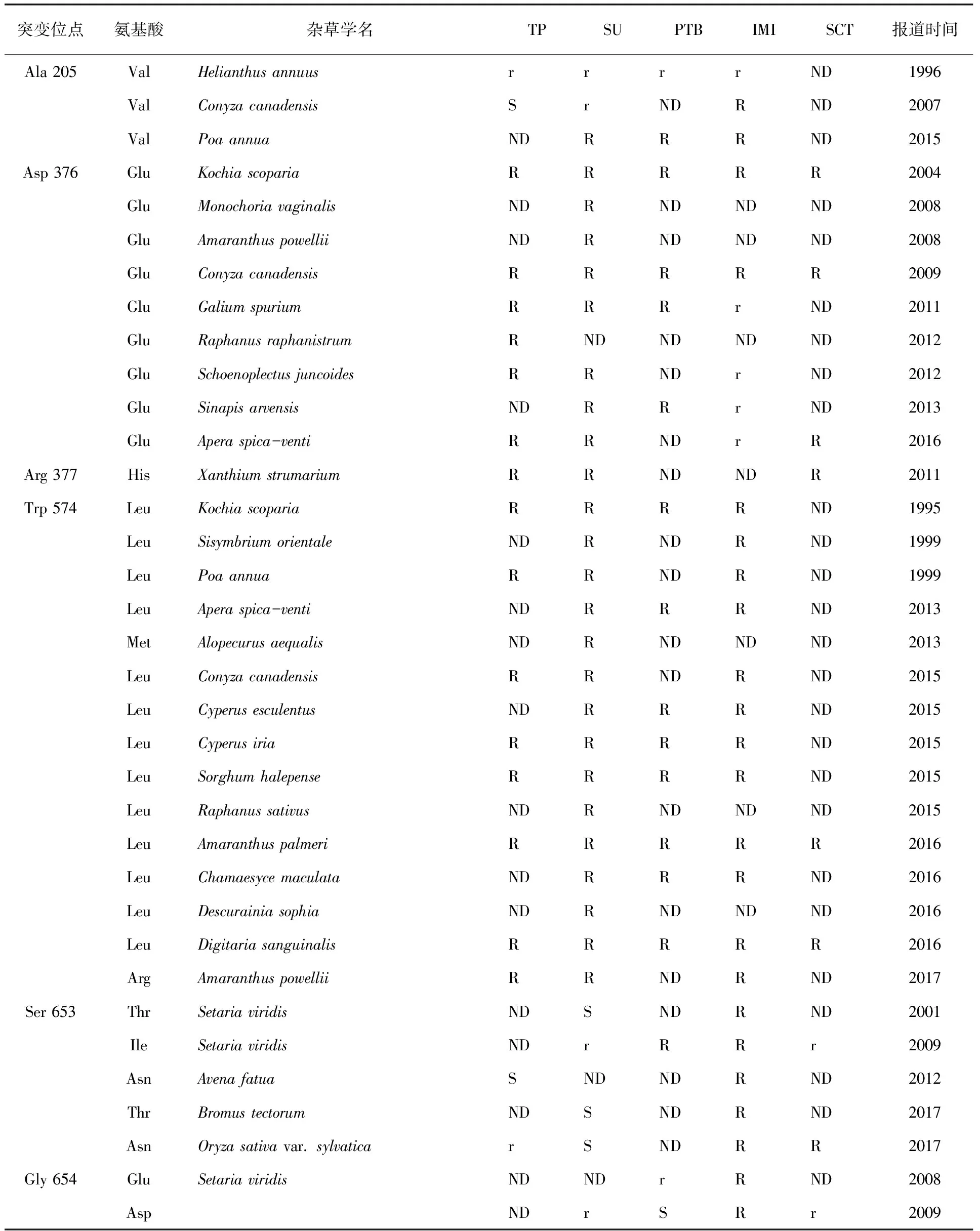
續表
2.2 代謝作用增強 植物體對 ALS 抑制劑的代謝作用增強是產生抗藥性一個重要原因。在植物體內,細胞色素P450單加氧酶(P450s)、谷胱甘肽還原酶(GSTs)、超氧化物歧化酶(SOD)等解毒代謝酶類可提高將除草劑代謝為無毒化合物的能力,導致雜草抗藥性的發生[12]。P450s 介導的雜草對除草劑的抗藥性研究主要集中在禾本科雜草上[11],如硬直黑麥草(Loliumrigidum)、鼠尾看麥娘(Alopecurusmyosuroides)、旱雀麥(Bromustectorum)、水稗(Echinochloaphyllopogon)、野燕麥(Avenasterilis)。研究硬直黑麥草對氯磺隆的抗性機制,結果顯示抗藥性生物型體內細胞色素P450s含量或活性的提高,是導致抗藥性發生的原因[13]。研究旱雀麥(BromustectorumL.)對丙苯磺隆的抗性機制,結果顯示細胞色素 P450s 提高對除草劑的代謝能力是導致抗藥性發生的原因[14]。研究水稗對雙草醚的抗性機制,結果表明:細胞色素P450s對除草劑的代謝能力的增強是導致抗藥性產生的原因[15]。其中,硬直黑麥草、鼠尾看麥娘和水稗是研究非靶標代謝抗性最多的禾本科雜草。在闊葉雜草方面,研究有關P450s介導產生非靶標抗性研究較少。主要有白芥菜(Sinapisarvensis)、綠穗莧(Amaranthushybridus)、糙果莧(Amaranthustuberculatus)。
植物的P450s基因家族龐大,多數由300~400個基因所構成,分離鑒定單個P450s基因的作用較為困難[16]。Iwakami等[17]報道,從抗雙草醚水稗鑒定的兩個P450基因CYP81A12和CYP81A21,轉到擬南芥和酵母后,表達的CYP81A12和CYP81A21酶可代謝除草劑。研究黑麥草的代謝抗性發現,CYP72A和CYP81A1基因表達差異顯著,使得黑麥草降解除草劑水平提高[16]。
3 結語
化學除草劑是機械化、規模化作物種植模式不可缺少的除草措施,對于糧食安全具有重要的意義。除草劑也是目前和未來很長時間田間除草的主要手段,由此帶來的抗性雜草問題不可避免。由于過度依賴除草劑,抗藥性雜草不斷發生發展,必須采取有效措施阻止或延緩雜草抗藥性發生和蔓延,延長除草劑使用年限,確保糧食安全[18]。我國雜草抗藥性研究起步較晚,研究技術和儀器設備相對落后,研究力量相對薄弱。鑒于目前我國農業逐步向集約化、規模化和現代化方向發展,必須針對我國雜草抗藥性現狀,特別是抗ALS抑制劑類雜草,避免ALS抑制劑的高風險抗性問題。需借鑒國外先進的抗藥性研究及監控技術,建立抗藥性雜草預測預報、快速檢測、抗性雜草的綜合治理體系,在雜草治理中要科學合理地應用除草劑,避免連續使用同一種或同一類作用機制的除草劑;充分發揮農藝措施,通過合理的輪作改變田間雜草群落的組成。延緩雜草抗藥性的發生。
猜你喜歡
科教新報(2022年22期)2022-07-02 12:34:28
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
今日農業(2019年15期)2019-01-03 12:11:33
現代園藝(2017年19期)2018-01-19 02:50:21
營銷界(2015年23期)2015-02-28 22:06:18
現代農業(2015年5期)2015-02-28 18:40:49
中國質量與標準導報(2013年8期)2013-03-11 19:54:07
