HvSAPK3基因在‘鳳大麥7號’抗白粉病中的功能研究
2019-05-05 01:15:02李慶偉宗英杰劉成洪陸瑞菊許建華黃劍華
上海農業學報 2019年2期
關鍵詞:分析
李慶偉,宗英杰,郭 嬌,劉成洪,陸瑞菊,許建華,黃劍華*
(1上海海洋大學水產與生命學院,上海201306;2上海市農業科學院,上海201106;3光明種業有限公司,上海 202171)
為了應對洪澇、干旱、冷、鹽和病蟲害等各種環境脅迫,植物在長期的進化過程中形成了復雜的抗逆機制。蛋白質的磷酸化和去磷酸化是細胞中極其重要的一種蛋白質翻譯后調控方式,與植物應答環境脅迫信號機制有關[1]。SnRK蛋白是一種高度保守的SerThr蛋白激酶,在植物中廣泛存在。SnRK蛋白激酶可以通過磷酸化修飾靶蛋白來調控植物應答脅迫中的多種信號途徑,在植物脅迫應答過程中起著至關重要的作用[2]。研究表明,根據蛋白結構差異,SnRK家族的38個成員可劃分為SnRK123三個亞家族;其中SnRK1蛋白與酵母SNF1蛋白以及哺乳動物AMPK蛋白存在著較高的序列相似性,而SnRK2和SnRK3蛋白是植物所特有的一類蛋白激酶[3]。
研究表明,SnRK2蛋白在植物脫落酸(ABA)脅迫中調節參與植物滲透脅迫信號通路[4]。當ABA存在時,SnRK2蛋白的抑制因子PP2C的活性被抑制,SnRK2蛋白的功能開始作用[5]。活化后的SnRK2蛋白可以通過磷酸化ABA響應元件ABFs來調節ABA響應基因的轉錄,還可以作用于下游的一些元件,如離子通道、bZIP轉錄因子、NADPH氧化酶[6],這些蛋白與植物抗逆性關系密切。在SnRK2蛋白參與的抗逆研究中,關于非生物脅迫的報道最為常見,如將玉米的SnRK2蛋白激酶基因轉到擬南芥中,明顯增強了擬南芥的耐鹽能力;過表達蘋果的MpSnRK2.10基因明顯增強了轉基因擬南芥的抗旱性[7-8]。也有一些SnRK2蛋白相關的植物抗病報道,如胡丹丹等[9]研究表明,沉默水稻中的SnRK2類基因OsSAPK2,轉基因水稻感白葉枯病比對照顯著提高。SnRK2基因是否參與大麥對白粉菌的抗性反應還未有報道。
白粉病是我國大麥生產中的重要病害,大麥感病后一般可造成20%以上的產量損失,嚴重時損失達30%[10-11]。培育抗白粉病大麥品種不但能減少產量損失,還能減少化學農藥的使用,是控制白粉病害最有效的手段。大麥抗白粉病相關基因的克隆和抗病機理研究是抗病育種的重要基礎。‘鳳大麥7號’是一個高抗白粉病的大麥品種[12],本試驗前期對‘鳳大麥7號’進行白粉菌侵染后的轉錄組測序,發現一個SnRK2家族基因HORVU1Hr1G055340在受白粉菌侵染的‘鳳大麥7號’葉片中上調表達。本試驗擬對該基因進行克隆,并初步分析其在大麥抗白粉病中的功能,以期對‘鳳大麥7號’抗白粉病機理進行更深入的研究。
1 材料與方法
1.1 材料
‘鳳大麥7號’為大理白族自治州農業科學推廣研究院引進的大麥品種,抗白粉病;‘花30’為本實驗室育成的大麥品種,感白粉病。大麥白粉菌取于上海本地大麥試驗田。
1.2 白粉菌處理大麥幼苗
‘鳳大麥7號’和‘花30’幼苗生長至二葉一心期時,在葉片上均勻抖落新鮮白粉菌孢子,取處理后0 h、6 h、12 h、24 h的大麥葉片,用于總RNA 的提取。
1.3 大麥總 RNA 的提取和cDNA第I鏈的合成
參照李穎波等[13]方法提取大麥總RNA。所提取的RNA采用1%甲醛變性膠電泳檢測RNA 的完整性,同時測定OD230、OD260、OD280吸收值以檢測純度。cDNA第Ⅰ鏈合成具體步驟按照RevertAidTMFirst Strand cDNA Synthesis Kit(Life)說明書進行。
1.4 HvSAPK3基因序列分析、開發閱讀框(ORF)全長克隆
1.5 生物信息學分析
HvSAPK3蛋白理論相對分子質量和等電點采用Expasy的ProtParam在線分析軟件(http:web.expasy.orgprotparam)分析并預測。HvSAPK3蛋白的同源蛋白、結構域利用NCBI的BlastP在線分析軟件(https:blast.ncbi.nlm.nih.gov)查找。
多重序列比對通過Clustal X 1.83軟件進行,采用 MEGA 4.1 軟件構建系統進化樹(neighbor-joining 方法),其中bootstrap設為1 000 replicates。
1.6 熒光定量PCR(qRT-PCR)分析
qRT-PCR分析使用ABI 7500 fast 定量PCR儀進行。反應體系20 μL,包括2×SYBR Mix(Life),上、下游引物各0.4 μmolL,每個反應含大約 30 ng cDNA,3 個重復,陰性對照不加模板體系。qRT-PCR 反應程序為:94 ℃預變性30 s;94℃變性5 s,60℃退火20 s,72℃延伸20 s,45個循環。通過產物溶解曲線分析 qRT-PCR 引物擴增的特異性。利用2-ΔΔCT方法計算相對表達量。qRT-PCR引物根據HvSAPK3基因序列設計:上游引物5’-CAAGGAACTTCCGGAAAACA-3’,下游引物5’-CTCAGGCTCCTTGAAGTTGG-3’;采用大麥的Actin基因作為內參基因(Actin-F:5’-GACTCTGGTGATGGTGTCAGC-3’,Actin-R:5’-GGCTGGAAGAGGACCTCAGG-3’)。
1.7 大麥條紋花葉病毒誘導的基因沉默(BSMV Virus induced gene silencing,VIGS)
VIGS試驗過程參照Wang等[14]方法。特異性引物VIGS-F:5’-CTAGCTAGCTACGAGGCGTTGAAG-GAGTTGG-3’,VIGS-R:5’-CTAGCTAGCTTCGAAGAGTTCTCCGCCAGCA-3’,序列長度為252 bp。在接種大麥條紋花葉病毒(Barley stripe mosaic virus,BSMV)病毒10 d后觀察‘鳳大麥7號’的葉片病毒癥狀。以沉默八氫番茄紅素脫氫酶基因(PDS)植株為參照,分別對Mock(接種緩沖液)和接種BSMV:HvSAPK3植株取樣,進行qRT-PCR分析,檢測目的基因的沉默效率。取出現病毒表型的植株葉片,整齊地擺放于已滅菌的6-BA保鮮培養基上,用抖落法接種新鮮白粉菌孢子,接種6—8d后觀察葉片發病情況。
2 結果與分析
2.1 HvSAPK3基因的克隆和序列分析
根據HORVU1Hr1G055340設計跨ORF引物,進行PCR擴增并測序,發現該PCR產物含有編碼長度為1 026 bp的完整ORF核苷酸序列。該基因編碼的蛋白質相對分子質量為38.54 ku,理論等電點為6.03。將該蛋白氨基酸序列進行BLASTp同源比對,發現該蛋白在5—243位氨基酸與SnRK2蛋白激酶家族典型的結構域Cdd:cd14662相似度很高(圖1),表明該基因屬于SnRK2基因家族,故將該基因命名為HvSAPK3。

“”表示序列一致,“:”表示保守性突變,“.”表示弱半保守性突變圖1 HvSAPK3蛋白與SNRK2保守結構域Cdd:cd14662序列比對 Fig.1 Sequence alignment of HvSAPK3 protein with the conserved domain Cdd:cd14662 of SNRK2
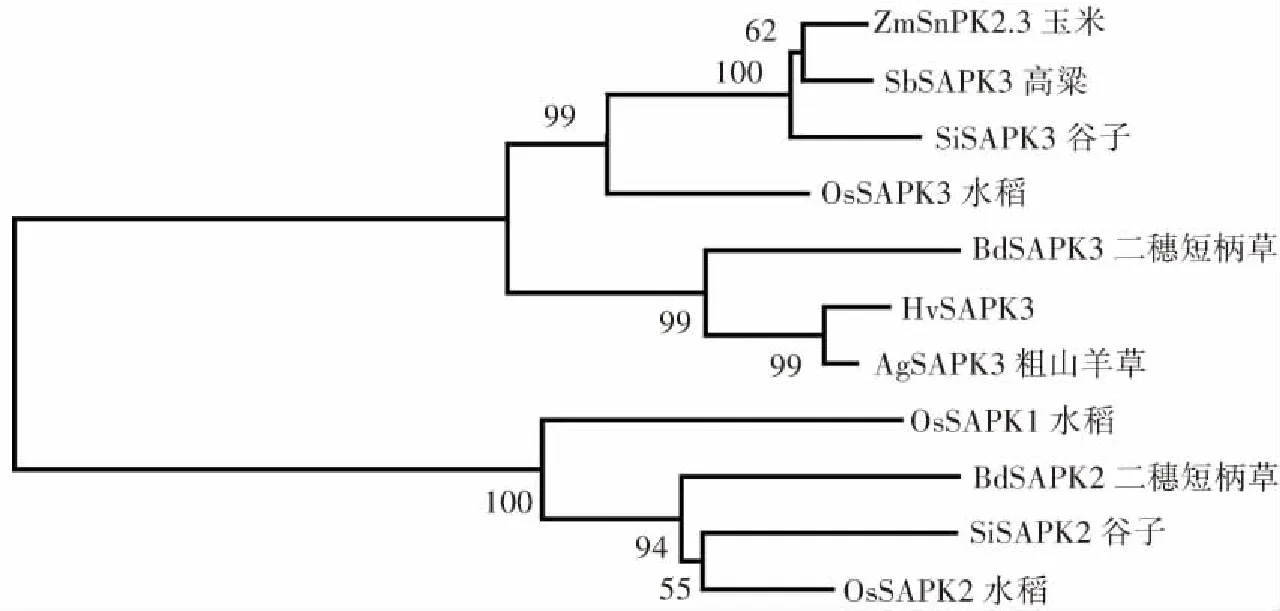
GenBank序列號:AgSAPK3(XP_020185663.1),BdSAPK3(XP_003574349.1),OsSAPK3(XP_015614267.1),ZmSnRK2.3 (NP_001136496.1),SbSAPK3(XP_021306693.1),SiSAPK3(XP_004983620.1),BdSAPK2(NP_001304800.1), SiSAPK2(XP_004958353.1),OsSAPK2(Q0D4J7.1),OsSAPK1(Q75LR7.1)圖2 部分禾本科植物SAPK同源蛋白的系統進化分析Fig.2 Phylogenetic analysis of SAPK homologous proteins in some Gramineae plants
2.2 HvSAPK3同源蛋白的系統進化分析
將HvSAPK3蛋白與粗山羊草、谷子、二穗短柄草、玉米、高粱、水稻的SnRK2 類蛋白進行系統進化分析表明:HvSAPK3蛋白和其他禾本科植物的SAPK3蛋白形成一個群(圖2),其中HvSAPK3蛋白和粗山羊草SnRK2 類蛋白AgSAPK3親緣關系最近,表明SAPK3蛋白在禾本科中進化比較保守。
2.3 白粉菌侵染后HvSAPK3基因的表達特征分析
在白粉菌侵染的‘鳳大麥7號’葉片中,HvSAPK3基因在12h時開始上調表達,隨后表達量逐漸恢復到侵染前水平(圖3A);而在感白粉病品種‘花30’ 葉片中,HvSAPK3基因在白粉菌侵染后下調表達(圖3B),表明HvSAPK3基因可能參與‘鳳大麥7號’的抗病反應。
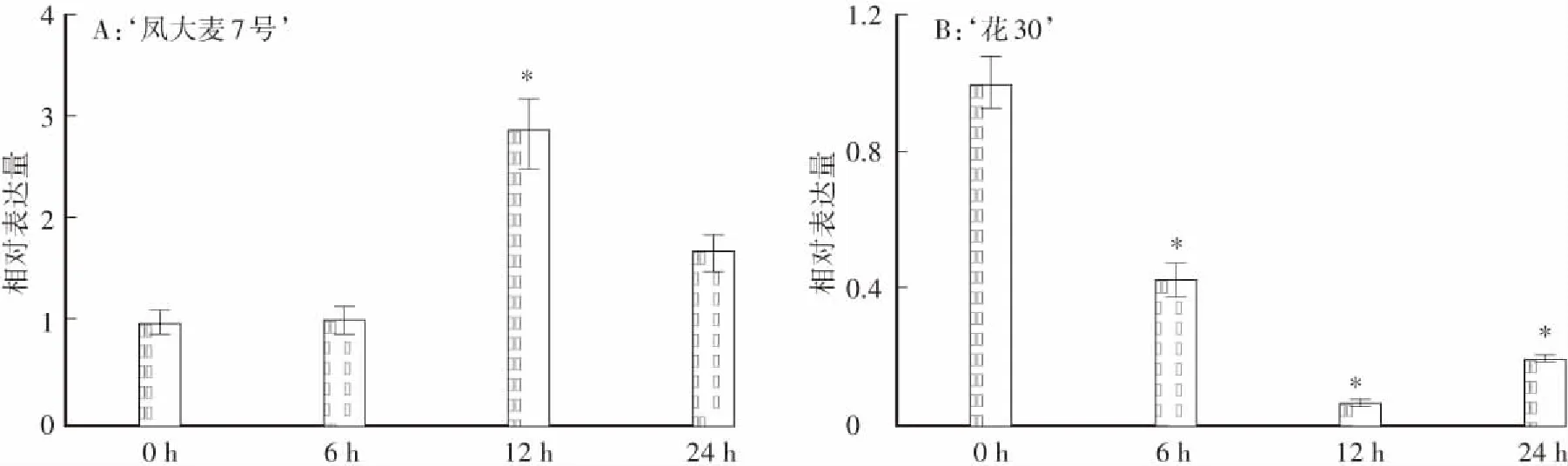
*表示P<0.05圖3 HvSAPK3基因在受白粉菌侵染的抗感大麥材料中的表達特征Fig.3 Expression characteristics of HvSAPK3 gene in resistant and susceptible barley infected by powdery mildew
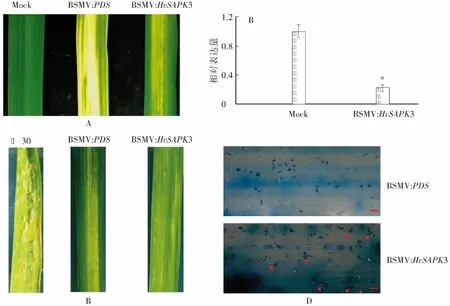
A:在‘鳳大麥7號’葉片上接種BSMV病毒載體10 d后葉片的癥狀;B:利用qRT-PCR檢測HvSAPK3基因的沉默效率,*表示P<0.05;C:沉默HvSAPK3基因植株葉片離體接種大麥白粉菌后的表型,以BSMV:PDS為對照,‘花30’為感病對照;D:接種BSMV:PDS和BSMV:HvSAPK3病毒重組載體的‘鳳大麥7號’葉片接種白粉菌后白粉菌侵染比較,紅色箭頭表示形成的分生孢子,標尺=100 μm圖4 利用VIGS分析HvSAPK3基因在‘鳳大麥7號’中的功能Fig.4 Analysis of the function of HvSAPK3 gene in ‘Fengdamai 7’ by VIGS
2.4 VIGS沉默‘鳳大麥7號’的HvSAPK3基因
將BSMV病毒α鏈、β鏈體外轉錄物分別與BSMV:PDS、BSMV:HvSAPK3體外轉錄物混合,接種二葉一心期‘鳳大麥7號’植株的第二張葉片,10d后觀察葉片上的病毒侵染癥狀。結果顯示,在參照植株(接種BSMV:PDS)的葉片上產生明顯的長條狀光漂白表型,表明植株內PDS基因已被成功沉默,病毒已成功侵染(圖4A);而在接種BSMV:HvSAPK3的葉片上,也出現明顯褪綠的病毒花斑癥狀。
病毒載體接種10 d后,利用qRT-PCR分析沉默植株中HvSAPK3基因的表達情況,同時離體接種鑒定其白粉病抗性。結果顯示,沉默植株中的HvSAPK3基因表達量和對照植株相比顯著降低,說明接種病毒葉片中HvSAPK3基因已被沉默(圖4B)。接種白粉菌孢子后,BSMV:PDS、BSMV:HvSAPK3和Mock的植株葉片上未觀察到明顯的白粉菌孢子堆生成(圖4C)。進一步通過顯微觀察發現,接種BSMV:PDS植株的葉片上白粉菌盡管能夠侵染,但無法形成分生孢子梗;而接種BSMV:HvSAPK3的植株葉片上,部分白粉菌能正常繁殖并形成次生菌絲(圖4D)。以上結果表明,在‘鳳大麥7號’中沉默HvSAPK3基因可降低其抗病性,證明該基因在抗白粉病中發揮作用。
3 討論
SnRK2是脅迫相關蛋白激酶,通過傳遞的信號誘發植物脅迫應答反應[2]。本研究以轉錄組數據為基礎,在高抗白粉病大麥品種中克隆了一個響應白粉菌侵染的SnRK2類基因HvSAPK3,進化分析顯示:HvSAPK3蛋白和其他禾本科作物中同類蛋白聚類到同一個群,表明HvSAPK3基因在禾本科作物中進化較保守,推測HvSAPK3基因在禾本科作物中的功能也是保守的。
Xu等[15]研究認為,在沒有抗病基因存在的條件下,水稻OsSAPKs家族基因不會被脅迫所誘導。在本研究中,HvSAPK3基因在‘鳳大麥7號’中受白粉菌侵染后表達上調,而在感白粉病品種‘花30’中受白粉菌侵染后表達下調,表明HvSAPK3基因可能參與‘鳳大麥7號’對白粉病的抗病反應。
胡丹丹等[9]沉默水稻中的OsSAPK2基因,轉基因水稻感白葉枯病顯著提高,顯示OsSAPK2基因的正向調控作用。在本研究中,VIGS試驗顯示,沉默HvSAPK3基因后,白粉菌能在‘鳳大麥7號’葉片上能形成次生菌絲,表明HvSAPK3基因在‘鳳大麥7號’抗病反應中起正向調控作用。目前,大麥對白粉菌專化抗性的研究較多,已報道的大麥主效抗白粉病基因位點有22個[16],HvSAPK3基因究竟參與哪個基因的調控,還需要對‘鳳大麥7號’進行更加深入的研究。
致謝:
感謝大理白族自治州農業科學推廣研究院李國強研究員提供‘鳳大麥7號’材料。
猜你喜歡
現代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
當代經濟研究(2016年5期)2016-12-01 03:12:05
現代農業(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
