臨床分離金黃色葡萄球菌ST22克隆毒力及耐藥基因分析
2019-05-09 08:50:36詹鈾超陳凱銳江雁瓊張艷玲劉正祥袁文常
實用醫學雜志 2019年7期
詹鈾超 陳凱銳 江雁瓊 張艷玲 劉正祥 袁文常,
1廣州醫科大學附屬第五醫院檢驗科(廣州510700);2廣州軍區廣州總醫院檢驗科(廣州510000);3新疆軍區總醫院檢驗科(烏魯木齊830000)
金黃色葡萄球菌(簡稱金葡萄)是引起人類感染的一種重要病原體,可引起人類皮膚和軟組織感染、肺炎、菌血癥和心內膜炎等各種疾病。金葡菌通常被分為耐甲氧西林金葡菌(methicillin-resistantS.aureus,MRSA)和甲氧西林敏感金葡菌(methicillin-susceptibleS.aureus,MSSA),MRSA 攜帶一葡萄球菌盒式染色體(staphylococcal cassette chromosomemec,SCCmec),其耐藥基因mecA可編碼PBP2a蛋白,對甲氧西林等β-內酰胺類抗生素親和力低,介導MRSA的耐藥性[1]。相對于MRSA而言,MSSA約占我國金葡菌感染的40%左右,通常認為MSSA為散在偶發株,不存在優勢克隆,具有更高的遺傳多樣性,攜帶有更多的毒力因子。但是最近的研究顯示,在我國MSSA也存在主要的流行克隆,主要有 ST121,ST398,ST5,ST7以及ST188[2]。ST22克隆是流行于歐洲的主要MRSA克隆,可以在人類、寵物(狗和貓)和醫院環境中定植傳播[3]。目前,ST22在歐洲、大洋洲及部分亞洲國家和地區廣泛流行。但是ST22克隆在中國廣泛流行未見報道,本課題通過檢測pvl等18種毒力基因、erm耐藥基因和耐藥譜分析對從烏魯木齊地區分離的105株ST22克隆進行了研究。不僅從一定程度上揭示了烏魯木齊地區ST22克隆的分子流行特征,而且可以為我國其他地區金葡菌感染和流行的檢測提供借鑒。
1 材料與方法
1.1 材料
1.1.1 菌株來源2011年6月至2016年6月期間分離于新疆軍區總醫院,經MLST分型鑒定的104株ST22型以及1株分離自病房空氣的ST22菌株。社區獲得性金葡菌(Community-acquiredS.aureus,CASA)判斷標準參照參考文獻[4]。在醫院內感染的為醫院獲得性金葡菌(Hospital-associatedS.aureus,HA-SA)。
1.1.2 儀器與試劑Bio-Rad PCR儀,Bio-Rad凝膠成像儀,生物梅里埃VITEK?2 Compact全自動細菌鑒定及藥敏分析系統及配套藥敏卡片;Tryptone Soya Broth(TSB)(CM0129B)培養基購自 OXOID公司,細菌基因組提取試劑盒(NEP021-2)購自北京鼎國生物技術公司,溶葡萄球菌素(9011-93-2)及溶菌酶(12650-88-3)購自Sigma公司。
1.2 方法
1.2.1 MRSA鑒定PCR方法檢測mecA與femB基因,反應體系為50 μL體系,反應程序設定為94℃預變性4 min,94℃變性45 s,50℃退火45 s,72℃延伸60 s,進行30次循環,最后72℃延伸2 min,反應結束后,擴增產物行1%瓊脂糖凝膠電泳,凝膠成像儀觀察結果。判斷標準以310 bp(mecA)、651 bp(femB)處出現條帶為陽性。若femB基因為陽性,mecA基因為陰性則判斷為MSSA菌株;若mecA與femB基因皆為陽性,則為MRSA菌株[1]。
1.2.2 spa分型設計引物spaF:5′-AGACGATCCTTCGGTGAGC-3′,spaR:5′-CAGCAGTAGTGCCGTTTG-3′,PCR擴增金葡菌spa基因的X區域[5]。PCR產物送北京鼎國昌盛生物技術有限公司測序,序列提交spa基因分型數據庫(http://www.ridom.de/spaserver)進行比對,根據重復序列的排列方式確定spa型別。
1.2.3 MRSA菌株的SCCmec分型應用5對特異性引物,對每株MRSA菌進行PCR擴增,擴增產物行1%瓊脂糖凝膠電泳,依據電泳圖,對菌株進行Ⅰ、Ⅱ、Ⅲ、Ⅳ和Ⅴ型的SCCmec分型[6]。
1.2.4 耐藥基因及毒力基因檢測通過PCR方法,用特異性引物擴增pvl等18種毒力基因以及紅霉素、克林霉素耐藥基因ermA、ermB、ermC,產物行1%瓊脂糖凝膠電泳驗證[7-9]。
1.2.5 藥敏試驗藥敏試驗使用VITEK?2 Compact藥敏分析系統及配套藥敏卡片GP67進行測定,抗菌藥物敏感性判斷根據美國臨床實驗室標準化協會(CLSI)2016版執行,以耐藥(R)、敏感(S)表示[10]。
2 結果
2.1 MRSA鑒定在104株ST22克隆臨床分離株中,6株細菌mecA基因陽性,定義為MRSA菌,占5.8%;MSSA菌98株,占94.2%。其中76株細菌為CA-SA,28株為HA-SA。CA-SA ST22主要引起皮膚軟組織感染(65.4%,68/104)和呼吸道感染(5.8%,6/104);HA-SA ST22主要引起術后傷口感染(13.5%,14/104)和呼吸道感染(8.7%,9/104)。見圖1。
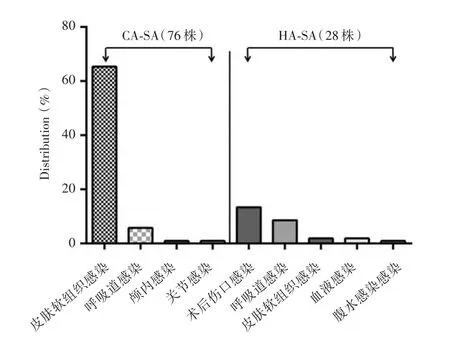
圖1 CA-SA及HA-SA ST22來源分布Fig.1 Distribution of the CA-and HA-SA of ST22 in different clinical specimens
2.2 分子分型經SCCmec分型及spa分型,6株MRSA菌株均為SCCmec IV型,t309型(圖2)。98株MSSA菌可分為12種spa型,其中86株為t309型,占87.8%(86/98),t790型2株,t12442、t12481、t13828、t15234、t1977、t284、t5335、t625、t8221,t8934型各1株。另外,從病房空氣中分離的1株MSSA-ST22為t309型。
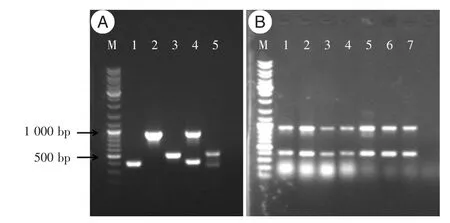
圖2 ST22-MRSA SCCmec分子分型凝膠電泳結果Fig.2 The SCCmec typing of ST22-MRSA
2.3 毒力基因檢測結果顯示104株ST22細菌中檢測到至少4種以上毒力基因,97.1%的菌株攜帶有5種以上毒力基因,73.1%的菌株攜帶有6種以上毒力基因,未檢測到see及etb基因(圖3)。sem,sen的攜帶率為100%,其次為seo(98.1%)、seg(97.1%)、pvl(79.8%)、sei(77.9%),其他毒力基因的攜帶率均低于20.0%(圖2B)。75%(78/104)的菌株攜帶有seiseg-sem-sen-seo五種腸毒素基因。對比76株CAST22與28株HA-ST22的毒力因子攜帶情況,CAST22與HA-ST22之間差異無統計學意義(表1)。
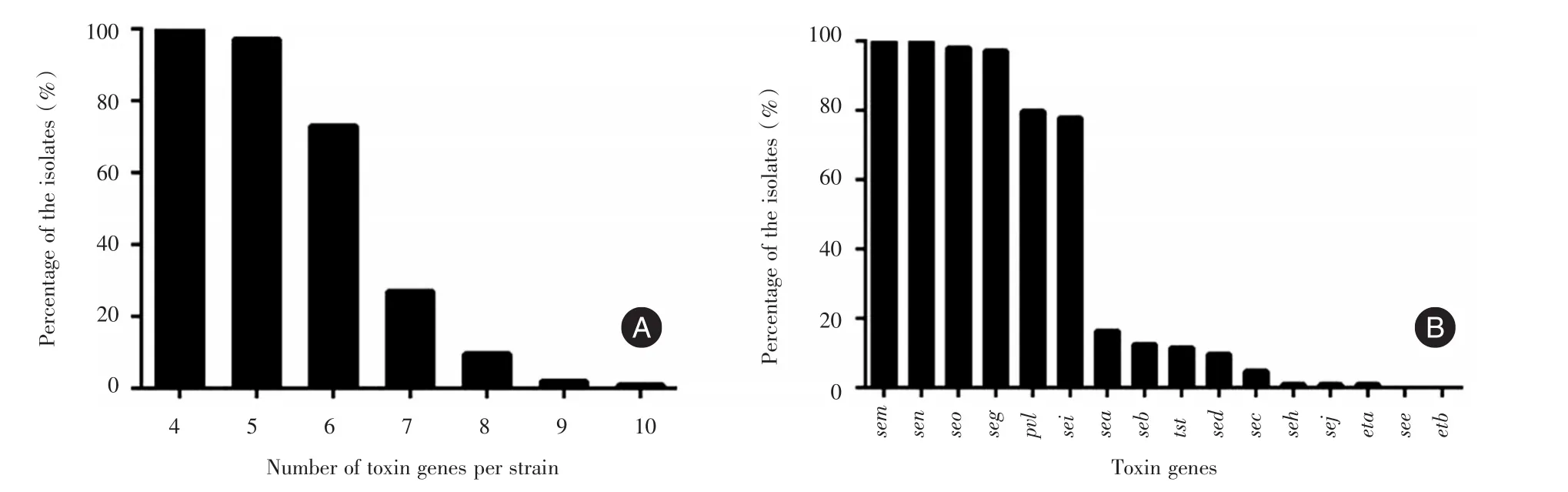
圖3 104株ST22中18種毒力基因檢測Fig.3 Detection of 18 virulence genes of 104 strains of ST22
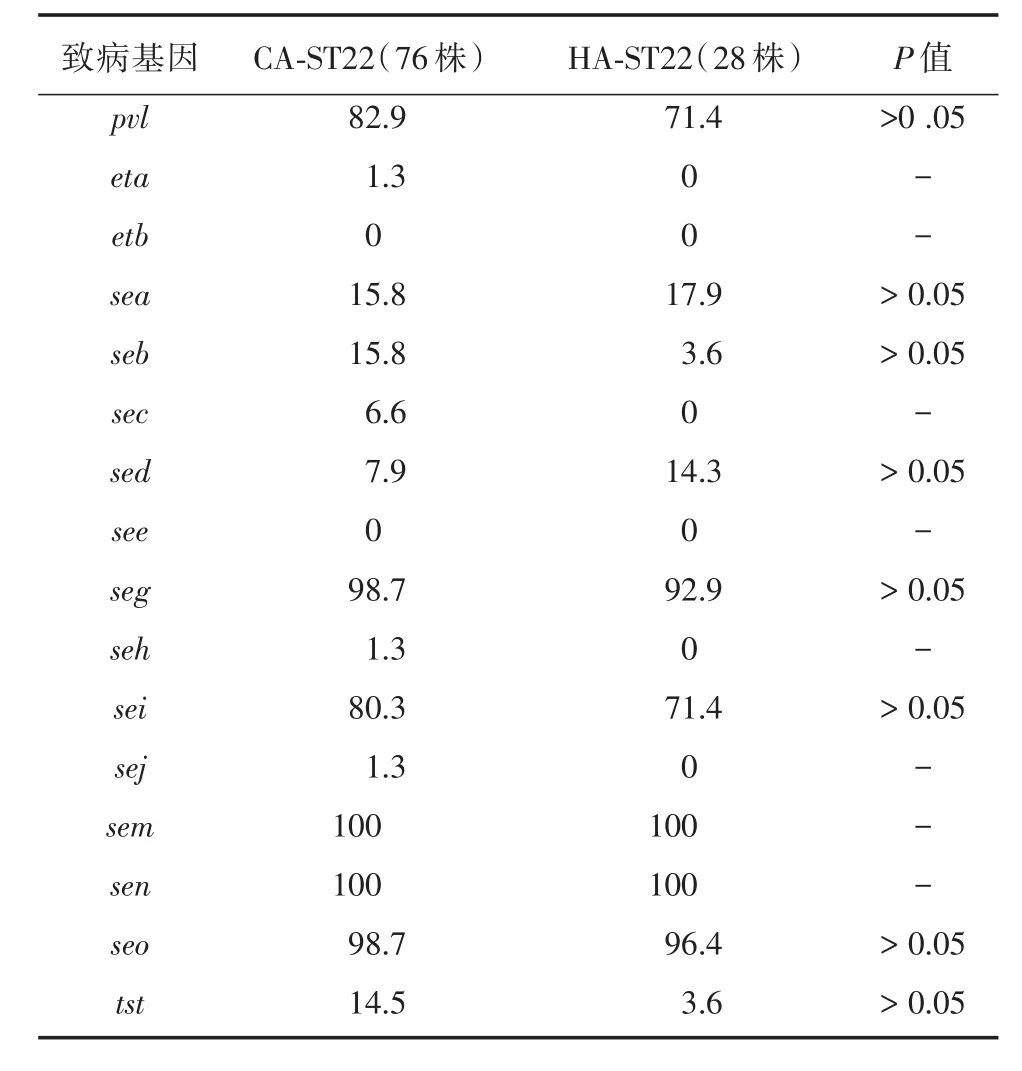
表1 金葡菌毒力基因攜帶率Tab.1 The virulence genes of S.aureus %
2.4 藥敏實驗分析及耐藥基因檢測ST22克隆對抗生素的藥敏試驗結果見表2。6株MRSA對苯唑西林、青霉素耐藥,5株對紅霉素耐藥,僅有1株細菌對克林霉素耐藥,1株對四環素耐藥,對其余被檢抗生素均敏感。MSSA菌株對青霉素耐藥,53.1%(52/98)的菌株對紅霉素耐藥,克林霉素耐藥率為7.1%(7/98),檢測到1株細菌對利福平耐藥。空氣中分離的ST22菌株僅對青霉素耐藥。
耐藥基因檢測結果顯示,在所有菌株中未檢測到ermA、ermB基因,僅在6株細菌中檢測到ermC基因,均對紅霉素耐藥,其中1株對克林霉素耐藥。
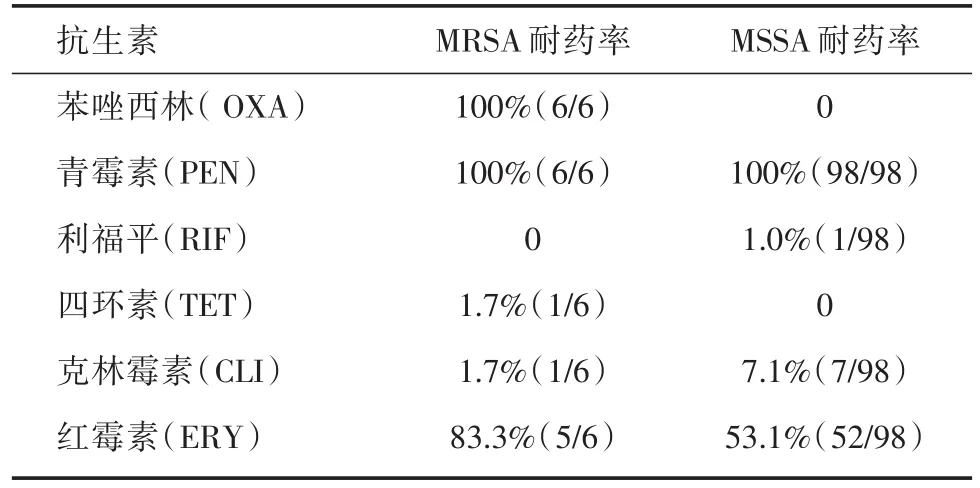
表2 金葡菌臨床分離株抗生素藥敏實驗結果Tab.2 Antimicrobial susceptibility profiles of S.aureus isolates
2.5 MRSA特征分析6株ST22-MRSA均為SCC-mecIV-t309克隆,其中4株為CA-MRSA,2株為HAMRSA,6株細菌均攜帶有sen、sei、seg、seo、sem基因。MRSA對抗生素的耐藥性不太一致,5株對紅霉素耐藥,2株對克林霉素耐藥,1株對利福平耐藥,只在1株細菌中檢測到ermC基因。
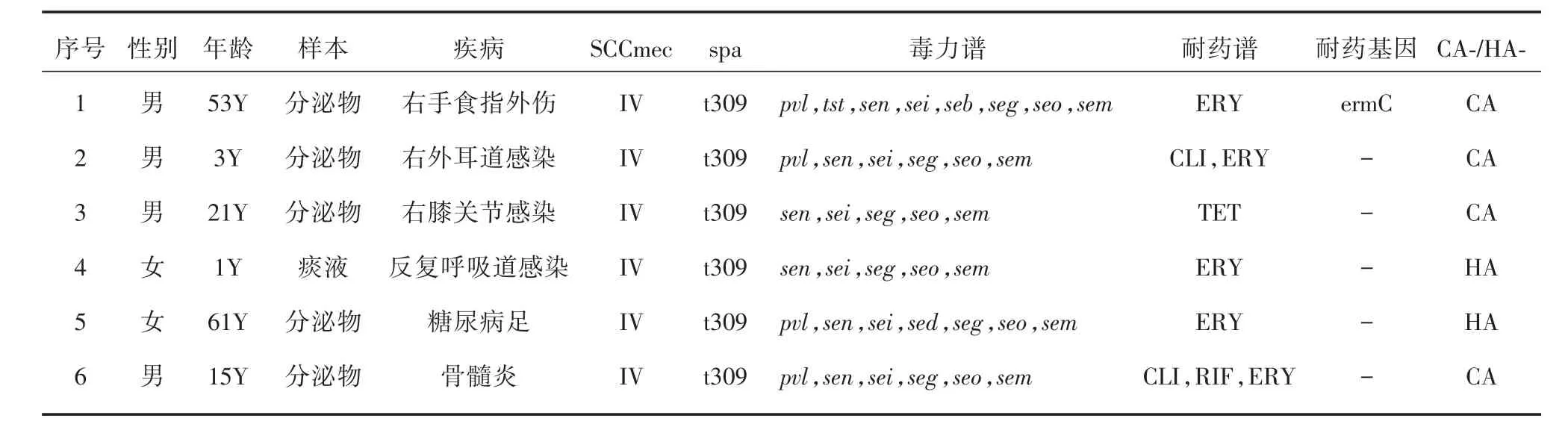
表3 ST22-MRSA臨床及分子特征Tab.3 Clinical and molecular features of ST22-MRSA
3 討論
金葡菌因其具有較強的外界環境適應能力和定植能力,在全球范圍內迅速傳播,已成為醫院和社區感染的重要病原菌。在我國,金葡菌是引起臨床感染的首要革蘭氏陽性致病菌,但是在不同地區金葡菌的主要流行克隆存在較大差異。
ST22克隆在歐洲被命名為EMRSA-15,1993年首次在英國報道,隨后廣泛的在歐洲的醫院內部、醫院之間以及不同國家快速傳播,是引起歐洲醫院感染的主要MRSA菌[3]。最近的研究顯示ST22-MRSA-SCCmecIV型克隆已經在新加坡、巴勒斯坦、印度尼西亞、印度和新加坡等亞洲國家和地區廣泛流行,并且有取代ST239-MRSA-SCCmecIII克隆的趨勢,但是在我國還很少有ST22-MRSA的報道[11-13]。LI等[14]對2011年分離自我國華山醫院的608株金葡菌的分析研究首次報道了2株ST22-MSSA克隆,其中1株為pvl陽性菌株。YANG等[2]對2013年分離自重慶的124株MSSA克隆研究發現,ST22克隆是主要流行的MSSA菌,優勢spa型別為t11413,71.4%的菌株攜帶有pvl基因。WANG等[15]研究發現,ST22-t309是引起北京地區兒童感染的主要克隆,且ST22克隆已經出現取代ST59克隆的趨勢。JIANG等[16]對55例社區獲得性皮膚軟組織感染的金葡菌研究發現,有2株為ST22-t309克隆,均為pvl陽性菌株。本研究發現ST22是引起烏魯木齊地區金葡菌感染的主要流行克隆,同時鑒定出6株ST22-MRSA-SCCmecIV-t309,與廣泛流行于歐洲及亞洲部分地區的ST22-MRSA(EMRSA-15)部分型別相一致。98株MSSA菌中,主要spa型別為t309型,另外還檢測到t790等11種spa型別,體現出遺傳的多樣性。廣泛流行于烏魯木齊地區ST22克隆與中東地區以及歐洲地區的廣泛流行ST22克隆是否具有相關性,需要進一步通過全基因組測序等方法進一步研究確認。
本研究中ST22-MRSA及ST22-MSSA,對大部分抗生素保持較高的敏感性,ST22-MRSA菌主要對紅霉素耐藥(83.3%),與流行于歐洲的EMRSA-15相似。ST22-MSSA主要對青霉素和紅霉素耐藥,對紅霉素等耐藥率達到53.1%。以往的研究顯示金葡菌對紅霉素或者克林霉素的耐藥與細菌是否攜帶有erm基因無相關性。由于erm基因存在于質粒上,容易丟失,所以部分菌株對紅霉素耐藥,但是檢測不到erm基因;同時有些菌株攜帶有erm基因,但是由于啟動子突變,菌株表現出對紅霉素敏感[7]。研究顯示MSSA菌株主要攜帶有ermC基因,但是本研究中僅在1株MRSA菌,5株MSSA菌中檢測到了ermC基因,未檢測到ermA及ermB基因,與53.1%的MSSA菌以及83.3%的MRSA菌對紅霉素耐藥表型不一致,其具體耐藥機制有待進一步研究。
金葡菌具有較強的致病性,和其攜帶有眾多的毒力因子有關。PVL與由金葡菌引起的皮膚和軟組織感染密切相關,MRSA和MSSA均有可能攜帶該蛋白,特別是社區獲得性金葡菌經常攜帶有該蛋白[17]。本研究中,79.8%的ST22克隆攜帶有pvl基因,78.8%的菌株分離子皮膚軟組織感染,進一步證實PVL蛋白與皮膚和軟組織感染密切相關。葡萄球菌腸毒素和食物中毒密切相關,是金葡菌的超抗原,能夠引起宿主細胞凋亡。本研究中ST22克隆sem、sen的攜帶率為100%,seo、seg及sei基因攜帶率也較高(均>75%),其他腸毒素基因攜帶率較低。腸毒素SEA、SEB、SEC、SED和SEE屬于經典腸毒素;SEG、SEH、SEI、SEM、SEN和SEO等屬于非經典腸毒素[9];seg、sei、sem、sen及seo這五種腸毒素基因屬于egc腸毒素基因簇,位于基因組島SaPI3上,相比于經典腸毒素,這5種腸毒素與血清的中和能力顯著降低,提示ST22-t309具有較強的致病性,有利于其傳播和流行。
綜上所述,本研究在一定程度上揭示了廣泛流行于烏魯木齊地區的ST22克隆的分子特征及耐藥情況,ST22克隆對抗生素較敏感,但是ST22-MRSA菌已出現。ST22克隆pvl基因陽性率較高,且攜帶有較多的毒力基因,具有高致病性。對ST22克隆如何在烏魯木齊地區廣泛傳播,需要結合全基因組測序等技術進行進一步研究,以防控其傳播與流行。
猜你喜歡
保健醫苑(2022年5期)2022-06-10 07:46:38
現代臨床醫學(2022年3期)2022-06-06 07:59:40
昆明醫科大學學報(2022年1期)2022-02-28 07:43:40
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
科學大眾(2020年12期)2020-08-13 03:22:22
海峽科技與產業(2016年3期)2016-05-17 04:32:12
