吐根堿對人臍靜脈內皮細胞增殖、遷移、侵襲的影響及機制
2019-05-13 06:59:40李溪朱曄吳培潔梁志偉周迎春
山東醫藥 2019年11期
李溪,朱曄,吳培潔,梁志偉,周迎春
(廣州中醫藥大學第一附屬醫院,廣州510405)
腫瘤的生長和轉移與血管的生成密切相關[1]。一方面,新生血管為腫瘤組織提供生長所需的氧氣和營養物質;另一方面,新生血管為腫瘤的轉移提供通道[2]。血管生成過程主要包括內皮細胞活化、增殖、遷移、成管[3]。因此,通過阻斷血管生成過程可能抑制腫瘤血管的形成,從而達到抑制腫瘤生長和轉移的效果。吐根堿是一種用于治療急性阿米巴病的臨床藥物,能抑制其蛋白質的形成[4]。然而,目前尚無相關報道揭示吐根堿對腫瘤血管新生的抑制作用及潛在作用機理。2018年3~11月,我們利用人臍靜脈內皮細胞(HUVECs)模擬腫瘤新生血管內皮細胞,探討吐根堿對其增殖、遷移、侵襲和成管能力的影響及其作用機制。
1 材料與方法
1.1 細胞與材料 HUVECs購于美國公司 ATCC 中心。吐根堿購于Sigam公司;DMEM培養液、胎牛血清和胰蛋白酶購于美國Hyclone公司;Transwell小室和Matrigel購于美國Corning公司;細胞裂解液購于浙江碧云天公司;PBS緩沖液、BCA蛋白定量試劑盒購于北京賽默飛世爾生物化學制品有限公司;青霉素、鏈霉、PVDF膜購于北京索萊寶科技有限公司;單克隆抗體購于Cell Signaling Technology公司;辣根過氧化物酶標記的二抗購于北京中杉金橋生物技術有限公司。
1.2 細胞培養、分組及干預方法 HUVECs用DMEM完全培養液(10%胎牛血清、100 μg/mL青霉素和鏈霉素)培養,培養溫度37 ℃,CO2濃度5%。置于培養箱內培養24 h后換液,待細胞生長至貼壁狀態,細胞融合接近90% 時傳代。取對數生長期的細胞,隨機分為陰性對照組、吐根堿低濃度組、吐根堿中濃度組、吐根堿高濃度組。陰性對照組加入細胞培養基,吐根堿低、中、高濃度組分別加入終濃度為10、20、40 nmol/L的吐根堿。每組設3個復孔。
1.3 細胞增殖能力檢測 采用MTS法。取對數生長期的HUVECs,制備成細胞密度為0.3×105/mL的懸液,每孔100 μL接種于96孔板中,同時每組設3個復孔。各組干預72 h,每孔加入20 μL的CellTiter 96 AQueous One Solution,繼續培養4 h,然后用全自動酶標儀在波長490 nm下檢測OD值,計算細胞相對存活率=(各濃度組OD值/陰性對照組OD值)×100%。
1.4 細胞遷移能力檢測 ①采用細胞劃痕實驗。取對數生長期的HUVECs,在預先用0.1%明膠包被的12孔板中每孔接種5 000個細胞。待細胞貼壁并長到90% 以上時,用滅菌的10 μL槍頭在孔中央劃十字交叉線。用PBS清洗劃掉的細胞。用不完全的DMEM饑餓細胞6 h之后棄掉培養基,陰性對照組中加不完全DMEM培養,低、中、高濃度藥物組在完全DMEM培養基中培養。置于培養箱中8 h后,用PBS洗兩次,加入2 μmol/L的calcein-AM處理細胞30 min,于倒置顯微鏡下觀察,計算細胞遷移率=(各濃度組細胞遷移數/陰性對照組細胞遷移數)×100%。②采用Transwell小室遷移實驗。將Transwell小室置于0.1%明膠包被的24孔板中,在培養箱中放置30 min以上。制備濃度為5×105/mL的HUVECs懸液,取200 μL置于EP管中,加入低、中、高濃度的藥物并放在培養箱中處理1 h。將細胞懸液接種到小室內,下層加入 600 μL完全DMEM培養基置于培養箱中 8 h, 4%多聚甲醛固定20 min,PBS洗3遍,最后用結晶紫染色30 min,倒置顯微鏡下拍照,計算細胞遷移率=(各濃度組細胞遷移數/陰性對照組細胞遷移數)×100%。
1.5 HUVECs形成管腔數量檢測 采用細胞成管實驗。每孔加80 μL的Matrigel于48孔板中,放在細胞培養箱中30 min。制備細胞密度為5×105/mL的HUVECs懸液,取300 μL置于EP管中,加入低、中、高濃度的藥物后放在培養箱中1 h。待Matrigel凝固后,將預處理的細胞懸液接到Matrigel上,放在培養箱中培養6 h。加入2 μmol/L的calcein-AM處理細胞30 min,于倒置顯微鏡下觀察并拍照,計數形成的管腔數量(成管數)。
1.6 NF-κB信號通路p65蛋白表達檢測 采用Western blotting法。用PBS洗兩遍HUVECs后,在每皿中分別加RIPA混合裂解液(含各種磷酸酶抑制劑和蛋白酶抑制劑)。4 ℃,10 000 r/min離心20 min,收集上清液,用BCA定量試劑盒測蛋白濃度,均冰上操作。蛋白加入上樣緩沖液,高溫變性后加樣,用10%分離膠和5%濃縮膠進行跑膠,轉膜、封閉 ,加入一抗,4 ℃孵育過夜,PBST洗3次,加入辣根過氧化物酶標記的羊抗兔的IgG室溫下孵育1 h,PBST洗3次,用化學發光法曝光顯影,用Image J軟件進行NF-κB細胞信號通路p65和磷酸化p65蛋白灰度值分析。
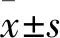
2 結果
2.1 各組細胞增殖能力比較 陰性對照組細胞存活率為100%,吐根堿低、中、高濃度組細胞存活率分別為82.22%±0.32%、70.31%±0.58%、38.54%±0.46%,吐根堿低、中、高濃度組細胞存活率均低于對照組,且吐根堿中、高濃度組均低于吐根堿低濃度組,吐根堿高濃度組低于吐根堿中濃度組(P均<0.05)。
2.2 各組細胞遷移能力比較 劃痕實驗顯示,吐根堿低、中、高濃度組的細胞遷移率均低于陰性對照組,且吐根堿高濃度組低于低、中濃度組,中濃度組低于低濃度組(P均<0.05)。Transwell小室實驗顯示,吐根堿低、中、高濃度組細胞遷移率均低于陰性對照組,且吐根堿高濃度組低于低、中濃度組,中濃度組低于低濃度組(P均<0.05)。見表1。
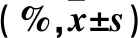
表1 各組細胞遷移率比較
注:與陰性對照組比較,*P<0.05;與吐根堿低濃度組比較,﹟P<0.05;與吐根堿中濃度組比較,△P<0.05。
2.3 各組成管數比較 陰性對照組、吐根堿低、中、高濃度組成管數分別為(38±2)、(30±2)、(20±3)、(5±1)個,吐根堿低、中、高濃度組成管數均少于陰性對照組,且吐根堿高濃度組少于中、低濃度組,中濃度組少于低濃度組(P均<0.05)。
2.4 各組NF-κB p65及磷酸化p65蛋白表達比較 吐根堿低、中、高濃度組NF-κB磷酸化p65蛋白表達均低于陰性對照組,且吐根堿高濃度組低于低、中濃度組(P均<0.05);低、中濃度組比較差異無統計學意義(P>0.05)。各組NF-κB p65蛋白表達差異均無統計學意義(P>0.05)。見表2。
3 討論
惡性腫瘤作為一類嚴重危害人類健康的疾病,是全球疾病致死的重要原因之一,全世界每年約有500萬人死于惡性腫瘤。其中,肝癌的發病率占第五位,病死率占第三位[5]。肝癌的預后普遍較差,而且多數發生在術后2年,并伴隨新的腫瘤產生[6]。肝癌發生發展較快,容易侵襲或轉移,主要是依賴于血管的生成。因此,抑制腫瘤血管生成成為治療腫瘤的重要策略之一[7]。目前FDA批準用于治療肝癌惟一的靶向藥物是抗血管新生抑制劑索拉非尼,但其藥效并不令人滿意,且不良反應大,如可引起嚴重的手足綜合征、腹瀉、皮疹等[8],而且治療費用昂貴。因此亟需要尋找安全、有效的抗腫瘤血管生成藥物用于肝癌的治療。
表2 各組NF-κB p65及磷酸化p65蛋白表達 比較
注:與陰性對照組比較,*P<0.05;與吐根堿高濃度組比較,﹟P<0.05。
新藥的研發不僅耗費大量的人力和資金,而且新藥研發周期長。諾貝爾得主詹姆斯·布萊克也曾說,“最有成效的藥物研發是從老藥開始”。例如具有解熱鎮痛作用的阿司匹林,由于其具有抗血小板聚集作用,又被人們用于預防血栓的形成[9]。因此,我們著眼于挖掘上市藥物或臨床在研藥物的抗血管生成新用途。前期研究中,我們通過HUVECs細胞增殖實驗篩選了1 200個FDA批準的藥物,同時結合內皮細胞分化和遷移功能實驗,初步發現了治療急性阿米巴病的藥物吐根堿具有潛在的抗血管新生功能。吐根堿是一種異喹啉型生物堿,存在于巴西產吐根的根中。臨床上長期被用做治療急性阿米巴病[4]。有研究發現,吐根堿可下調髓細胞白血病序列1重組蛋白(Mcl-1)蛋白從而引起腫瘤壞死因子相關的凋亡誘導配體(TRIAL)介導的凋亡[10]。除此之外,蘇宏領等[11]研究發現,吐根堿可通過抑制PI3K-Akt信號通路活性,對腎癌具有顯著的抑制作用。然而,目前尚無相關報道揭示吐根堿對內皮細胞的抑制作用及潛在作用機理。
在本研究中,我們評價了吐根堿對HUVECs多個功能的抑制效果,從而探討其可能的作用機制。本研究發現,吐根堿低、中、高濃度組細胞存活率均低于對照組,且吐根堿中、高濃度組均低于吐根堿低濃度組,吐根堿高濃度組低于吐根堿中濃度組,表明吐根堿能夠抑制HUVECs的增殖。基于以上結果,我們推測吐根堿的作用靶標可能是血管內皮細胞。
內皮細胞的遷移是血管生成中一個非常重要的過程,而腫瘤的生長和轉移離不開血管新生。因此我們分別用劃痕實驗和Transwell遷移實驗檢測吐根堿對HUVECs遷移和侵襲的影響。本研究采用劃痕實驗及Transwell小室實驗均顯示,吐根堿低、中、高濃度組的細胞遷移率均低于陰性對照組,且吐根堿高濃度組低于低、中濃度組,中濃度組低于低濃度組,表明吐根堿能抑制HUVECs的侵襲和遷移,并且在一定范圍內,吐根堿對內皮細胞的抑制濃度呈梯度依賴性,隨著藥物濃度的提高,對遷移的抑制效果不斷增強。
血管生成是一個非常復雜的過程,內皮細胞的管狀結構的形成是其中的一個關鍵步驟。本研究采用體外小管形成實驗發現,吐根堿低、中、高濃度組成管數均少于陰性對照組,且吐根堿高濃度組少于中、低濃度組,中濃度組少于低濃度組,提示吐根堿能干擾HUVEC形成管腔結構,從而阻礙腫瘤血管的生成,減少養分供給。
NF-κB信號通路是一條控制著很多生理活動的信號通路,參與細胞生長、分化、生存、凋亡、遷移、血管生成等眾多生理功能[12]。在NF-κB信號通路中,磷酸化的p65起著主要的調節作用[13]。NF-κB p65第536位絲氨酸位點磷酸化(磷酸化的p65 Ser536) 蛋白在胃癌中的表達降低,其表達與胃癌的分化程度、淋巴結轉移及TNM分期有關,并且磷酸化的p65 Ser536可能抑制胃癌細胞的遷移與侵襲能力[14]。歐志濤等[15]認為,細胞因子誘導的凋亡抑制分子1 能促進HepG2細胞增殖,與磷酸化的 p65表達量增多有關。本研究發現,吐根堿低、中、高濃度組NF-κB磷酸化p65蛋白表達均低于陰性對照組,且吐根堿高濃度組低于低、中濃度組,提示吐根堿能夠下調NF-κB磷酸化p65的表達,從而抑制HUVECs的增殖、侵襲、遷移和形成管腔結構。
綜上所述,吐根堿能抑制HUVECs的增殖、侵襲、遷移及形成管腔能力,其作用機制可能與下調NF-κB p65的磷酸化有關。
