乳酸菌和烷基多糖苷對玉米秸稈黃貯品質及其體外發酵特性影響研究
2019-05-23 06:24:36楊大盛汪水平韓雪峰湯少勛譚支良尹夢潔駱東梅
草業學報 2019年5期
楊大盛,汪水平,韓雪峰,湯少勛*,譚支良,尹夢潔,駱東梅
(1.西南大學動物科學學院,重慶 402460;2.中國科學院亞熱帶農業生態過程重點實驗室,湖南動物安全生產協同創新中心,湖南省畜禽健康養殖工程技術中心,中國科學院亞熱帶農業生態研究所,湖南 長沙 410125;3.習水富興牧業有限公司,貴州 習水 556400;4.白玉玉牧豐源畜牧科技有限公司,四川 白玉 627150)
隨著我國畜牧業的快速發展,人畜爭糧現象日益嚴重,合理利用秸稈資源,發展節糧型畜牧業,是緩解這一矛盾的重要途徑。據報道,我國農作物秸稈年產量已達8.4億t,其中玉米(Zeamays)秸稈資源最大,占秸稈可收集資源利用量的34.7%[1]。青貯和黃貯是秸稈資源保存和有效利用的主要方法。目前,對于青貯和黃貯發酵促進劑的研究主要集中在乳酸菌制劑、酶制劑和可溶性糖等3個方面。乳酸菌是黃貯過程中最重要的微生物,黃貯時加入乳酸菌菌種,可促進乳酸菌增殖,使乳酸菌快速達到所需數量,產生乳酸,降低pH值,抑制有害微生物活動,獲得較好的黃貯品質[2]。
有研究認為,微生物對有機物的分解是在一層液態膜中進行的[3]。那么,改善微生物與發酵底物之間的微環境,促進其與底物間的相互作用,可能是改善黃貯品質的有效途徑。表面活性劑可降低固/液表面張力,其疏水端可與有機物親和,使分子固定在底物表面,親水端溶于底物間隙的水中[3]。兩親基團的不同作用,使水在底物表面形成穩定的液態膜,最大可能的為微生物反應提供表面環境,此外,表面活性劑疏水端可提高可溶性有機物在水膜中的溶解度,促進微生物快速發酵[3]。目前,表面活性劑在動物生產中已進行一定的應用試驗研究,如Cong等[4]曾采用人工瘤胃產氣法研究了烷基多糖苷(alkyl polyglycoside,APG)對玉米秸稈、稻草和小麥(Triticumaestivum)秸稈體外發酵特性的影響,發現APG能夠提高粗飼料消化率。劉勇等[5]研究發現,分別在人工瘤胃液中添加0、0.02%、0.05%、0.12%(V/V)濃度的APG,其人工瘤胃液表面張力分別為54、46、43、36 mN·m-1,同時,適當降低發酵液表面張力可提高中性洗滌纖維消失率,當表面張力降低到36 mN·m-1時,會抑制底物中性洗滌纖維的消失率。但通過添加表面活性劑改變微生物與飼料作用微環境,進而改善玉米秸稈黃貯品質的研究非常少見。烷基多糖苷是一種以淀粉或葡萄糖和天然脂肪醇為基本原料,經化學合成得到的一種新型非離子表面活性劑,生產過程中無三廢排放、而且無毒、無刺激、對人體安全,對環境無害[3,5]。本試驗擬在玉米秸稈黃貯時通過聯合添加乳酸菌和烷基多糖苷,研究其對玉米秸黃貯品質的影響,并應用體外發酵技術評定黃貯玉米秸稈的體外發酵特性,最終為玉米秸黃貯提供更多有效的技術方法。
1 材料與方法
1.1 試驗材料與供體動物
乳酸菌由臺灣亞芯生物科技有限公司提供,將10 g粉狀乳酸菌(1×1012CFU·g-1)溶解在100 mL清水中,按試驗設計濃度進行噴灑;烷基多糖苷由上海國藥集團化學試劑有限公司提供;玉米秸由湖南省作物研究所提供,為秋季收獲玉米籽粒后的秸稈。
體外發酵試驗中,選用3頭成年去勢、平均體重(20.00±2.20)kg、體況良好并裝有永久性瘤胃瘺管的瀏陽黑山羊作為瘤胃液供體動物。試驗動物基礎飼糧由水稻秸稈和精料補充料組成,精粗比為40∶60,每天飼喂精料200 g,粗料300 g。山羊精飼料參照我國《肉羊飼養標準》(NY/T816-2004)配制,精料組成玉米47%、豆粕24%、麩皮22%、食鹽0.77%、石粉2.23%、預混料4%。每只瘺管羊每天08:00和16:00各等量飼喂1次,試驗期內單籠飼養,自由飲水。
1.2 試驗方法
1.2.1玉米秸稈黃貯試驗 試驗于2017年4月到2017年7月在中國科學院亞熱帶農業生態研究所進行。采用3×3雙因素試驗設計,設3個乳酸菌水平(0、1×1010、2×1010CFU·kg-1)和3個烷基多糖苷水平(0、0.10、0.20 g·kg-1),每個處理3個重復。玉米秸稈切斷至2~3 cm,加入乳酸菌與烷基多糖苷,水分調節至70%。混合均勻裝入青貯罐,壓實,避光貯藏90 d開封取樣。
1.2.2體外發酵試驗 玉米秸稈黃貯試驗結束后,采用體外瘤胃發酵的方法評定其瘤胃降解特性。參照Wang等[6]的方法,利用自主研發的全自動體外模擬瘤胃發酵系統進行體外發酵。黃貯后的玉米秸稈65 ℃烘干后,粉碎過1 mm篩。用分析天平準確稱取0.6 g發酵底物于150 mL發酵瓶中。人工瘤胃緩沖液的配制參照Menke等[7]的方法。配制好的人工瘤胃緩沖液置于恒溫磁力攪拌器上加熱使其溫度保持在39.5 ℃,并持續通入純二氧化碳2 h左右,加入少量硫化鈉以保證厭氧環境(以刃天青變為無色來判斷)。
晨飼前,通過瘤胃瘺管分別采集3只瀏陽黑山羊的新鮮瘤胃液于保溫杯中,迅速帶回實驗室,6層脫脂紗布過濾,迅速量取600 mL濾液加入到準備好的2.4 L人工瘤胃緩沖液中(瘤胃液與人工瘤胃緩沖液體積比為1∶4),用磁力攪拌器攪拌并加熱以保持瘤胃液與緩沖液混合均勻并使人工瘤胃培養液溫度保持在 39.5 ℃。利用分液器準確移取 60 mL人工瘤胃培養液依次加入到150 mL發酵瓶中,放入恒溫培養箱中進行體外模擬瘤胃厭氧發酵。恒溫培養箱設定溫度為39.5 ℃,振蕩頻率50 r·min-1,培養時間為72 h。每個黃貯玉米秸稈樣品設3個重復,每個重復2個平行。
1.3 樣品的采集與測定
1.3.1黃貯試驗樣品的采集與測定 玉米秸稈黃貯結束后,按照劉建新等[8]推薦青貯玉米感官評分標準進行評定。參照傅彤等[9]的方法對黃貯中的有機酸進行提取和凈化。取新鮮的黃貯玉米秸稈25 g于三角瓶中,加入225 mL去離子水,封口膜密封,4 ℃冰箱中浸提24 h。取2 mL浸提液,在12000 r·min-1、4 ℃條件下離心10 min后,取1 mL上清液體,-20 ℃保存用于測定揮發性脂肪酸(volatile fatty acid,VFA)、乳酸以及氨氮的測定。另取5 mL浸提液于10 mL離心管中,用于pH值的測定。剩余黃貯玉米秸稈放入信封,稱重,65 ℃烘干用于常規營養成分的分析。
VFA含量測定方法參考Wang等[10]的方法。樣品在常溫下解凍,在12000 r·min-1、4 ℃條件下離心10 min后取0.6 mL上清液裝于上機瓶中,利用氣相色譜儀(安捷倫7890A,美國)測定發酵液中的各VFA含量。采用高效液相色譜儀(安捷倫1260,美國)測定乳酸含量。采用比表面儀(Quadrasorb SI,美國康塔儀器公司)測定黃貯后玉米秸稈的比表面積(specific surface area,SSA)。比色法[11]測定氨氮含量。
按照楊勝[12]確定的常規測定方法測定黃貯后玉米秸稈干物質(dry matter,DM)、粗蛋白質(crude protein,CP)、粗脂肪(ether extract,EE)、總能(gross energy,GE)含量;依照Hall等[13]使用Fi-bretherm FT12全自動纖維儀(Gerhardt analytical systems,德國)測定中性洗滌纖維(neutral deter-gent fiber,NDF)和酸性洗滌纖維(acid detergent fiber,ADF)含量。
1.3.2體外發酵試驗樣品的采集與測定 體外發酵72 h后,通過操作系統使發酵瓶與壓力傳感器斷開連接,迅速將發酵瓶從恒溫培養箱取出終止發酵。取2 mL發酵液,在12000 r·min-1、4 ℃條件下離心10 min后,取1 mL上清液體,加入0.1 mL 25%偏磷酸固定,靜置15 min后,-20 ℃保存用于測定VFA和氨氮含量。另取3 mL發酵液,加入事先通有氮氣的5 mL離心管中,立即送往試驗室,測定Zeta電位與電子遷移率(electronic mobility,EM)。剩余樣品用于pH值、氧化還原電位和干物質消失率(dry matter disappearance,DMD)的測定。
體外瘤胃發酵VFA含量測定方法參考Wang等[10]方法。樣品在常溫下解凍,在12000 r·min-1、4 ℃條件下離心10 min后取0.6 mL上清液裝于上機瓶中,利用氣相色譜儀(安捷倫7890A,美國)測定發酵液中的各VFA含量。
用pH計(REX PHS-3C,上海儀器設備廠)迅速依次測定每個發酵瓶中發酵液的pH。發酵液經紗布抽濾,將抽濾后的發酵底物用紗布包好再將其放置鋁盒中,在105 ℃烘箱中烘8 h,稱量并記錄,計算DMD。利用Zeta電位電位儀(Brookhaven,美國布魯克海文儀器公司)測定Zeta電位與電子遷移率。利用微電極(unisense,丹麥)測定發酵液氧化還原電位。
1.4 計算公式
應用非線性軟件程序(NLREG),按照Wang等[14-15]提出的模型對體外模擬瘤胃發酵產氣曲線進行擬合。模型及相關參數計算公式如下:
式中:GPt表示t時刻的累積產氣量(mL·g-1);Vf表示潛在最大產氣量(mL·g-1);k表產氣速率(h-1);b表示形狀參數。下式同。
起始底物降解速率[FRO0,mmol·(g·h)-1],即當t=0時的底物降解速率,計算公式參考Wang等[15-16]的模型,數學表達方程為:
1.5 統計分析
用Excel初步記錄數據并做簡單處理,然后采用SAS 8.2的MIXED過程統計,對于所有分析指標,統計模型中乳酸菌與烷基多糖苷及其二者間的互作效應為固定效應,不同粗飼料間的差異采用Turkey選項進行多重比較,統計結果以平均值表示,P<0.05表示差異顯著,P<0.01表示差異極顯著。
2 結果與分析
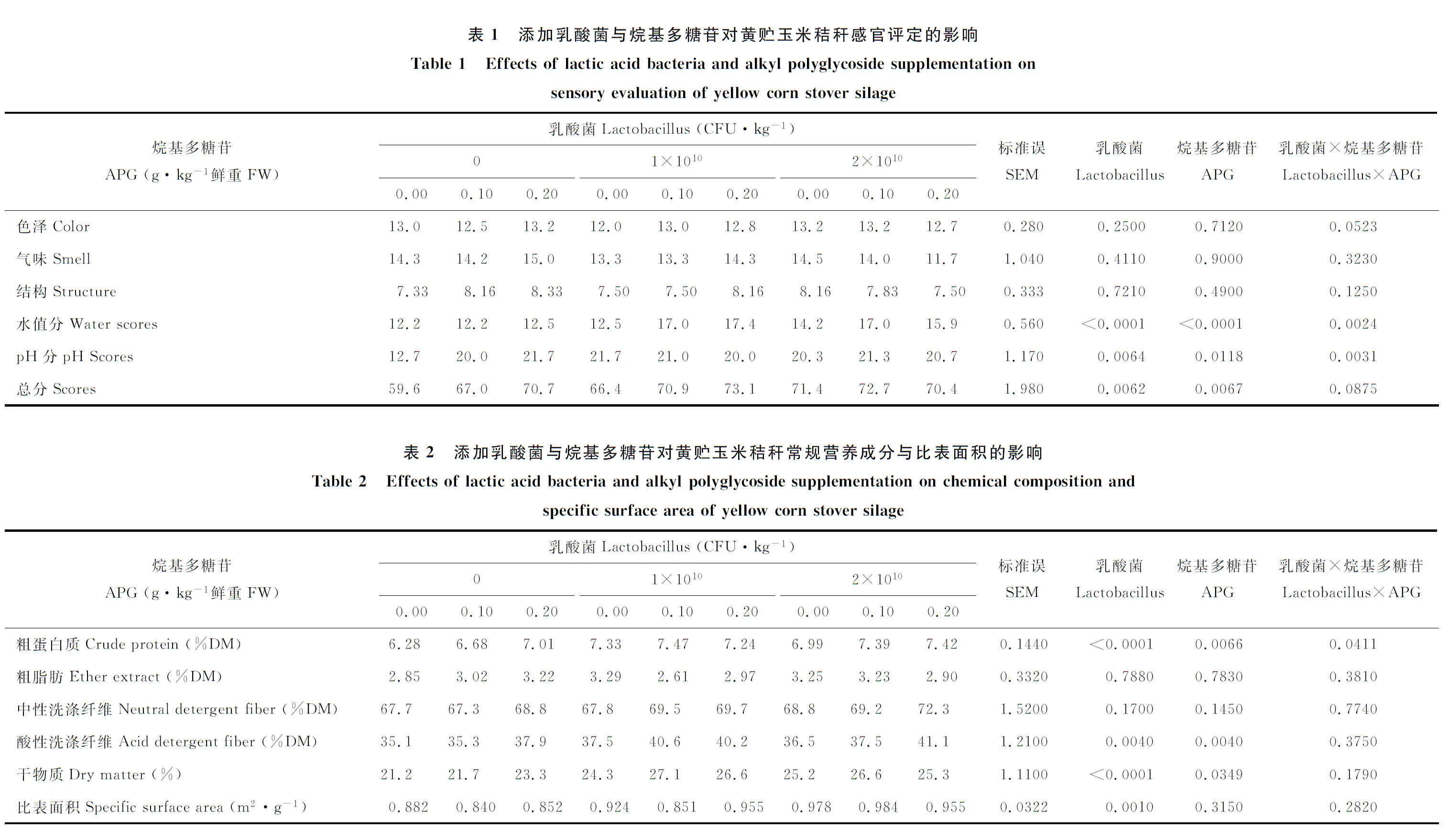
2.1 不同乳酸菌與烷基多糖苷添加水平對黃貯玉米秸稈感官評定的影響
由表1可知,乳酸菌和APG以及二者的交互作用對黃貯玉米秸稈水分含量評分存在極顯著(P<0.01)影響。當乳酸菌添加量為0、1×1010CFU·kg-1時,水分含量評分隨APG添加量的增加而升高;當乳酸菌添加量為2×1010CFU·kg-1時,水分含量評分隨APG添加量的增加先升高后降低。乳酸菌(P<0.01)和APG(P<0.05)以及二者的交互作用(P<0.01)對黃貯玉米秸稈pH分存在顯著影響。當乳酸菌添加量為0時,pH分值隨APG添加量的增加而升高;乳酸菌添加量為1×1010、2×1010CFU·kg-1時,pH分值隨APG添加量增加變化不明顯。添加乳酸菌和APG后均增加(P<0.01)了黃貯玉米秸稈的總評分值,但不同添加量之間差異不顯著(P>0.05)。
2.2 不同乳酸菌與烷基多糖苷添加水平對黃貯玉米秸稈常規營養成分與比表面積的影響
由表2可知,乳酸菌和APG以及二者的交互作用對黃貯玉米秸稈CP含量有顯著(P<0.05)或極顯著(P<0.01)影響。乳酸菌含量為0、2×1010CFU·kg-1時,黃貯玉米秸稈CP隨APG添加量增加而升高;乳酸菌含量為1×1010CFU·kg-1時,CP隨APG添加量增加先升高后降低,但是APG添加量之間差異不顯著(P>0.05)。乳酸菌和APG對黃貯玉米秸稈ADF有極顯著(P<0.01)影響,但二者交互作用影響不顯著(P>0.05)。黃貯玉米秸稈ADF隨APG添加量增加而顯著(P<0.05)升高;乳酸菌添加量為1×1010、2×1010CFU·kg-1時ADF含量顯著(P<0.05)高于乳酸菌添加量為0組,但是1×1010CFU·kg-1組和2×1010CFU·kg-1組差異不顯著(P>0.05)。黃貯玉米秸的比表面積隨乳酸菌添加量的增加而顯著(P<0.05)升高,當乳酸菌達到2×1010CFU·kg-1時比表面積達到最高。乳酸菌和APG對黃貯玉米秸稈DM有顯著(P<0.05)或極顯著(P<0.01)影響,但二者交互作用影響不顯著(P>0.05)。黃貯玉米秸稈DM隨乳酸菌和APG添加量的增加顯著(P<0.05)或極顯著(P<0.01)升高,乳酸菌添加量為1×1010、2×1010CFU·kg-1兩組間差異不顯著(P>0.05),APG添加量0.1與0.2 g·kg-1之間差異也不顯著(P>0.05)。
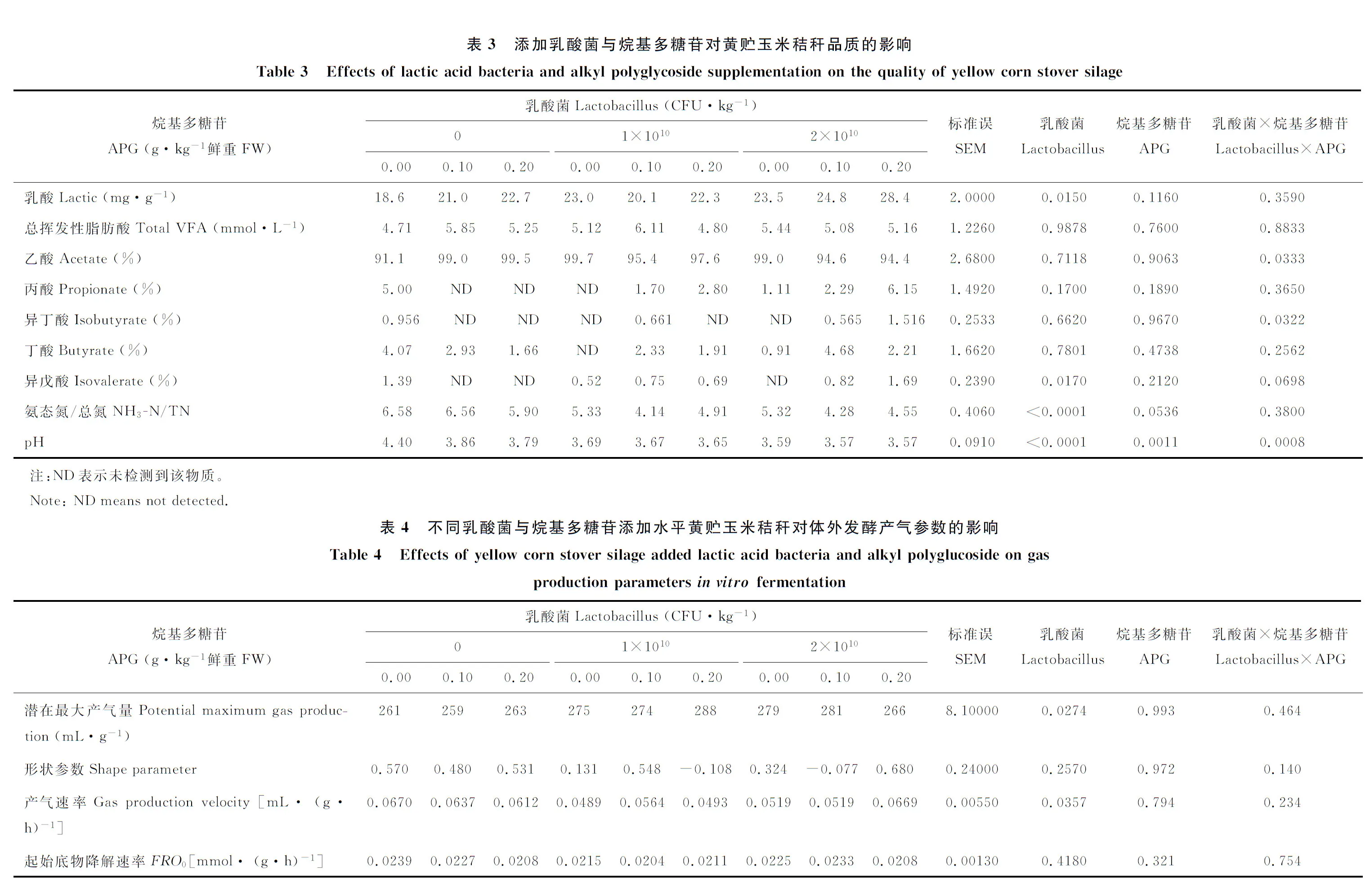
2.3 不同乳酸菌與烷基多糖苷添加水平對黃貯玉米秸稈品質的影響
由表3可知,黃貯玉米秸稈乳酸含量隨乳酸菌添加量增加而顯著(P<0.05)升高,但是APG以及二者的交互作用對其沒有顯著影響(P>0.05)。乳酸菌與APG對黃貯玉米秸稈乙酸含量具有顯著(P<0.05)的交互作用。當乳酸菌添加量為0時,乙酸含量隨APG添加量的增加而升高;當乳酸菌添加量為1×1010CFU·kg-1,乙酸含量隨APG添加量的增加先降低后升高;當乳酸菌添加量為2×1010CFU·kg-1時,乙酸含量隨APG添加量的增加而降低。黃貯玉米秸稈氨氮/總氮隨乳酸菌與APG添加量的增加而顯著(P<0.05)或極顯著(P<0.01)降低,乳酸菌添加量為1×1010、2×1010CFU·kg-1兩組間差異不顯著(P>0.05),APG添加量0.1與0.2 g·kg-1之間差異也不顯著(P>0.05)。乳酸菌與APG以及二者的交互作用對黃貯玉米秸稈pH值有極顯著(P<0.01)影響。乳酸菌添加量為0時,pH值隨APG添加量增加而降低;乳酸菌添加量為1×1010、2×1010CFU·kg-1時,pH值隨APG添加量增加變化不明顯,但是pH值均低于乳酸菌添加量為0的處理。
2.4 不同乳酸菌與烷基多糖苷添加水平黃貯玉米秸稈對體外發酵產氣參數的影響
由表4可知,添加乳酸菌與APG的黃貯玉米秸稈進行體外發酵時,乳酸菌對體外發酵潛在最大產氣量與產氣速率有顯著(P<0.05)影響。乳酸菌添加量為0時,潛在最大產氣量顯著(P<0.05)低于乳酸菌添加量為1 ×1010CFU·kg-1組與2×1010CFU·kg-1組。乳酸菌添加量為1×1010CFU·kg-1時,產氣速率顯著(P<0.05)低于乳酸菌添加量為0的處理,當乳酸菌水平提高到2×1010CFU·kg-1時,產氣速率與0、1×1010CFU·kg-1水平差異均不顯著(P>0.05)。
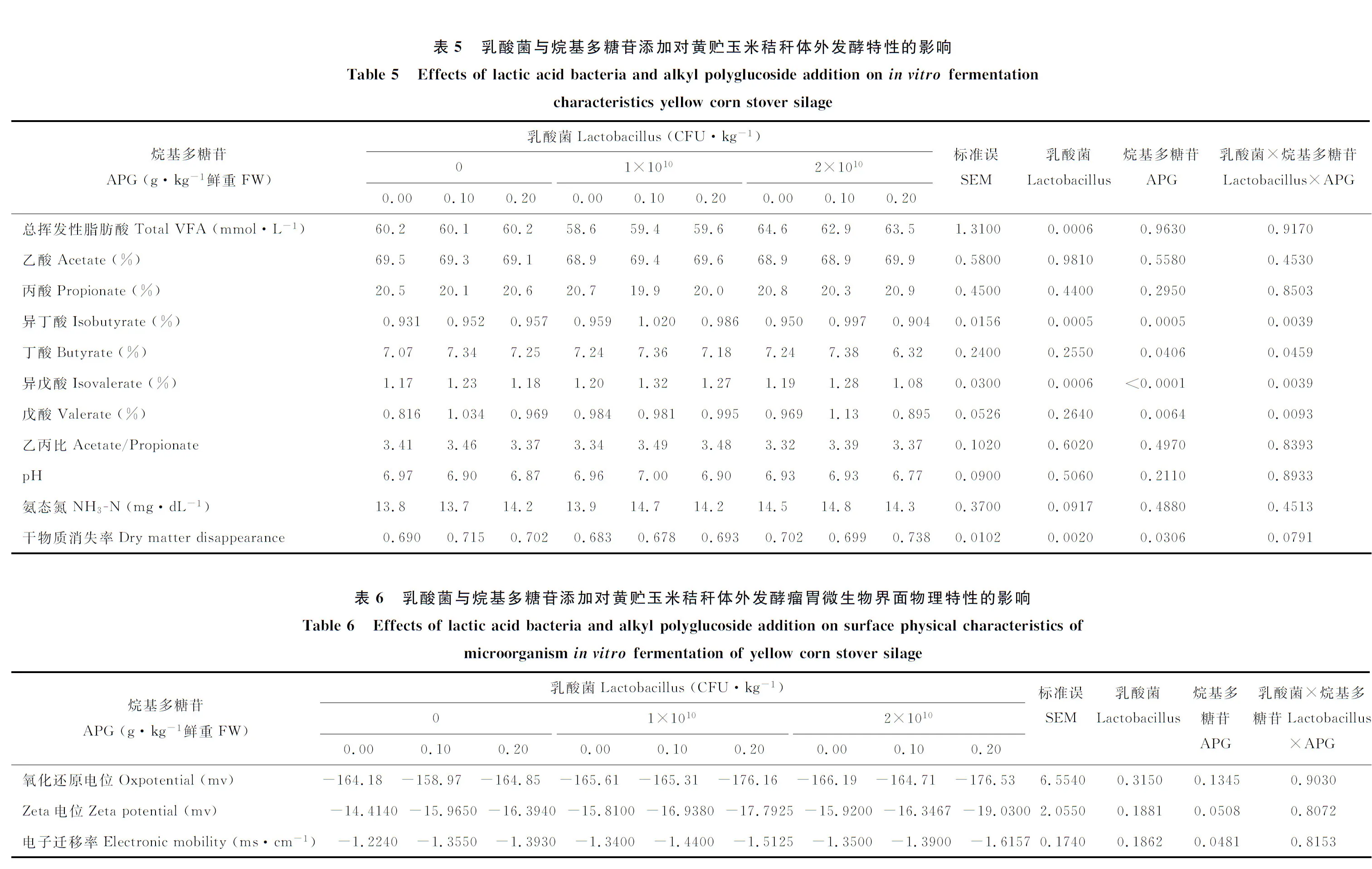
2.5 添加乳酸菌與烷基多糖苷對黃貯玉米秸稈體外發酵VFA、pH值、氨態氮及干物質消失率的影響
由表5可知,添加乳酸菌與APG的黃貯玉米秸稈進行體外發酵,乳酸菌添加量為2×1010CFU·kg-1時總VFA極顯著(P<0.01)高于乳酸菌添加量為0和1.00×1010CFU·kg-1組。乳酸菌與APG對異丁酸、丁酸、異戊酸、戊酸均存在顯著(P<0.05)或極顯著(P<0.01)的交互作用。除乳酸菌添加量為0時,異丁酸隨APG添加量的增加而升高外,其余各乳酸菌添加水平,異丁酸、丁酸、異戊酸、戊酸均隨APG添加量的增加先升高后降低。氨氮濃度隨乳酸菌添加量的增加而升高,但是差異不顯著(P>0.05)。乳酸菌與APG對DMD存在顯著(P<0.05)或極顯著(P<0.01)影響。DMD隨APG添加量的增加而顯著(P<0.05)升高;DMD隨乳酸菌添加量的增加先降低,然后升高,當乳酸菌添加量為2×1010CFU·kg-1時DMD最高。
2.6 不同乳酸菌與烷基多糖苷添加水平黃貯玉米秸稈對瘤胃電化學指標的影響
由表6可知,添加乳酸菌與APG的黃貯玉米秸稈進行體外發酵,Zeta電位與電子遷移率隨APG添加量的增加而顯著(P<0.05)提高,當APG添加量為0.2 g·kg-1時Zeta電位與電子遷移率最高。添加乳酸菌與APG的黃貯玉米秸稈進行體外發酵不影響發酵液的氧化還原電位。
3 討論
3.1 乳酸菌和烷基多糖苷對玉米秸稈黃貯品質的影響
黃貯過程主要包括好氧菌活動、乳酸菌發酵以及發酵穩定3個階段[17]。厭氧條件下乳酸菌產生乳酸,降低環境中的pH值,抑制其他腐敗微生物的活動,從而保證飼料保存及有效利用[18]。本研究中,添加乳酸菌和APG后黃貯玉米秸稈的水值分、pH分、總分均增加,總分為51~75,為良好品級。
風干后的玉米秸稈表面附著的乳酸菌較少,主要依靠外源性的乳酸菌進行發酵。鐘書等[19]認為,全株玉米青貯中添加乳酸菌,可增加乳酸的生成量,改善青貯品質。本研究發現,隨著乳酸菌添加量的增加黃貯玉米秸稈中乳酸含量顯著增加,這與Guo等[20]的研究結果一致。
本研究中,添加乳酸菌和APG對黃貯玉米秸稈pH值有極顯著的交互作用,其pH值隨乳酸菌和APG的添加而降低。APG兩親基團的不同作用最大可能的為微生物反應提供表面環境[21]。同時,其疏水端提高了可溶性碳水化合物在水膜中的溶解度[22]。故APG可為乳酸菌與玉米秸稈反應提供良好的微環境,同時,增加乳酸菌的發酵底物,加快乳酸菌繁殖和乳酸分泌,從而降低黃貯玉米秸稈pH值。
黃貯過程中氨態氮主要由微生物分解利用蛋白質和氨基酸產生,氨態氮/總氮反映了青貯飼料蛋白質降解的程度[23-24]。Li等[25]研究發現,在玉米秸稈中接種復合乳酸菌制劑后,氨態氮/總氮含量顯著降低。本研究中,黃貯玉米秸稈氨氮/總氮隨乳酸菌與APG添加量的增加而降低。表明黃貯玉米秸稈添加乳酸菌后可減少腐敗菌的繁殖,從而降低了蛋白質的分解。綜上所述,黃貯玉米秸稈中添加乳酸菌和烷基多糖苷可增加乳酸產量,降低pH值,從而抑制腐敗菌的生長和繁殖,降低蛋白質的分解,提高玉米秸稈的黃貯品質。
3.2 乳酸菌與APG對黃貯玉米秸稈營養成分與比表面積的影響
全株玉米黃貯的營養成分是評價黃貯質量好壞的一項重要指標。本研究中,添加乳酸菌和APG對黃貯玉米秸稈CP含量有顯著的交互作用,其CP含量隨乳酸菌和APG的添加而升高。Li等[25]研究發現,接種乳酸菌后,可提高玉米秸稈中的CP含量。張相倫等[2]認為,乳酸菌可通過降低發酵過程中蛋白質的降解或抑制腐敗微生物的分解,提高CP含量。對于APG對黃貯玉米秸稈CP的影響還不清楚。但是,APG通過改變乳酸菌與玉米秸稈作用的微環境,并為乳酸菌提供更多的可溶性糖,降低黃貯玉米秸稈pH值,抑制腐敗菌對CP的降解,從而達到減少蛋白質降解的效果。
黃貯玉米秸稈ADF隨APG添加量增加而升高;隨乳酸菌添加量增加先升高后降低,但降低不明顯。這與Addah等[26]的研究結果一致。乳酸菌在發酵過程中會利用玉米秸稈中豐富的可溶性碳水化合物,使細胞壁中的結構性碳水化合物比例升高[26]。APG能促進玉米秸稈中可溶性碳水化合物溶解,從而可能提高黃貯玉米秸稈中ADF比例。
黃貯玉米秸稈DM隨乳酸菌和APG添加量的增加而升高。Cai等[27]研究表明,黃貯原料表面附著的乳酸菌數較少,添加乳酸菌制劑可以保證黃貯初期發酵所需的乳酸菌數量,使之盡快進入乳酸發酵階段,使pH迅速下降,抑制腐敗微生物對營養物質的利用,減少DM損失。APG具有促進乳酸菌發酵的作用,但是未添加外源性乳酸菌的黃貯玉米秸稈表面附著的乳酸菌較少,故添加APG后DM含量略有提高,但是仍然低于乳酸菌添加組。
SSA指固體物質單位質量的表面積,包括外表面積與內表面積。本研究發現,SSA隨乳酸菌添加量的增加而升高,當乳酸菌達到2×1010CFU·kg-1時SSA達到最高。可能是乳酸菌在發酵過程中,對玉米秸稈的侵蝕加劇,使秸稈表面變得疏松多孔,增加了其比表面積。
3.3 乳酸菌與APG對黃貯玉米秸稈體外發酵特性的影響
本研究發現,乳酸菌添加量為1×1010與2×1010CFU·kg-1時,體外發酵潛在最大產氣量顯著高于添加量為0的處理。張桂杰等[28]研究發現,理論最大產氣量與CP含量呈正相關,而與NDF、ADF含量呈負相關。體外發酵潛在最大產氣量與本研究中CP的變化趨勢一致,但是與ADF的變化不一致。ADF在體外發酵過程中產氣速度較慢,而本研究對玉米秸稈體外發酵時間較長,可能使ADF的降解增加,故而降低了ADF對理論最大產氣量的影響。
乳酸菌添加量為1×1010CFU·kg-1時,產氣速率顯著低于乳酸菌添加量為0的處理,當乳酸菌水平提高到2×1010CFU·kg-1時,產氣速率與0、1×1010CFU·kg-1水平差異均不顯著。體外發酵產氣速率NDF與ADF含量呈負相關,結構性碳水化合物會降低微生物對牧草的降解速率[29]。本研究中,產氣速率的變化趨勢與添加乳酸菌和APG后玉米秸稈ADF的變化趨勢一致。
李袁飛等[30]認為,體外發酵總VFA含量與NDF/CP呈負相關關系。本試驗表明,乳酸菌添加量為2×1010CFU·kg-1時總VFA極顯著高于乳酸菌添加量為0和1×1010CFU·kg-1組,而NDF/CP隨乳酸菌添加量的增加先降低后增加。可能是乳酸菌提高了黃貯玉米秸稈的比表面積,增加了微生物與對黃貯玉米秸作用的位點,從而增加了其瘤胃降解特性,使更多的有機物在瘤胃中發酵,進而提高了總VFA含量。
瘤胃VFA中乙酸,丙酸,丁酸主要由碳水化合物發酵產生,支鏈VFA主要來源于氨基酸的降解[31]。本研究發現,除乳酸菌添加量為0時,異丁酸隨APG添加量的增加而升高外,其余各乳酸菌添加水平,異丁酸、丁酸、異戊酸、戊酸均隨APG添加量增加先升高后降低。傅海燕[3]研究發現,給山羊飼喂不同APG添加水平,不會改變瘤胃異丁酸、丁酸、異戊酸、戊酸。Cong等[4]認為,APG對VFA的影響與APG的添加劑量以及底物有關。因此,體外發酵中支鏈脂肪酸的差異可能是由于APG與乳酸菌在玉米秸稈黃貯過程中改變了其氨基酸組成所致。
氨氮是瘤胃微生物合成微生物蛋白的主要原料,其濃度高低與粗飼料蛋白質濃度以及可利用性有關。體外發酵結果表明,氨氮濃度隨乳酸菌添加量的增加而升高。發酵液中氨氮是氨基酸脫氨基后的主要產物,由于乳酸菌提高了黃貯玉米秸稈中CP含量,為微生物提供了更多可利用的發酵底物,故提高了氨氮濃度。
李袁飛等[30]認為,體外發酵DMD與NDF/CP呈負相關關系。本研究發現,DMD隨APG添加量的增加而升高,而NDF/CP隨APG添加量的增加先降低后增加。與傅海燕[3]的研究結果一致。飼料在瘤胃中的降解主要靠瘤胃微生物酶的作用[32-33],非離子表面活性劑因其界面活性作用而提高了微生物和微生物酶對底物的吸附性[34],增加了微生物作用的表面積,使黃貯玉米秸稈在瘤胃中的降解率增加[35]。DMD隨乳酸菌添加量的增加先降低而后升高,而NDF/CP隨乳酸菌添加量的增加先降低后增加。乳酸菌添加量為1×1010CFU·kg-1時,DMD下降可能與該添加量下ADF含量較高有關。當乳酸菌添加量為2×1010CFU·kg-1時,DMD升高,可能是由于乳酸菌提高了該組的比表面積,增加了其降解特性,這與總VFA的變化規律一致。
微生物表面含有磷酰基、羧基和氨基等官能團,表面活性劑與這些基團結合后會發生不同程度的電離[36]。微生物表面的Zeta電位對維持細胞膜以及反應體系的穩定性具有重要作用[37]。對反應體系而言,Zeta電位絕對值越高,體系越穩定[38]。Hong等[39]認為,微生物表面的Zeta電位對微生物對底物的吸附過程起到一定的作用。本研究發現,隨APG添加量的增加微生物表面的Zeta電位和電泳遷移率均提高。表明APG可以提高瘤胃反應體系的穩定性并且對瘤胃微生物對底物的黏附有一定的作用。
4 結論
添加乳酸菌和烷基多糖苷可增加黃貯玉米秸稈中乳酸產量,降低pH值,降低蛋白質的分解,提高玉米秸稈的黃貯品質與營養價值。
添加乳酸菌和烷基多糖苷可改善黃貯玉米秸稈其體外發酵特性,提高干物質消失率。
乳酸菌添加量為2×1010CFU·kg-1,烷基多糖苷添加量為0.2 g·kg-1時,黃貯玉米秸稈既能有較好的黃貯品質,又能提高其瘤胃降解特性。
