亞熱帶北緣次生闊葉林土壤性狀和菌根真菌多樣性隨坡位的變化
2019-05-31 08:55:08鄂曉偉田野李曉鳳馬旭徐葉寧
生態環境學報 2019年4期
鄂曉偉,田野,李曉鳳,馬旭,徐葉寧
南京林業大學林學院/南京林業大學南方現代林業協同創新中心,江蘇 南京 210037
菌根是土壤真菌與植物根系在長期演化過程中形成的具有特定結構和功能的互利共生體(劉潤進等,2007),是自然生態系統中廣泛存在的共生現象。據估計,陸地生態系統中90%以上的植物根系都能形成菌根(Allen,1992)。菌根根據其共生方式可分為外生菌根、內生菌根和內外生菌根,而森林生態系統中通常以外生菌根類型占主導地位(Buee et al.,2005;Izzo et al.,2005)。森林生態系統中的許多木本植物的生長和生存都依賴于菌根。菌根釋放的菌絲體作為寄主植物的主要營養器官的延展,能顯著改善寄主植物對于水分和氮、磷等營養元素的吸收(Grelet et al.,2009;Leake et al.,2004;Singh et al.,2007)。菌根真菌的存在也可以影響根際土壤的理化性質,改變植物根際的微生物環境,最終影響寄主植物的生長(吳克,2003)。因此,菌根真菌的侵染和共生是林木適應瘠薄土壤條件和困難立地的有效方式。
菌根真菌在大多數生態系統中具有高度多樣性,其組成和多樣性的不同對寄主植物生長的影響也可能表現出“多樣性效應”(Hoeksema et al.,2010;Koide,2010)。雖然影響菌根真菌多樣性的機制仍然不清楚,但寄主植物特性以及土壤條件的差異可能對其具有關鍵性作用(Bruns,1995)。不同類型的菌根真菌利用土壤養分的能力不同,對非生物因子的適應能力也有所不同,這些因素都可能影響真菌與林木的共生過程,導致菌根的結構和功能表現出不同的差異。土壤養分及有機質含量會影響土壤真菌的侵染率,如N、P含量過高時會降低菌根真菌侵染率(安秀娟等,2007;孫玥等,2007;王曉英等,2009;鐘思遠等,2016)。
在山地生態系統中,坡位的差異會導致土壤的溫、濕度以及養分等條件產生差異,從而可能影響菌根真菌的侵染及其群落結構特征。因此,把握菌根真菌的群落和種群結構特征對理解這些共生真菌種群在山地不同坡位森林生態系統中所起的作用至關重要(Egger,1994;Dahlberg et al.,1995;Dahlberg,2001)。
目前有關菌根真菌方面的研究多側重于寒帶、溫帶和熱帶,對亞熱帶的研究則主要集中在人工林方面,對地帶性的闊葉林研究較少(Smith et al.,2008)。隨著我國天然林保護和退耕還林等林業重點工程的實施,亞熱帶地區地帶性次生闊葉林面積迅速增加。這些次生闊葉林多分布于土壤條件貧瘠、惡劣的山地,在其形成、恢復和演替過程中,與菌根真菌的共生可能是其適應該種立地的關鍵,因此研究菌根真菌的特性隨自然立地條件的變化,在將來不斷擴大的次生林管理方面有很大的參考價值。本研究以亞熱帶北緣丘陵山區處于不同坡位的天然恢復的次生闊葉林為研究對象,探討由于坡位不同所引起的自然立地條件變化對林木菌根真菌侵染及多樣性的影響,并分析菌根真菌侵染特征與土壤養分條件之間的關系,以期為次生闊葉林的經營管理提供參考。
1 材料與方法
1.1 研究地概況
研究區位于江蘇省句容市南京林業大學下蜀林場的空青山(N31°59′,E119°14′),屬寧鎮山脈東段的低山丘陵區,最高海拔 322.6 m。該地區屬于亞熱帶季風氣候區北緣,年均氣溫 15.2 ℃,年均降水量1005.6 mm。該區的地帶性植被為帶有常綠成分的落葉闊葉林,以殼斗科櫟屬落葉樹種為主,是北亞熱帶向暖溫帶過渡的植被類型。該地區的土壤為在石英砂巖和砂頁巖上發育而成的黃棕壤。
本研究以空青山東南坡面 1950年代初封山育林后恢復形成的次生落葉闊葉林為研究對象,林分群落保存完好。林分上層建群喬木以麻櫟(Quercus acutissima)和栓皮櫟(Q.variabilis)為主,含少量楓香(Liquidambar formosana)。小喬木和灌木層種類較多,主要包括短柄枹(Q.glandulifera)、白檀(Symplocos paniculata)、刺楸(Kalopanax septemlobus)、葉下珠(Phyllanthus urinaria)、杜鵑(Rhododendron simsii)、構骨(Ilex cornuta)、絡石(Trachelospermum jasminoides)、菝葜(Smilax china)等。草本層較少,以麥冬(Ophiopogon japonicus)和少量蕨類植物為主。雖然山體海拔較低,但由于整體上為石質山地,坡度較大,巖石出露較多,土層淺薄,石礫含量高,立地條件差,主林層的高度從下坡到上坡也下降明顯。本研究在次生闊葉林內根據主林層的大致生長情況,從上坡到下坡選擇3個調查樣區,用于調查土壤養分條件和菌根真菌侵染特征。調查樣區的基本情況見表1。
1.2 樣品采集及處理
于2016年7月在上、中、下3個坡位各隨機選擇3個10 m×10 m的重復取樣小區,在每個小區內設置3個20 cm×20 cm的小樣方,剪除地表全部林下植被,去除地表枯枝落葉層后,將樣方內0-20 cm土層的土壤與根系全部取出,用自封袋密封后,置于4 ℃冰盒內低溫保存并帶回實驗室,盡快將根系與土壤分離。新鮮土壤樣品過2 mm篩去除石礫,測定含水量后,取一部分保存在4 ℃冰箱內,用于土壤無機氮含量和微生物生物量測定;另一部分樣品風干,用于土壤pH值和有效磷含量以及土壤有機碳和全氮、全磷含量測定,其中土壤有機碳、全氮和全磷含量測定樣品需進一步過0.1 mm篩。分離出的植物根系在靜水中將附著的土壤顆粒輕輕抖落,然后用純水潤洗干凈后,用吸水紙吸干水分,一部分保存于4 ℃冰箱,用于菌根真菌的侵染狀況及菌根形態觀察;另一部分保存于-80 ℃冰箱內,用于根系共生的菌根真菌DNA提取和后續的多樣性分析。
1.3 分析測定方法
1.3.1 土壤含水量及養分含量測定
土壤含水量采用烘干法測定。土壤pH值采用電極法測定,土水比為 1∶2.5(w/v)。土壤全氮、無機氮、全磷及有效磷含量的具體測定參照美國土壤學會的土壤分析測定方法(Sparks et al.,1996)。土壤全氮含量采用凱氏消煮-靛酚藍比色法測定。土壤全磷含量采用濃硫酸-高氯酸消煮-鉬銻抗比色法測定。使用 2 mol·L-1KCl溶液浸提(w/v=1∶5,180 r,1 h)土壤,采用靛酚藍比色法測定NH4+-N含量,雙波段紫外分光光度法測定NO3--N含量。土壤有效磷含量采用 0.5 mol·L-1NaHCO3(pH=8.5)浸提(w/v=1∶20,180 r,30 min)-鉬銻抗比色法測定。土壤有機碳含量采用重鉻酸鉀氧化-外加熱法測定。土壤微生物生物量采用氯仿熏蒸抽提法進行提取(Tian et al.,2008),微生物生物量碳(MBC)含量采用重鉻酸鉀氧化-外加熱法測定,微生物生物量氮(MBN)含量采用茚三酮顯色法測定。

表1 調查樣區基本情況表Table 1 Basic information of the investigation forests in different slope positions
1.3.2 外生菌根侵染率測定及形態觀測
本研究僅針對外生菌根進行了侵染率和形態觀測。參照劉潤進等(2007)的描述從形態方面對根尖菌根真菌侵染和形態進行了觀察,凡是能夠觀察到根尖明顯膨大,有菌絲套覆蓋,或有外延菌絲,則表明根尖已經被侵染。將洗凈的細根樣品置于70%的乙醇中浸泡固定,測定時從中隨機取一定量的完整活根在40倍體視顯微鏡(MOTIC SMZ-140 SERIES)下進行鏡檢和拍照,分別判斷和記錄每個視野的根尖總數量及被菌根侵染的根尖數量。整個觀察過程,上坡采集了593個視野共1928個根尖,中坡采集了432個視野共1616個根尖,下坡采集了227個視野共1550個根尖。
1.3.3 菌根真菌多樣性分析
菌根真菌的多樣性采用 PCR-變性梯度凝膠電泳(PCR-DGGE)法進行分析。將于-80 ℃保存的根系樣品采用液氮進行研磨后,利用試劑盒(MOBIO PowerSoil DNA isolation kit)提取與細根共生的菌根真菌總DNA,提取方法按照試劑盒使用說明進行操作。
選用真菌18S rDNA基因特異性引物對GCFung(5′-CGCCCGCCGCGCCCCGCGCCCGGCCCGCCG CCCCCGCCCCATTCCCCGTTACCCGTTG-3′) 和NS1(5′-GTAGTCATATGCTTGTCTC-3′)對提取的菌根真菌總 DNA進行 PCR擴增(SCILOGEX TC1000-G,America)。50 μL PCR擴增反應體系為:2 μL Template DNA;1 μL Primer F;1 μL Primer R;25 μL Premix Taq 酶;21 μL ddH2O。擴增反應條件為:94 ℃ 30 s,47 ℃ 45 s,72 ℃ 1 min,35個循環。
采用Bio-Rad公司的DcodeTM基因突變檢測系統對PCR擴增產物進行DGGE分析。聚丙烯酰胺凝膠濃度為 6%,變性劑濃度梯度范圍為 30%-60%。電泳運行條件:1×TAE電泳緩沖液,75 V電壓,60 ℃電泳溫度,電泳時間16 h。
電泳結束后對凝膠進行銀染,采用Bio-Rad全自動紫外凝膠成像系統進行觀測并拍照,獲得菌根真菌的DGGE條帶圖譜。使用Quantity One軟件(Version 4.5,Bio-Rad,USA)對條帶圖譜去噪后,確定每個樣品的條帶數量和每個條帶的相對亮度峰值,并計算菌根真菌的群落多樣性指數。
1.4 數據統計和分析
菌根侵染率=侵染根尖數/鏡檢總根尖數×100%。
根據Quantity one軟件的測定結果計算菌根真菌基于基因型的群落多樣性指數,包括Shannon-Weiner多樣性指數(Shannon-Weiner diversity index,H′)、Simpson 指數(Simpson diversity index,D)、豐富度指數(Richness index,S)、Pielou均勻度指數(Pielou evenness index,J)。各指數的計算方式如下:

式中,Pi為第i個DGGE條帶出現概率;Ni為第i個DGGE條帶的擴增量;N為土壤微生物群落中DNA的DGGE條帶擴增總量(N=∑Ni);S為樣品全部真菌種類數(條帶數)。
運用Microsoft Excel 2013對各樣地的土壤養分指標以及菌根侵染率進行計算、處理和繪圖;SPSS 19.0對各指標進行單因素方差分析(One-way ANOVA)和多重比較(Duncan);quantity one 軟件分析凝膠電泳圖譜。
2 結果與分析
2.1 不同坡位土壤特性比較
2.1.1 土壤含水量、pH值及養分含量
不同坡位林地土壤含水量、pH值及養分含量見表 2。不同坡位林分土壤的含水量范圍在 25.1%-29.8%之間,隨坡位下降略有升高,但差異不顯著(P=0.39)。不同坡位林地土壤均呈明顯的酸性,pH值范圍在 3.8-4.1之間,隨著坡位下降略有上升,也不存在顯著差異(P=0.14)。不同坡位林地土壤有機碳含量為 23.6-35.2 g·kg-1,隨著坡位下降含量升高,其中上坡的有機碳含量顯著低于中、下坡(P=0.03)。不同坡位林地土壤全氮和全磷含量變化規律相似,均為上坡<下坡<中坡,上坡位土

表2 不同坡位林地土壤理化性質Table 2 Physical and chemical properties of forest soils in different slope positions
壤全氮、全磷含量顯著低于中坡位,而下坡位與上、中坡位沒有顯著差異(P<0.05)。土壤無機氮含量以中坡最高,達 38.6 mg·kg-1,顯著高于上坡和下坡(P=0.01)。各坡位土壤無機氮以硝態氮為主,占 56.7%-75.4%。土壤硝態氮含量以上坡最低(14.3 mg·kg-1),顯著低于中坡和下坡(P=0.01);上坡和中坡的土壤銨態氮含量略高于下坡,但差異不顯著(P=0.10)。土壤有效磷含量范圍在 8.9-10 mg·kg-1之間,上坡和下坡略高于中坡,差異不顯著(P=0.39)。
2.1.2 土壤微生物生物量碳、氮含量
不同坡位林地土壤微生物生物量碳、氮含量變化見圖 1。不同坡位林地的土壤微生物生物量碳含量存在差異,以下坡最高,達718 mg·kg-1,顯著高于上坡和中坡(P=0.05)。不同坡位土壤微生物生物量氮含量表現為上坡<中坡<下坡,在不同坡位間差異不顯著,含量在 192.7-229.4 mg·kg-1之間(P=0.17)。上、中、下坡位林地土壤微生物生物量C/N比范圍為 2.5-3.1,中坡顯著低于上、下坡(P=0.04)。
2.2 菌根真菌的侵染率和形態特征
上、中、下坡林地林木細根的菌根真菌侵染率見圖 2。不同坡位間菌根真菌的侵染率差異顯著(P=0.04),呈現出從上坡向下坡顯著下降的趨勢。上坡的菌根真菌侵染率達到92.1%,中坡的菌根真菌侵染率有所下降,為84.7%。而下坡的侵染率最低,僅為74.6%。
不同坡位林分菌根的形態略有差異,但并不明顯。上坡林分中的菌根形態多表現為黃色至暗紅色(圖3A),膨大,光滑,無外延菌絲;部分表現為棕色至黑色,具有外延菌絲(圖3B);還有部分為白色并具疏松菌鞘(圖3C)。中坡絕大多數菌根表現為黃色至暗紅色,膨大,光滑,無外延菌絲(圖3D-F)。下坡與中坡相似(圖 3G-I),菌根侵染較少,且形態變化不大,多為簇狀。
2.3 菌根真菌的群落多樣性
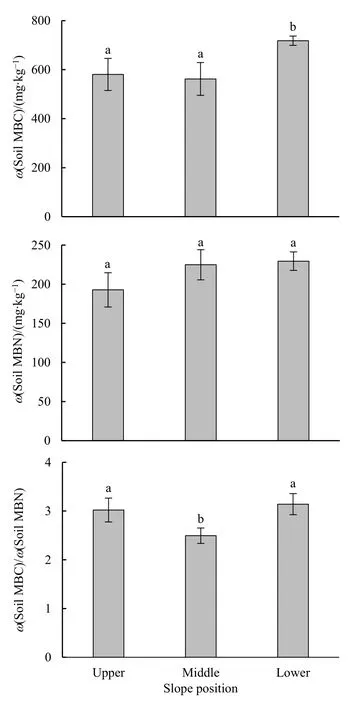
圖1 不同坡位林地土壤的微生物生物量碳、氮含量及碳氮比Fig. 1 Microbial biomass carbon and nitrogen content and C/N ratio of forest soils in different slope positions
不同坡位林木菌根真菌的 DGGE電泳圖譜及泳道識別圖如圖4所示,泳道識別圖以7號泳道為標準進行分析,泳道條帶按照與 7號泳道條帶相似度大小排列,百分數越大,條帶之間相似性越高,說明它們之間的真菌群落結構相似程度較高。泳道中的條帶粗細不一,條帶比較粗黑則表示真菌群落密度較大。各個坡位可檢測的條帶數量為10-20條,其中上坡的條帶數量明顯高于中、下坡。圖中的特異性條帶表明不同坡位土壤間菌根真菌群落結構有差異,其中以上坡和中坡的特異性條帶較多。

圖2 不同坡位林地土壤的菌根真菌侵染率Fig. 2 Mycorrhizal fungal infection ratio of forest soils in different slope positions
基于 DGGE電泳圖譜所計算出來的不同坡位林分細根菌根菌群落的多樣性指數見表 3。不同坡位菌根真菌群落的 Shannon-Wiener指數(H')、Simpson指數(D)和豐富度指數(E)差異顯著,表現為上坡>中坡>下坡(P<0.05)。而各坡位土壤菌根真菌群落的均勻度指數(J)沒有顯著差異,整體上隨著坡位降低而略有下降。總體而言,上坡的菌根真菌種群具有更加豐富的多樣性。
2.4 菌根真菌侵染率、多樣性和土壤理化性狀的關系
對菌根真菌侵染率及多樣性指數與土壤性狀進行相關性分析,結果(表 4)表明,不同坡位林地菌根真菌侵染率與土壤有機碳、硝態氮含量呈顯著負相關,但與其他土壤養分因子相關性不明顯。菌根真菌Shannon-Wiener指數與土壤pH、有機碳含量和硝態氮含量均呈顯著負相關,與土壤銨態氮含量呈顯著正相關;Simpson指數與土壤有機碳含量呈極顯著負相關;豐富度指數與土壤含水量、有機碳含量、有效磷含量和微生物生物量氮含量呈顯著負相關。

圖3 上、中、下坡林分細根的代表性菌根形態Fig. 3 Representative mycorrhizal morphology of fine roots in upper, middle and lower slopes

圖4 不同坡位林分菌根真菌DGGE電泳圖譜及泳道識別圖Fig. 4 DGGE bands and lane maps of mycorrhizal fungi in different slopes

表3 不同坡位林地土壤菌根菌群落DGGE群落多樣性指數Table 3 Diversity indexes of soil mycorrhizal community in different slopes based on DGGE analysis
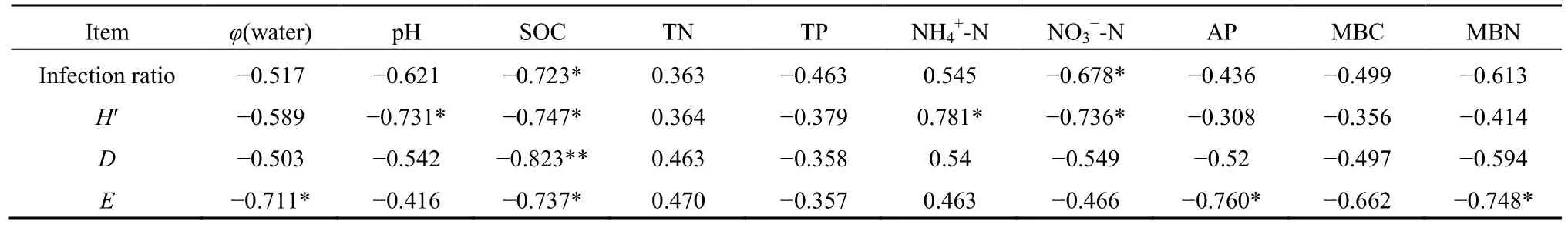
表4 菌根侵染率、多樣性指數與土壤理化性質的相關性Table 4 Correlation of mycorrhizal infection ratio, diversity index and soil physical and chemical properties
此外,從圖5可以進一步看出,不同坡位的林地土壤中,隨著土壤有機碳含量的增大,菌根真菌的侵染率、Shannon-Wiener指數和豐富度指數均顯著地線性下降,其中侵染率和豐富度指數的下降幅度極為明顯。而從圖6可以看出,土壤硝態氮含量是決定菌根侵染率和菌根真菌 Shannon-Wiener指數的另一重要影響因子,隨土壤硝態氮含量的升高,菌根真菌侵染率和Shannon-Wiener指數也均呈現顯著的線性下降趨勢。另外,菌根真菌的豐富度指數還隨著土壤微生物生物量氮含量的增大呈現顯著的線性下降(圖7)。
3 討論
本研究通過對亞熱帶北緣丘陵山區處于不同坡位的天然恢復的次生闊葉林所進行的調查和研究結果表明,坡位對林地土壤的理化特性產生了一定的影響。隨著坡位的上升,林地土壤的含水率及pH有所下降,上坡林地土壤的硝態氮、有機質、和全氮含量均顯著低于中、下坡位,土壤無機氮和全磷含量也表現為上坡低于下坡,總體上隨著坡位的上升,立地條件顯著下降。這一結果與前人的研究結果基本一致(侯曉麗等,2013;韋建宏等,2017;張順平,2014)。不同坡位由于雨水沖刷程度、水分下滲、蒸騰蒸發、溫度變化以及養分分解和淋溶等方面的作用和效果不同產生一定的生活因子再分配作用,是造成其土壤水分和養分差異的主要原因(李奕等,2014;薛立等,2011)。本研究區多雨和濕潤的氣候,雨水長期的沖刷和下滲作用將上坡的許多礦物風化產物、以及凋落物及其分解產物向下坡方向不斷淋洗和遷移,此外,由于下坡具有更好的水熱條件,凋落物的分解速度更快,因此,下坡具有更好的養分和水分條件。與此相對應,上坡土壤微生物生物量碳的含量顯著低于下坡,且土壤微生物生物量氮的含量也隨坡位上升呈現減小的趨勢。土壤微生物對環境十分敏感,受到氣候條件、土壤質地、土壤養分含量等諸多綜合因素的影響(成毅等,2010)。但從土壤微生物生物量C/N比來看,本研究林地土壤微生物生物量C/N比在不同坡位并不存在差異,均在3左右,表明該區域次生闊葉林中進行凋落物分解的土壤微生物以細菌占優勢,凋落物質量和分解情況總體較好。
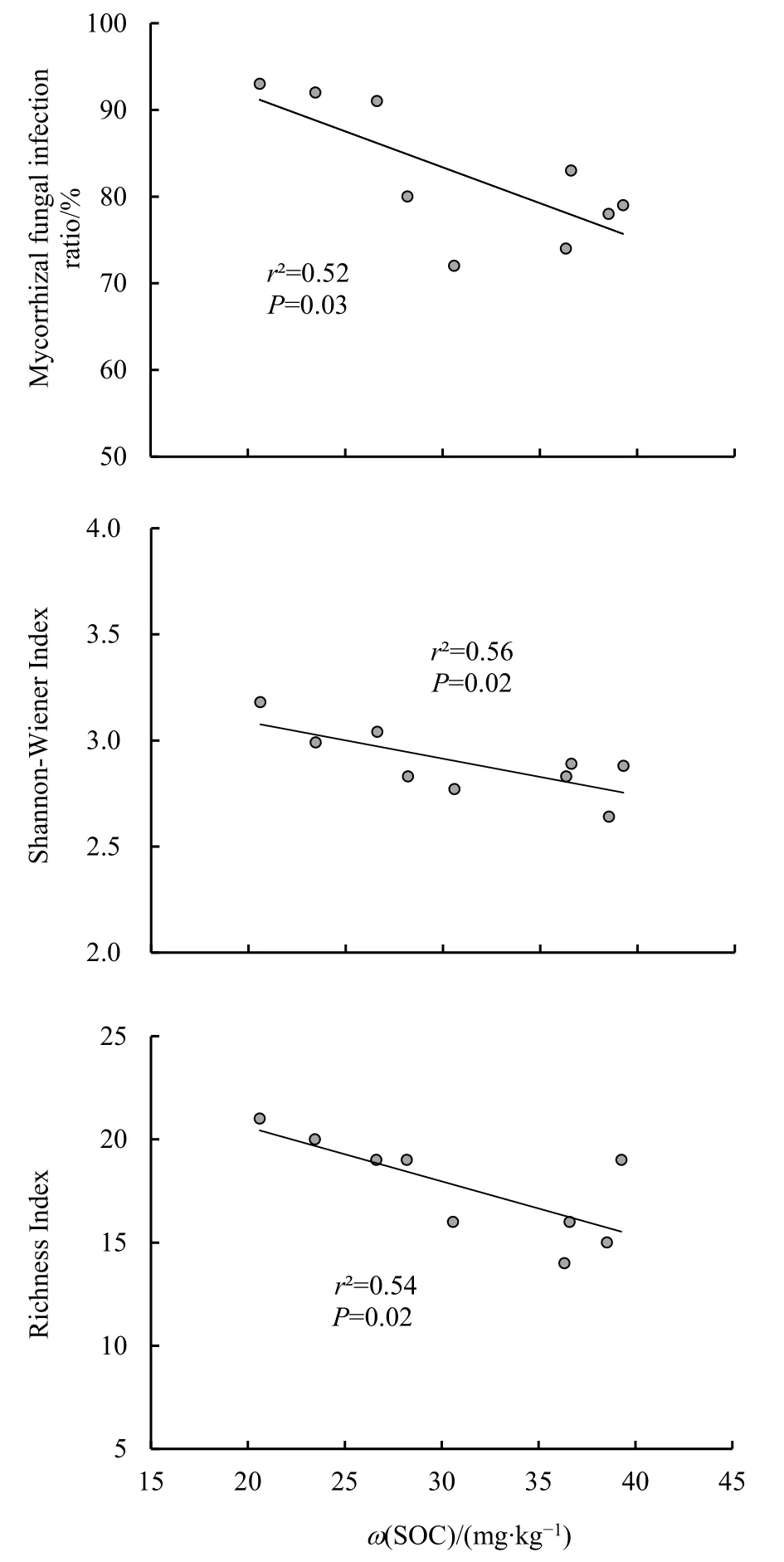
圖5 菌根真菌侵染率及多樣性指數與土壤有機碳含量關系Fig. 5 Relationship between infection ratio and diversity indexes of mycorrhizal fungi with soil organic carbon content

圖6 菌根真菌侵染率及Shannon-Wiener多樣性指數與土壤硝態氮含量關系Fig. 6 Relationship between infection ratio and Shannon-Wiener index of mycorrhizal fungi with soil nitrate nitrogen content
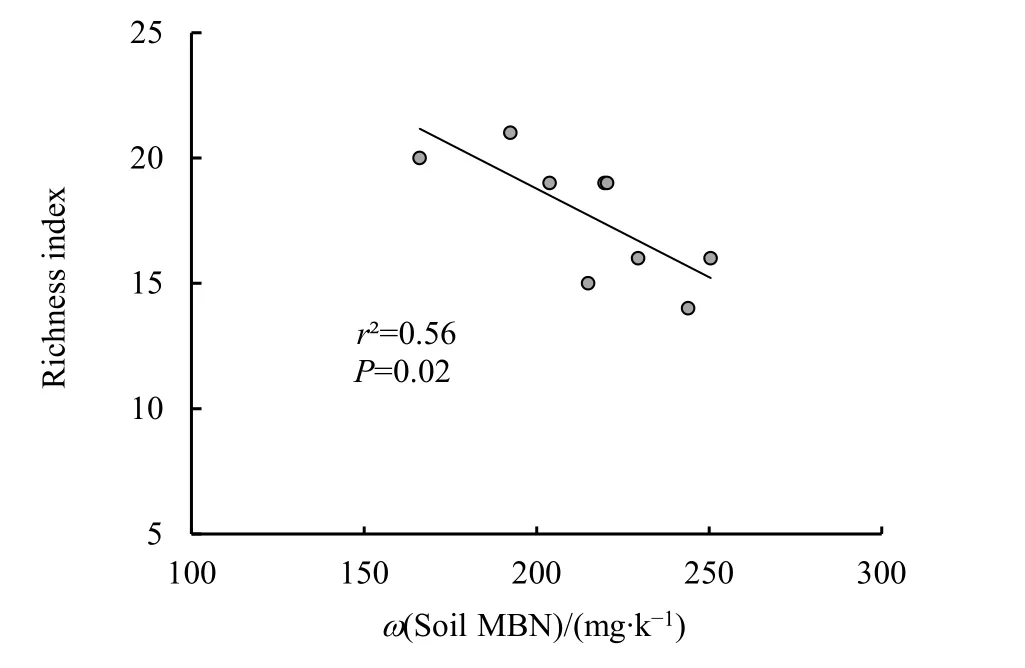
圖7 菌根真菌豐富度指數與土壤MBN含量關系Fig. 7 Relationship between richness index of mycorrhizal fungi with soil microbial biomass nitrogen content
本調查地不同坡位的菌根真菌侵染率與菌根真菌群落多樣性變化規律相似,林地上坡菌根侵染率和Shannon-Wiener指數、Simpson指數、豐富度指數均顯著大于中坡和下坡。宋福強等(2006)針對亞高山地區森林的研究表明,坡度較小的林地菌根侵染率低于坡度較大的林地。本研究涉及的3個調查林地整體上從上坡往下坡坡度明顯減小(表1),在菌根侵染率方面的研究結果與宋福強等(2006)的結果一致。大量的研究表明,物種的數量和分布均勻程度能夠比較全面地反映物種的群落分布(劉林馨等,2018)。本研究中,上坡菌根真菌的Shannon-Wiener指數、Simpson指數、豐富度指數最大,與上坡菌根真菌侵染率最高相對應。說明上坡菌根真菌群落最豐富,林木根系受到的侵染程度更高。林木菌根真菌的侵染狀況取決于宿主植物和菌根種類的的生物學特性以及土壤因子、水分和溫度等綜合因素的共同影響(孫婧玨,2012;韋莉莉等,2016;吳勇等,2013)。本研究針對次生闊葉林所進行的研究結果表明,與中、下坡相比,上坡受養分淋溶、沖刷等的影響,林地土壤的有機碳以及氮素相對缺乏,土壤肥力較低,而與之相對應,上坡的菌根真菌侵染率以及真菌的多樣性均顯著高于中、下坡,說明土壤有機質以及氮素的供應程度的高低在一定程度上可能成為林木與菌根真菌共生的重要誘因。從林木可以直接吸收利用的無機氮組分來看,硝態氮含量總體上與菌根真菌的侵染率以及多樣性呈現顯著的負相關關系(圖 6)。與銨態氮相比,硝態氮在土壤中移動能力強,對林木而言可利用程度更高(陳永亮等,2014;宋海星等,2005)。土壤硝化過程主要受土壤氮礦化水平和銨態氮供應的影響(Reynold et al.,2010;Smiciklas et al.,1992),此外,參與硝化作用的專性微生物對環境有相對較高的要求(Van et al.,2010)。本研究地上坡相對較差的土壤環境以及較低的土壤有機碳、氮含量在一定程度上可能限制了土壤氮的礦化過程以及銨態氮的生成,從而抑制了硝化微生物的活性以及硝化作用;另一方面,雨水的淋洗作用也容易導致土壤硝態氮從上坡向中、下坡的流失,從而導致上坡林木土壤硝態氮的供應顯著降低,從而促進了林木與菌根真菌在更大程度上的共生,并相應提高了菌根真菌的多樣性,通過多樣化的菌根真菌的菌絲擴張增強對養分的吸收能力,維持林木的正常生長。從這一層面看,土壤硝態氮的供應水平可能是決定菌根真菌共生更為重要的因子。
本研究林地土壤微生物生物量氮含量與菌根真菌的豐富度呈現顯著的負相關(圖 7)。上坡較低的土壤微生物生物量氮含量以及與下坡基本一致的MBC/MBN(圖1)進一步表明了上坡土壤氮的缺乏,需要有更多樣而豐富的菌根真菌的共生來維持上坡林木的養分需求。此外,上坡較低的土壤MBN含量在一定程度上也表明土壤微生物區系與中、下坡相比在向較低氮素需求的群體轉變,這一轉變有可能會進一步影響土壤氮素的轉化過程和供應,因此,將來還需要針對土壤微生物區系的變化進行相應研究,從而深入全面地把握坡位對立地條件的影響,為次生闊葉林的自然恢復以及生態系統的可持續管理提供進一步的數據支撐。
4 結論
亞熱帶北緣丘陵山區次生闊葉林天然恢復過程中,坡位對林地土壤的養分含量和分布存在一定影響,并決定了不同坡位林木的菌根侵染特征。上坡土壤有機碳含量、全氮含量以及硝態氮含量顯著低于中、下坡,其他養分含量在不同坡位之間差異不顯著,但基本上均隨坡位上升而下降。土壤微生物生物量碳、氮含量也均以下坡最高,能夠有效反映林地地力隨坡位的變化。該地區不同坡位的次生闊葉林均存在較高的菌根真菌侵染率(74.6%-92.1%),但與土壤養分和微生物生物量的變化規律相反,上坡的侵染率顯著高于中、下坡。菌根真菌的多樣性與其侵染率的變化規律相似,Shannon-Wiener指數、Simpson指數和豐富度指數均為上坡大于中、下坡。土壤有機碳、全氮以及硝態氮的含量是該地區次生闊葉林菌根真菌侵染和多樣性形成的重要決定性因子。
