凍融循環對亞高山森林土壤微生物生物量及群落結構的影響
2019-05-31 08:55:10諶亞賈雨橋王欣宇楊萬勤吳福忠張麗衛芯宇陳子豪楊玉蓮譚波
生態環境學報 2019年4期
諶亞 ,賈雨橋,王欣宇,楊萬勤,吳福忠,張麗,衛芯宇 ,陳子豪,楊玉蓮 ,譚波*
1. 四川農業大學生態林業研究所/長江上游林業生態工程四川省重點實驗室/高山森林生態系統定位研究站/水土保持與荒漠化防治省級重點實驗室,四川 成都 611130;
2. 綿陽師范學院/生態安全與保護四川省重點實驗室,四川 綿陽 621000
土壤微生物是土壤生態系統中重要的組成部分,其主要以地上和地下凋落物、根系分泌物、木質殘體以及動物殘骸為碳源,在森林凋落物的分解、土壤肥力的形成與維持、養分循環與碳氮周轉等生態過程中發揮著重要作用(Schloter et al.,2003;Green et al.,2004)。土壤微生物對外界環境的變化極為敏感,其生物量和群落結構變化是土壤特征變化的早期響應指標(Zelles et al.,1992),被認為是比土壤總有機質更可信的環境變化指示因子(Chen et al.,2008)。
土壤的季節性凍融是全球中、高緯度或高海拔地區生態系統的普遍現象(孫輝等,2008),短期的凍融循環更是季節性凍融過程中重要組成部分,其對土壤微生物的影響可能更為明顯。凍融過程會改變土壤結構、破壞土壤團聚體和凋落物中微生物及動植物殘體細胞,但其釋放出的底物和養分又為存活的土壤微生物提供了有效基質(Groffman et al.,2001;Herrmann et al.,2002)。普遍認為,凍融循環對土壤微生物群落的作用機制可能包括兩方面,(1)凍融的致死作用。冬季土壤凍結和凍融循環作用能降低土壤微生物活性甚至將其致死(Urakawa et al.,2014)。(2)養分供應不足。隨著凍融循環次數的增加,從土壤團聚體和凋落物中釋放出來的可溶性養分下降,導致微生物生物群落發生變化(Cleveland et al.,2014;Herrmann et al.,2002)。因此,凍融期間強烈的環境變化可能極大地影響著土壤微生物群落的組成和結構。
川西亞高山森林是中國西部高山林區的主體,在氣候調節、水土保持、水源涵養、生物多樣性保育和全球碳循環等方面具有不可替代的作用和地位(Yang et al.,2005)。受青藏高原隆起影響,該區森林土壤每年具有明顯的季節性凍融過程(Yang et al.,2007)。凍融循環研究對于預測全球氣候變化背景下,土壤生態系統對凍融特征格局變化的反饋有非常重要的科學意義,但作為全球氣候變化的敏感區域,目前有關亞高山森林土壤微生物對凍融循環變化響應的研究還較少,這不僅限制了對亞高山森林土壤微生物群落及亞高山森林生態系統物質循環和能量轉換過程的了解,還很難滿足全球變化與區域響應研究的需要。因此,以受季節性凍融影響明顯的川西亞高山森林土壤為研究對象,采用微縮實驗,模擬凍融循環對亞高山森林土壤微生物的影響,以期為深入理解全球變暖背景下亞高山森林生態系統過程對季節性凍融變化的響應提供參考。
1 材料與方法
1.1 研究區域概況
研究區域位于四川省西部,綿陽市平武縣王朗國家級自然保護區(32°49′-33°02′N,103°55′-104°10′E,海拔2300-4980 m),地處青藏高原與四川盆地的過渡地帶。該區域屬丹巴-松潘半濕潤季風氣候,干濕季節差異明顯,年降水量 801-825 mm,主要集中在5-8月,年平均溫度為2.9 ℃,土壤季節性凍融期長達 5-6個月。研究區域內優勢喬木為岷江冷杉(Abies faxoniana)、川西云杉(Picea balfouriana)、白樺(Betula platyphylla)、紅樺(Betula albo-sinensis)等,優勢灌木為康定柳(Salix paraplesia)、高山杜鵑(Rhododendron lapponicum)、缺苞箭竹(Fargesia denudata)等,土壤為棕壤和暗棕壤(Yang et al.,2005;衛芯宇等,2018)。
1.2 試驗設計
2017年5月采集供試土壤,在王朗國家級自然保護區的針闊混交林、針葉林以及闊葉林內分別選取3個坡向、坡度相近的樣地(表1)。在每個樣地內隨機布設1個5 m × 5 m的樣方,收集各樣地內的凋落葉,土壤采樣時,先將土壤表面的凋落物及苔蘚等地被物清理干凈,通過5點取樣法,以長寬各為30 cm的方形采集0-10 cm土壤,各林型分別取得 15份樣品,混合均勻后取部分樣品帶回實驗室,剔除根系和雜質,一部分用于測定土壤理化性質,另一部分過2 mm篩,存于4 ℃冰箱待用。對應收集各樣地的凋落葉,風干備用,同時取一部分凋落葉測定其理化性質。土壤及凋落葉的理化性質詳見衛芯宇等(2018)。
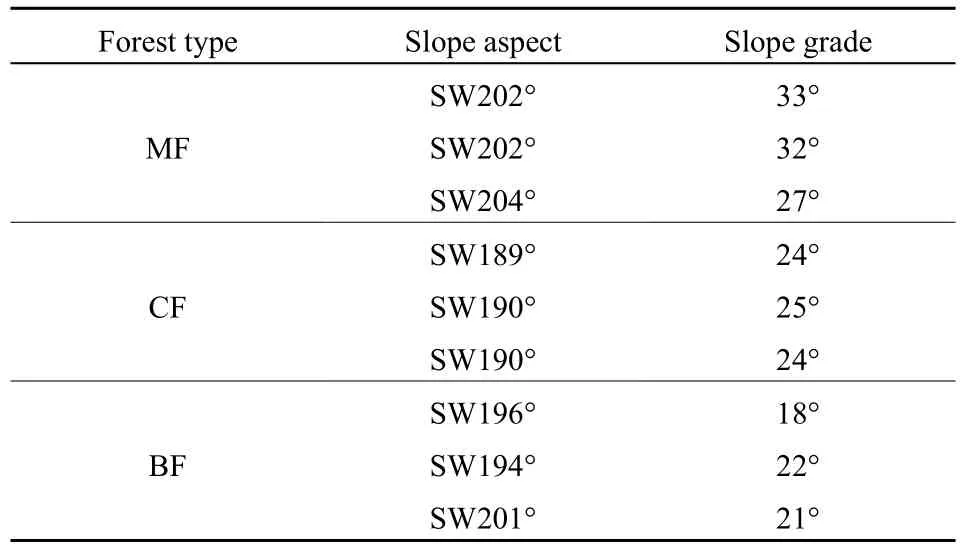
表1 樣地的坡度和坡向Table 1 Slope aspect grade of the sampling sites
每個樣地各稱取250 g過篩新鮮土樣,置于350 mL培養罐中。土壤含水量統一調節為60%田間持水量。為了解凍融循環對凋落葉分解過程中土壤微生物群落的影響,模擬表層土原始狀態,在每一類型土壤中添加凋落葉,依據課題組前期對該地區各森林類型凋落葉年凋落量的調查研究(Yang et al.,2006),計算該地區單位面積上的實際凋落葉凋落量,針葉林、針闊混交林和闊葉林分別為 324.4、246.8和 171.2 g·m-2,再根據培養罐的橫截面積(50.24 cm2)確定凋落葉的添加量,每個處理設置3個重復。本試驗添加的針葉林、針闊混交林和闊葉林凋落葉有機碳含量分別為 348.00、342.86和452.25 g·kg-1(衛芯宇等,2018),碳含量差異不顯著,便于進行不同林型間研究結果的比較。基于前期Wu et al.(2010)的觀測數據和溫度動態特征,該地區冬季土壤凍結時和土壤融化時溫度分別長時間維持在-5 ℃和5 ℃左右,預實驗結果表明,在-5 ℃條件下凍結12 h,5 ℃條件下解凍12 h,能保證土樣凍結及完全解凍,所以將這樣的一天定義為1個凍融周期,共80次。用保鮮膜封住培養罐口防止其他雜質的輸入,并用針在保鮮膜上扎多個小孔,保持培養罐內外空氣流通。
1.3 樣品采集與分析
分別在凍融循環培養的第 5、10、20、40、80個周期取樣并測定各項指標。PLFA提取的所有操作均在通風櫥中進行。提取過程參考 Bossio et al.(1998)的方法加以改進,進行單項萃取。稱取(1.00±0.05) g樣品裝入30 mL聚四氟乙烯的棕色玻璃瓶內,依次加入提取液PBS 4.8 mL(磷酸鉀緩沖液,pH 7.4)、甲醇(HPLC)12 mL、三氯甲烷(HPLC)6 mL,渦旋30 s,超聲10 min(37 ℃,100 Hz),37 ℃水浴30 min,冷卻后倒入50 mL廣口三角瓶中,加6 mL三氯甲烷(HPLC)和6 mL PBS,輕輕搖勻后靜置,避光過夜。吸取下層氯仿相液體,過0.45 μm孔徑的有機纖維濾膜,再過經5 mL三氯甲烷(HPLC)活化的硅膠柱。依次用(2 mL×3次)三氯甲烷(HPLC)和(2 mL×3次)丙酮(HPLC)除去非極性磷脂和糖脂。將用甲醇(HPLC)(2 mL×3次)洗脫的極性磷脂用N2吹干,提取物用1 mL堿性甲醇(質量濃度為5.6 g·L-1KOH甲醇溶液)和1 mL 甲苯-甲醇溶液(V/V=1∶1)分解,渦旋 30 s,37 ℃水浴30 min,冷卻后用1 mol·L-1乙酸中和,生成的FAMEs(脂肪酸甲酯)再用(2 mL×2次)三氯甲烷-正已烷(V/V為 1∶4)提取,氮氣吹干。吹干后的提取物用200 μL三氯甲烷-正已烷(V/V為1∶4)溶解,再用島津QP2010GC-MS分析儀(GC/MS,Model QP-2010,Shimadzu,Japan)檢測。
土壤微生物群落結構分析采用磷脂脂肪酸(PLFA)生物標記法。PLFA采用標準命名法進行分類命名,分子式以“X:YωZ(c/t)”表示,其中,X表示脂肪酸分子的碳原子總數,Y表示不飽和烯鍵數,ω表示烯鍵距羧基的位置,Z表示烯鍵或環丙烷鏈的位置;前綴i(iso)和a(anteiso)分別代表順式異構和反式異構;cy代表環丙基;Me代表甲基;OH代表羥基;后綴c(cis)和t(trans)分別代表順式和反式同分異構(Frosteg?rd et al.,1993)。表征各微生物種群的磷脂脂肪酸標記物見表 2,采用各類群標記物生物量之和,計算真菌/細菌和革蘭氏陽性菌/革蘭氏陰性菌比值。
1.4 數據處理和統計分析
PLFAs為樣品中各類微生物的PLFA之和。引入生物多樣性指數:Shannon-Wiener多樣性指數(H)、Pielou均勻度指數(J)、Margalef豐富度指數(SR)和Simpson優勢度指數(D)來分析土壤微生物多樣性。其中,
多樣性指數:

均勻度指數:

豐富度指數:

優勢度指數:

式中,ni為每種 PLFA的質量摩爾濃度,nmol·g-1;N為 PLFA 總質量摩爾濃度,nmol·g-1;S為PLFA種類數(常晨暉等,2014)。
運用 Excel 2016和 SPSS 20.0(IBM SPSS Statistics,Chicago,IL,USA)軟件對數據進行統計分析。采用一般線性模型對3種林型各指標進行重復測量方差分析,單因素方差分析(one-way ANOVA)對同次取樣不同林型和同林型不同凍融循環次數各指標進行方差分析,LSD法進行多重比較(α=0.05)。運用 Origin Pro 2018 軟件作圖,Canoco for Windows 5.0分析微生物群落結構與環境因子的關系。
2 結果與分析
2.1 土壤微生物生物量
重復測量方差分析結果表明(圖 1),凍融循環(FTC)對細菌、革蘭氏陽性菌、革蘭氏陰性菌及真菌 PLFAs含量和真菌/細菌比值都有顯著影響(P<0.05)。3種森林土壤中細菌、革蘭氏陽性菌和革蘭氏陰性菌的 PLFAs含量呈現隨凍融循環次數增加而逐漸降低的趨勢(圖1a-圖1 c),真菌PLFAs含量和真菌/細菌比值則隨凍融循環次數增加表現出先降低后在第 40次凍融循環時升高再降低的趨勢(圖1d和圖1e),其中真菌/細菌比值僅闊葉林存在顯著差異(P<0.05),在第5次凍融循環時比值最大,在第 20次凍融循環時最低。凍融循環次數和林型對革蘭氏陽性菌/革蘭氏陰性菌都無顯著影響。
單因素方差分析結果表明,不同林型的土壤細菌和革蘭氏陽性菌的PLFAs含量在第5次和第80次凍融循環時才有顯著差異(P<0.05),且混交林顯著高于闊葉林,表現為混交林>針葉林>闊葉林。混交林、針葉林和闊葉林第80次凍融循環的細菌PLFAs含量分別較第5次低71.9%、73.7%、74.6%,革蘭氏陽性菌PLFAs含量分別較第5次低77.1%、68.8%、73.7%(圖1a和圖1b)。各林型的革蘭氏陰性菌和真菌的 PLFAs含量在各個測定凍融循環周期差異都不顯著,混交林、針葉林和闊葉林在第80次凍融循環的革蘭氏陰性菌 PLFAs含量較第 5次低70.6%、78.1%、78.8%,真菌PLFAs含量較第5次則分別低 55.5%、75.4%、84.7%(圖 1c和圖1d)。

表2 表征特定微生物的PLFA生物標記物Table 2 PLFAs used in this study as biomarker
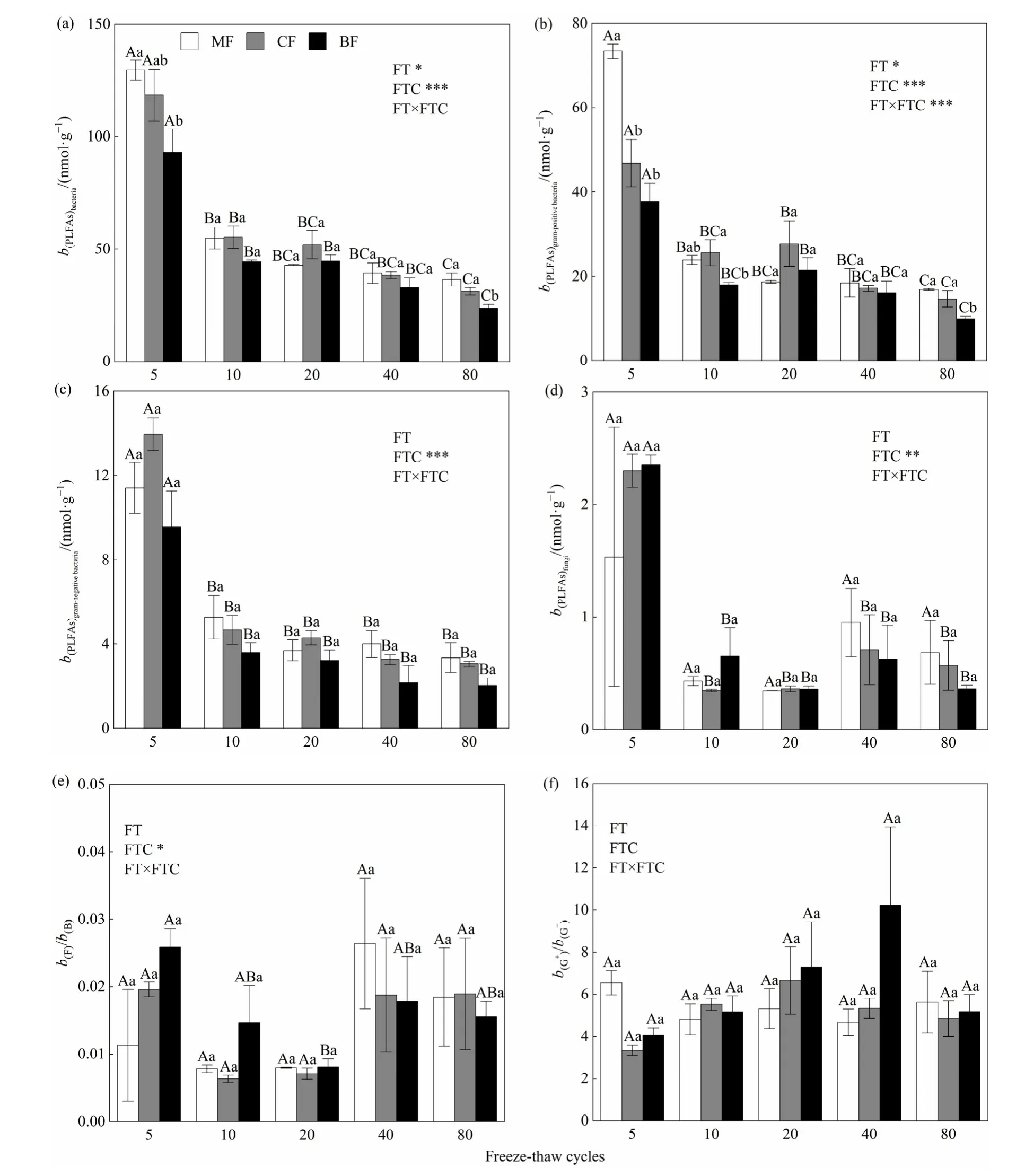
圖1 3種林型土壤微生物群落磷脂脂肪酸質量摩爾濃度及比值Fig. 1 Soil microbial PLFAs molality and ratio in three forest types.
2.2 土壤微生物群落多樣性
凍融循環對 3種林型的 Margalef豐富度指數(SR)、混交林和針葉林的Shannon-Wiener多樣性指數(H)和 Pielou均勻度指數(J)及混交林的Simpson優勢度指數(D)都有顯著影響(P<0.05)。混交林Shannon-Wiener多樣性指數和Pielou均勻度指數隨凍融循環次數增加而逐漸增加,針葉林和闊葉林均呈現下降趨勢,而Simpson優勢度指數則表現出與之相反的變化動態。3種林型的Margalef豐富度指數整體呈上升趨勢(圖2)。在第5次凍融循環時,混交林的 Shannon-Wiener多樣性指數和Pielou均勻度指數都低于針葉林和闊葉林,隨著凍融循環次數的增加,在第 80次凍融循環時,混交林和針葉林的Shannon-Wiener多樣性指數和Pielou均勻度指數都顯著高于闊葉林,而Simpson優勢度的結果完全相反,表現為混交林和針葉林顯著低于闊葉林。混交林的Margalef豐富度指數在第5次凍融循環時顯著低于針葉林和闊葉林,但隨著凍融循環次數的增加,各林型間的差異變得不顯著(圖2),說明可能隨著凍融循環次數的增加,土壤微環境中的土壤微生物種類趨于穩定。
3種林型5個凍融循環周期的指數相關性分析表明,多樣性指數與均勻度指數相關系數為0.913,呈極顯著正相關,優勢度指數與多樣性指數相關系數為0.899,與均勻度指數相關系數為0.921,都呈極顯著負相關,而與豐富度指數的相關系數小于0.5(表3)。由此推測,微生物多樣性更易受均勻度和優勢度的影響,即與PLFA含量變化有關,尤其是優勢脂肪酸含量變化對多樣性指數影響較大。
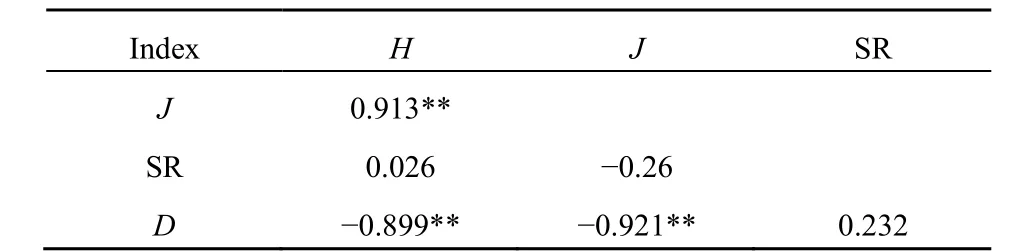
表3 不同林型下土壤微生物各指數相關性分析結果Table 3 Results of correlation analysis of soil microbial indices under different forest types

圖2 3種林型土壤微生物多樣性指數Fig. 2 Soil microbial diversity index of three forest types
2.3 土壤微生物群落結構與土壤環境因子的關系
冗余分析結果表明,前2軸共同解釋了86.5%的變異,其中第1軸的貢獻值為63.0%(圖3)。土壤環境因子中,銨態氮(F=5.3,P=0.008)和硝態氮(F=4.6,P=0.024)顯著影響了土壤微生物群落結構,表現為銨態氮和硝態氮與總微生物量、細菌、真菌、革蘭氏陽性菌和革蘭氏陰性菌 PLFAs含量呈顯著正相關,真菌/細菌比值與銨態氮呈正相關,與硝態氮呈負相關,革蘭氏陽性菌/革蘭氏陰性菌比值銨態氮呈負相關,與硝態氮不相關。

圖3 土壤微生物群落結構與土壤環境因子的冗余分析Fig. 3 Redundancy analysis (RDA) of relationships between soil microbial community structure and soil environmental factors
3 討論
凍融循環一般通過改變土壤水分的狀況及分布來直接影響土壤微生物(DeLuca et al.,1992),其會對土壤微生物生物量和群落結構造成影響,這在亞高山森林土壤中尤為明顯(張麗等,2015)。本研究中,相同林型下細菌、革蘭氏陽性菌、革蘭氏陰性菌和真菌的 PLFAs含量都隨著凍融循環次數的增加而降低,一方面可能是因為凍融循環直接致使部分土壤微生物死亡,降低了可培養微生物的多樣性(DeLuca et al.,1992;Walker et al.,2006),另一方面可能是因為本試驗有限的碳源(衛芯宇等,2018),使得微生物可利用的分解底物受限導致部分微生物死亡。在培養期間土壤細菌、革蘭氏陽性菌和革蘭氏陰性菌的 PLFAs含量都隨凍融循環次數的增加而逐漸降低,而真菌PLFAs含量則在第 40次凍融循環表現出升高趨勢,可能原因是此時土壤的水分遷移達到了一個動態平衡,土壤處于新的動態穩定平衡狀態(王大雁等,2005),真菌群落的耐受性高(Buckeridge et al.,2008),在環境穩定初期取得了生存優勢。同時,在第 40次凍融循環后,各林型的細菌、革蘭氏陽性菌、革蘭氏陰性菌和真菌的 PLFAs含量隨凍融循環次數增加都無顯著差異(圖1),都表現為混交林>針葉林>闊葉林,也可以說明此時凍融循環狀態下的土壤微環境是比較穩定的。
不同林型的凋落物物種組成不同,可改善林型的養分狀況,進而增加微生物可獲得資源的變異度(陳法霖等,2011),不同種類土壤微生物對這些變化的反應可能存在差異(李晶等,2013),導致土壤微生物群落結構和功能之間存在相互影響(Heemsbergen et al.,2004)。在第40次凍融循環后,混交林的細菌、真菌、革蘭氏陽性菌和革蘭氏陰性菌PLFAs含量都高于單一針葉林和闊葉林(圖1),表明與混交林相比,單一針葉林和闊葉林為土壤微生物群落提供生境和食物的能力較低,混交林凋落物更容易被土壤微生物群落利用從而合成自身的生物量,與胡亞林等(2005)的針葉與闊葉混合凋落物的分解能提高了微生物生物量的研究結果相似。
真菌/細菌和革蘭氏陽性菌/革蘭氏陰性菌的比值常被用作微生物群落整體結構變化的重要指標(Boyle et al.,2008),較大的真菌/細菌比值表明真菌生物量以及真菌菌絲體增多,與土壤有效養分的接觸面積增大,土壤養分元素的流失減少,因而土壤生態系統越來越穩定(Franciska et al.,2006)。凍融循環對真菌/細菌比值有顯著影響,且表現為先下降后在第40次凍融循環升高后降低的趨勢(圖1e),與真菌PLFAs含量的變化趨勢一樣,也說明在第40次凍融循環后土壤環境趨于穩定。從圖 1可以看出革蘭氏陽性菌PLFAs含量遠高于革蘭氏陰性菌,Margesin et al.(2009)研究表明革蘭氏陰性菌更易生長在營養豐富的土壤中,而革蘭氏陽性菌將在資源受限的土壤中占優勢,較高比例的革蘭氏陽性菌是土壤環境從富營養到寡營養的轉變(張地等,2012),說明在一個無外來物質循環和能量流動的環境中可利用資源是會逐漸減少的。
凍融循環對3種林型的Margalef豐富度指數、混交林和針葉林的 Shannon-Wiener多樣性指數和Pielou均勻度指數及混交林的Simpson優勢度指數都產生了顯著影響,混交林多樣性指數和均勻度指數隨凍融循環次數增加而逐漸增加,針葉林和闊葉林均呈現下降趨勢,而優勢度指數則表現出與之相反的變化動態。3種林型的豐富度指數整體呈上升趨勢(圖 2)。頻繁的凍融循環使微生物群落處于一個極不穩定的狀態,凋落物受凍融循環的影響而破碎,剩余只能被部分微生物利用的物質為少數的微生物提供了有效基質(Campbell et al.,2005),不穩定的環境使得部分微生物總體數量減少,而低溫環境下存活的耐寒微生物一定程度影響了豐富度指數的變化(李俊等,2016)。嚴海元等(2010)研究認為多樣的凋落物能夠為微生物提供多樣豐富的基質來源,因而混合凋落物比單一凋落物有更多適應性的微生物存在,所以混交林多樣性指數和均勻度指數隨凍融循環次數增加而逐漸增加,針葉林和闊葉林均呈現下降趨勢,優勢度指數與多樣性指數和均勻度指數呈極顯著負相關。
此外,土壤氮的有效性常常能影響微生物的生長,環境中氮有效性越高,微生物的活性越強,繁殖也越快(Craine et al.,2007)。通常地,土壤凍融循環初期的土壤團聚體結構被破壞(Edwards,1991),大團聚體破碎成小團聚體,使土壤中原本不可被利用的養分釋放,增加了土壤的可利用養分(李忠佩等,2004;Freppaz et al.,2007)。因此,在第5次凍融循環后,細菌、真菌、革蘭氏陽性菌和革蘭氏陰性菌PLFAs含量最高。然而,隨著凍融循環次數的增加,分解底物逐漸被耗盡,微生物數量也逐漸減少(孫輝等,2008;張麗等,2015)。這與已有的研究(Freppaz et al.,2007;羅亞晨等,2014;張麗等,2015)以及冗余分析中土壤養分有效性與微生物生物量和群落結構顯著相關的結果一致(圖3)。
4 結論
凍融循環對土壤微生物群落的影響隨林型的不同而存在差異,與5次凍融循環相比,3種林型的細菌、真菌、革蘭氏陽性菌和革蘭氏陰性菌的PLFAs含量在經過80次凍融循環后均出現降低,在第 40次凍融循環后,土壤微環境可能處于一個動態穩定平衡狀態。凍融循環顯著影響闊葉林的真菌/細菌比值,林型對真菌/細菌比值和革蘭氏陽性菌/革蘭氏陰性菌比值影響不顯著。凍融循環對3種林型的豐富度指數、混交林和針葉林的多樣性指數和均勻度指數及混交林的優勢度指數都產生了顯著影響,耐寒性強的微生物在凍融循環作用下更具生存優勢。凍融過程中土壤養分有效性與微生物群落顯著相關,全球變暖導致冬季凍融循環格局變化可通過改變土壤養分有效性影響亞高山森林土壤微生物生物量和多樣性。
