生物炭添加對半干旱區土壤細菌群落的影響
2019-06-06 08:21:28孫層層周際海王彤彤鄭紀勇
中國環境科學 2019年5期
王 穎,孫層層,周際海,王彤彤,鄭紀勇,3*
?
生物炭添加對半干旱區土壤細菌群落的影響
王 穎1,孫層層1,周際海2,王彤彤1,鄭紀勇1,3*
(1.西北農林科技大學資源環境學院,陜西 楊凌 712100;2.南昌工程學院,江西 南昌 330099;3.西北農林科技大學,黃土高原土壤侵蝕與旱地農業國家重點試驗室,陜西 楊凌 712100)
以半干旱區固原生態試驗站生物炭修復4a的表層土壤為對象,采用高通量測序技術研究了不同添加類型(槐樹皮生物炭、鋸末生物炭)和比例(1%、3%、5%,質量百分比)的生物炭對土壤細菌多樣性及群落結構的影響.結果表明,生物炭應用提高了土壤細菌群落的多樣性,鋸末生物炭優于槐樹皮生物炭,且3%鋸末生物炭對細菌群落的多樣性影響最佳,其香農指數為6.22;優勢門主要為放線菌門(Actinobacteria)、變形菌門(Proteobacteria)、綠彎菌門(Chloroflexi)、酸桿菌門(Acidobacteria)和Saccharibacteria,相對豐度共占76.80%~85.31%;優勢綱有放線菌綱(Actinobacteria)、α-變形菌綱(Alphaproteobacteria)、酸桿菌綱(Acidobacteria),其相對豐度占48.13%~57.08%;屬水平上,施加生物炭增加了芽孢桿菌屬()、硝化螺旋菌屬()的相對豐度,降低了土微菌屬()、根瘤菌屬()的相對豐度;層級聚類及冗余分析(RDA)發現,施加生物炭對細菌群落結構有影響,其中,微生物量碳、含水率、銨態氮、有機碳對細菌群落結構的影響較大.細菌優勢門與環境因子相關性熱圖分析表明,銨態氮與放線菌門、綠彎菌門呈顯著相關性.銨態氮是影響細菌群落的主要理化因子.
生物炭;土壤細菌;多樣性;群落結構;高通量測序
生物炭在污染物吸附、土壤改良和結構改善等方面廣泛應用[1-4].微生物是維系土壤生態系統功能穩定的主導因子[5-6].且微生物對土壤環境的變化非常敏感[7-8],由于生物炭的施用改變了土壤理化性質,進而導致了微生物的變化[9-10].
研究表明,生物炭施用對土壤細菌群落產生影響,主要會改變細菌群落的α-多樣性和群落結構[11-13].Hu等[14]發現生物炭添加到土壤96d改變了土壤細菌群落結構,增加了細菌多樣性,且豐富了主要的細菌類群,如放線菌門. Dai等[15]研究發現熱解溫度和土壤類型的變化對主要類群影響不大.也有學者研究發現與對照相比,生物炭誘導形成了不同的細菌群落,且不同種類生物炭的誘導程度不同[16].目前大多數關于生物炭對土壤微生物群落影響的文獻報道都是短期試驗(1a以內),而生物炭中存在直接影響土壤微生物群落的不穩定物質,在短時間內可以被微生物迅速礦化[17],但也不能排除其長期應用后對環境的潛在影響.然而,與生物炭的短期效應相比,生物炭對土壤微生物長期影響研究較少,這限制了對生物炭長期應用后對生態效應的全局把控.
生物炭作為土壤添加劑,被廣泛應用于水土流失嚴重、生態環境脆弱的黃土高原半干旱地區[18].因此,本研究以半干旱區固原生態試驗站為定位試驗平臺,采用MiSeq測序技術和冗余分析方法,基于對單次施用生物炭后并保持4a的土壤細菌群落的研究,探討了不同添加類型和比例的生物炭對土壤細菌多樣性和物種組成的變化規律,闡述了環境因子對土壤細菌群落結構的影響,以期為科學施加生物炭,提高半干旱區地力和維持土壤微生物多樣性等提供參考.
1 材料與方法
1.1 試驗地概況與試驗設計
試驗區域概況及試驗設計方法參照文獻[19]. 2012年4月于中國科學院固原生態試驗站進行長期定位試驗小區的布設,該地區位于黃土高原中部偏西,屬于半干旱地帶,植被由半干旱典型草原向灌叢草原過渡.年均日照時數2518h,年降雨量472mm,無霜期 152d.土壤以黑壚土和湘黃土為主,顆粒組成為砂粒(0.02~2mm)含量為45.85%,粉粒(0.002~0.02mm)含量為34.47%,黏粒(<0.002mm)含量為19.68%.試驗所用的生物炭以槐樹皮、梧桐鋸末為原料高溫(550℃)熱解形成,均由陜西億鑫生物能源科技開發有限公司提供,其基本理化性質如表1所示.
槐樹皮生物炭(HB)和鋸末生物炭(JB)均設置3個添加比例(1%、3%、5%,質量百分比),試驗共設6個處理及一個空白對照,記為HB1、HB3、HB5、JB1、JB3、JB5、CK,每個處理3個重復,共有21個試驗小區.各小區采用完全隨機布設,小區規格為2.4m× 2.4m.生長季前清除土表雜草等雜物,剝離表層20cm深的土壤,將生物炭按設定比例完全混合再原位回填,生物炭一次性施用.
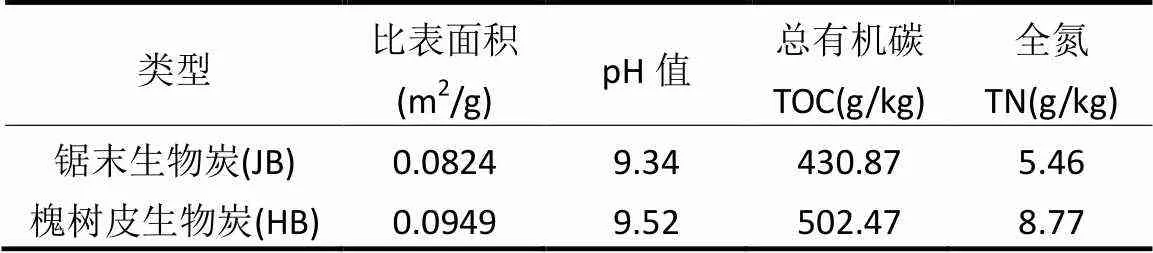
表1 供試生物炭基本理化性質
1.2 土壤樣品的采集
于2016年11月采集去除表層枯落物等雜物后的土壤(0~10cm),每個小區按照對角線采樣法采集5個點,混合并過2mm的篩,將部分土樣置于離心管中,用液氮罐保存,運回實驗室并儲存于-80℃冰箱中,用于土壤微生物的測定.另一部分鮮土樣用于硝態氮、銨態氮、含水量等土壤理化性質的測定.
1.3 指標測定
1.3.1 土壤理化性質測定 土壤含水量采用烘干法,土壤pH值采用電位法(水土比=2.5:1),土壤速效磷采用0.5mol/LNaHCO3浸提-鉬銻抗比色法,土壤全氮含量采用半微量凱氏定氮法,土壤總有機碳測定采用濃硫酸-重鉻酸鉀消煮-硫酸亞鐵滴定法.土壤銨態氮和硝態氮用1mol/L KCl 溶液浸提鮮土樣后,采用流動分析儀測定[20].微生物量碳采用氯仿熏蒸直接浸提法[21].
1.3.2 土壤DNA提取和16S rRNA基因高通量測序 土壤總DNA 使用美國OMEGA 公司的試劑盒,每個樣品稱取約0.5g新鮮土壤,按照試劑盒提取步驟進行.用338F(5’-ACTCCTACGGGAGGCAG- CAG-3’)806R(5’-GGACTACHVGGGTWTCTAAT-3’)引物對V3~V4區進行PCR擴增,每個樣品的上游引物5′端添加一段長度為8bp的特異性多肽 (barcode),用于區分樣品[22-23].用2%的瓊脂糖凝膠電泳檢測提取DNA 的純度和完整性,用核酸定量儀(Nano Drop ND-1000)檢測提取DNA的濃度和純度.根據Illumina MiSeq平臺(Illumina, SanDiego, USA)標準操作規程進行雙末端(Paired-end)測序,將純化后的擴增片段構建PE 2*300的文庫.PCR 采用TransGen AP221-02: TransStart Fastpfu DNA Polymerase; PCR儀為ABI GeneAmp? 9700型;每個樣本3個重復,將同一樣本的PCR產物混合后用2%瓊脂糖凝膠電泳檢測,使用AxyPrepDNA凝膠回收試劑盒(AXYGEN公司)切膠回收PCR產物,Tris_HCl洗脫.測序服務委托上海美吉生物醫藥科技有限公司完成.
1.4 數據處理與統計分析
1.4.1 基本數據處理 所有數據經過Excel 2013處理,利用SPSS 20.0進行單因素方差分析(One Way by ANOVA)和最小顯著差異法(LSD)比較,利用Origin8.5進行作圖.土壤因子與細菌種群分布特征之間的關系利用Canoco 4.5進行冗余分析(RDA),研究土壤性質對細菌分布特征的影響.
1.4.2 測序數據處理 MiSeq測序得到的是雙端序列數據,根據PE reads之間的overlap關系,將成對的reads拼接(merge)成一條序列,同時對reads的質量和merge的效果進行質控過濾,根據序列首尾兩端的barcode和引物序列區分樣品得到有效序列,并校正序列方向,得到優化數據.將優質序列聚類成操作分類單元(OTU),閾值設置為97%,軟件平臺: Usearch (vsesion 7.0 http://drive5.com/uparse/).為了得到每個OTU對應的物種分類信息,采用RDP classifier貝葉斯算法對97%相似水平的OTU代表序列進行分類學分析,并分別在各分類學水平統計各樣本的群落組成,將不能聚類在任何分類水平已知類群的序列定義為未分類.
2 結果與分析
2.1 土壤理化性質
不同生物炭處理下的土壤理化性質如表2所示,與對照相比,在pH值、總有機碳、速效磷、全氮、硝態氮、銨態氮、含水率7項指標中生物炭處理組均呈現出增大趨勢.施加生物炭4a后,土壤pH值、總有機碳、全氮隨著生物炭添加量的增加而顯著提高;銨態氮含量隨著生物炭添加量的增加而降低;無論HB或JB,添加量為3%時,速效磷含量最大;其他指標無統一變化規律.在相同添加量時,pH值、總有機碳、全氮、銨態氮和含水率5項指標均表現為HB大于JB;速效磷表現為HB小于JB.

表2 不同生物炭處理下的土壤理化性質
注:同列不同字母表示差異顯著(<0.05).
2.2 細菌α多樣性指數
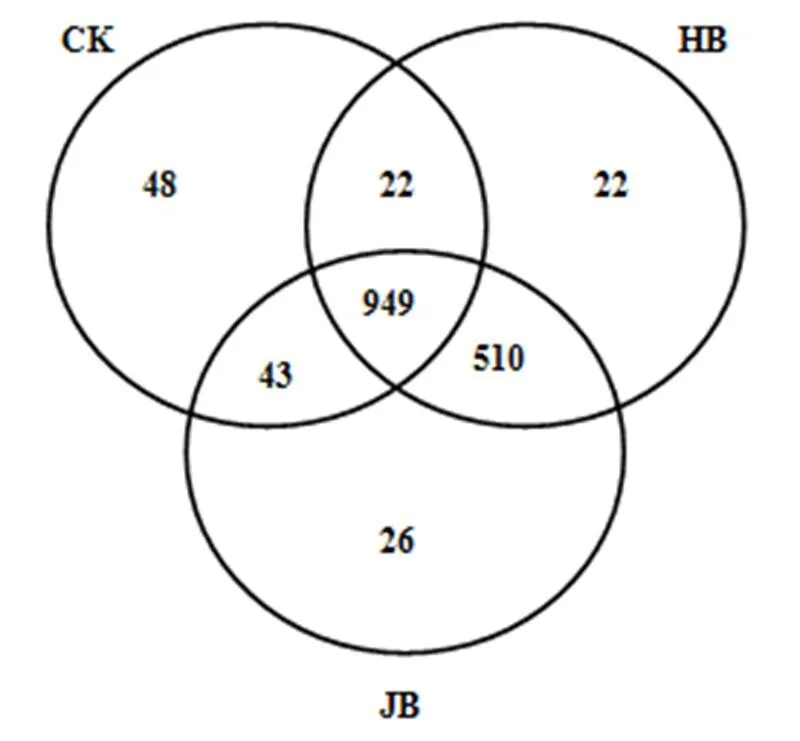
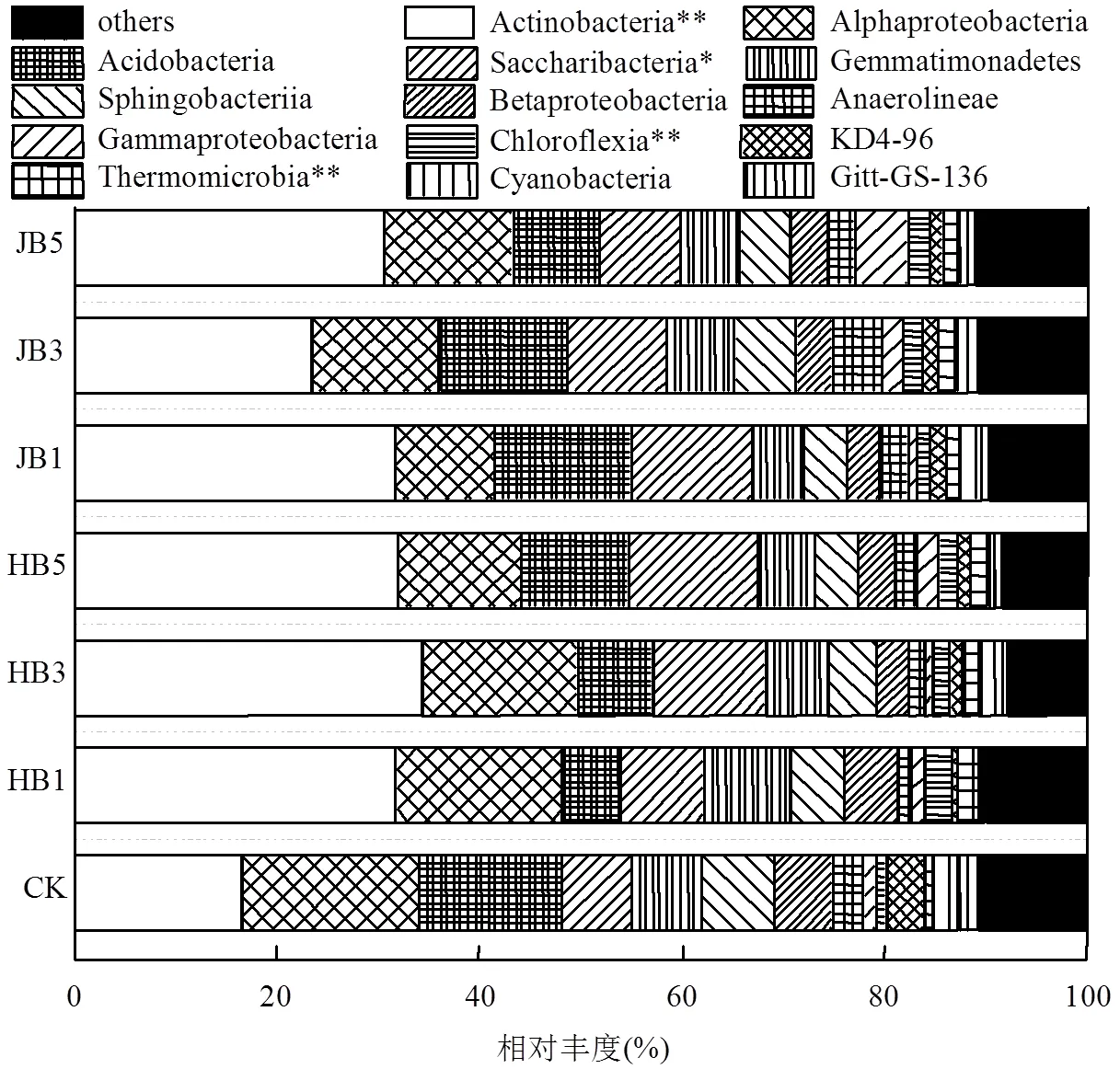
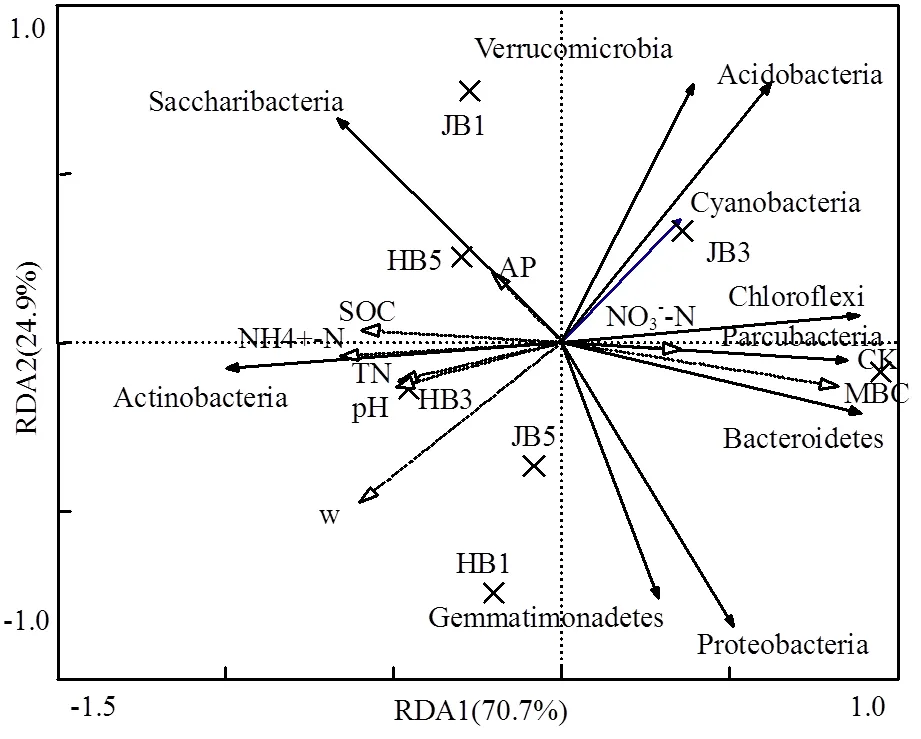
高通量測序數據及細菌多樣性指數如表3所示.ACE指數和Chao1指數可以表征物種的豐富度,對其而言,與對照相比生物炭處理組呈增大趨勢,說明生物炭可以提高細菌群落的物種豐富度;隨著施炭比例的增加,2種生物炭處理的豐富度指數均在3%施炭量達到最大.香農指數(Shannon)可以估算細菌群落的多樣性,表3呈現為JB3>HB1>JB5>JB1>HB5>CK>HB3,說明施加生物炭可以提高細菌群落的多樣性,其中,3%JB對細菌群落多樣性的影響效果最佳,為6.22.通過計算多樣性指數的平均值(JB=6.05, HB=5.91)、極大值(JB=6.22, HB=6.09)和中值(JB=6.02, HB= 5.88)發現JB>HB,說明相對而言JB更優.Smith Wilson's指數可以反應群落的均勻度,表2呈現為JB 表3 細菌16S r RNA 基因 Illumina Mi Seq 高通量測序數據及細菌多樣性指數 將所有樣品分為3組,槐樹皮生物炭(HB),鋸末生物炭(JB)和不施加生物炭(CK),如圖1所示. 3組微生物總OTUs數分別為1503,1528,1062個;3個處理組公有OTUs(949個)占HB、JB、CK 3個處理中總OTUs的比例分別為63.14%、62.11%和89.36%.HB、JB、CK 3個處理各自特有的OTUs數量占相應樣品總OTUs的比例為1.46%、1.70%和4.52%.由此可見,施加生物炭增加了土壤微生物物種的數量,與未施生物炭處理對比,施加生物炭處理土壤微生物組成相似性更高. 圖1 不同生物炭處理下細菌組成的Venn圖 *代表差異顯著(<0.05);**代表差異極顯著(<0.01),下同 在門水平上,共獲得35個類群,將相對豐度小于0.5%的類群歸類為其他,可得到13個類群,如圖2所示.其中,放線菌門(Actinobacteria)、變形菌門(Proteobacteria)、綠彎菌門(Chloroflexi)、酸桿菌門(Acidobacteria)和Saccharibacteria為優勢門,其相對豐度分別為16.42%~34.36%、15.28%~25.56%、10.06%~14.20%、5.68%~14.08%、6.91%~12.90%,這5類群微生物占CK、HB1、HB3、HB5、JB1、JB3、JB5相應處理總類群的76.80%、80.77%、83.28%、85.31%、83.15%、79.59%、82.70%.通過Illumina MiSeq高通量測序也檢測出相對豐度較小的細菌門,它們的相對豐度為0.5%~8%,如擬桿菌門(Bacteroidetes)、芽單胞菌門(Gemmatimonadetes)、硝化螺旋菌門(Nitrospirae)、厚壁菌門(Firmicutes).與空白對照(CK)相比,生物炭施用顯著提高了放線菌門(<0.01)、Saccharibacteria、硝化螺旋菌門、厚壁菌門(<0.05)的相對豐度,降低了酸桿菌門和疣微菌門的相對豐度. 在綱水平上,共獲得72個類群,將相對豐度小于2%的類群歸類為其他,共得到15個類群,如圖3所示.其中,放線菌綱(Actinobacteria)、-變形菌綱(Alphaproteobacteria)、酸桿菌綱(Acidobacteria)為主要類群,相對豐度分別為16.42%~34.36%、9.91%~ 17.63%、5.68%~14.08%,合計為48.13%~57.08%.與空白對照(CK)相比,施加生物炭顯著增加了Saccharibacteria、綠彎菌綱、熱微菌綱(Thermomicrobia)、放線菌綱的相對豐度(<0.05).-變形菌綱、酸桿菌綱、-變形菌綱的相對豐度隨著生物炭添加量的增加有降低的趨勢. 圖3 不同生物炭處理綱水平細菌組成 如圖4所示,施用生物炭增加了芽孢桿菌屬()、硝化螺旋菌屬()、氣微菌屬()的相對豐度,降低了土微菌屬()、細桿菌屬()、根瘤菌屬()的相對豐度,而慢生根瘤菌屬()則無明顯變化規律.隨著HB施用量的增加,芽孢桿菌屬、硝化螺旋菌屬、土微菌屬、根瘤菌屬、鞘氨醇單胞菌屬()的相對豐度均呈現下降趨勢,僅氣微菌屬呈上升趨勢;隨著JB施用量的增加,氣微菌屬呈現下降趨勢,與HB變化規律相反,說明生物炭對細菌屬水平的影響與生物炭種類相關. 圖4 不同生物炭處理8個屬水平功能細菌的相對豐度 圖5 不同生物炭處理細菌群落結構的層級聚類樹 如圖5所示,與對照相比,施加生物炭對細菌群落結構的影響顯著.其中, JB3、HB3、JB53個處理距離較近,說明對土壤細菌群落結構的影響相似.JB1和HB5處于一個分支,說明不同種類的生物炭在不同添加比例下對土壤細菌群落結構的影響可達到相同效果. HB1、HB3、HB5和JB1、JB3、JB5均處于不同的分支,說明生物炭的不同添加量對細菌群落結構有影響. 圖6 環境因子對細菌群落結構影響的冗余分析 如圖6所示,第一主軸和第二主軸分別解釋了微生物群落結構與土壤理化性質關系總變異的70.7%和24.9%.不同處理的樣本點相對較分散,說明不同生物炭處理對細菌群落結構的影響不同.除JB3樣本點外,生物炭處理的其他樣本點與對照組CK在RDA1軸方向上明顯分離.此外,在所選環境因子中,微生物量碳的解釋度最高,達到了25.85%,含水率、銨態氮和有機碳分別解釋了16.78%、16.61%、13.68%.如圖7所示,銨態氮與放線菌門呈顯著正相關關系,與綠彎菌門呈顯著負相關關系.含水量與芽單胞菌門(Gemmatimonadetes)、硝化螺旋菌門(Nitrospirae)、擬桿菌門(Bacteroidetes)呈負相關關系,有機碳和全氮與絕大部分菌群呈負相關關系. 圖7 細菌優勢門與環境因子相關性熱圖 生物多樣性指數用于描述生物類型數和均勻度,它可反映生物群落中物種的豐富度及其各類型的分布比[24].本研究結果顯示,生物炭添加提高了土壤細菌的豐富度且3%添加量的提高程度最大,生物炭添加提高了土壤細菌的多樣性,1%HB和3%JB添加量最佳,但這和豐富度的結果不一致,主要是因為細菌多樣性由物種豐富度和均勻度共同決定,細菌多樣性結果會因豐富度和均勻度二者所占比重而有所不同[25].Doan等[26]、Nguyen等[27]研究生物炭施用對土壤細菌特性的影響發現添加生物炭后土壤細菌的多樣性提高,均與本研究結果相似.此外,生物炭的添加在一定程度上可改善土壤微環境,這可能有益于個別類群的生長,從而引起細菌群落個體大小或數量的差異,群落結構發生變化且均勻度降低,進而導致多樣性指數減小,細菌群落的多樣性降低[28-29].由于生物炭巨大的比表面積和疏松多孔的結構為細菌生存提供生長空間和附著位點[30],同時生物炭添加也改變了土壤的理化性質,從而影響細菌的生長代謝,直接或間接地改變了土壤的細菌多樣性.通過計算多樣性指數的平均值、極值和中值發現,JB>HB,說明生物炭種類不同,其內部結構不完全一樣,微生物對不同生物炭引起的環境變化的適應性會有差異,所以,生物炭對細菌多樣性的影響結果不同.生物炭對細菌多樣性的影響程度也因為作用時間不同而產生差異.研究結果顯示,生物炭短期施用會顯著改變細菌的多樣性[27],而隨著時間的推移,生物炭對細菌多樣性影響程度降低,這可能由于生物炭短期應用于土壤中,會對土壤有激發效應,土壤中氮磷鉀含量均會顯著提高[31],促進了土壤微生物的發育繁殖;長期應用后,土壤環境和外源生物炭逐漸融合,生物炭成為土壤環境的一部分且達到一個穩定的狀態,可能不再滿足某類微生物的生長繁殖,微生物多樣性很難再發生變化. 生物炭對不同分類學層次菌群組成的影響不完全相同.本研究中,門水平優勢菌群為放線菌門、變形菌門、綠彎菌門、酸桿菌門和Saccharibacteria,這和黃土高原地區以及東北地區關于土壤細菌群落的研究結果均相似[32].與對照相比,生物炭添加顯著提高了放線菌門、硝化螺旋菌門、厚壁菌門的相對豐度,降低了酸桿菌門和疣微菌門的相對豐度,原因可能是放線菌門類群隨著土壤pH值下降而豐度降低[33],且放線菌門為富營養類群,能利用速效碳源快速生長[34].生物炭的施用提高了土壤pH值,從而使放線菌門類群在含有生物炭的土壤環境中較好的生長繁殖;硝化螺旋菌門是參與亞硝酸鹽的氧化和碳氮循環的主要類群,生物炭通過改變土壤基本理化性質及在土壤中的降解過程,直接或間接的增加了硝化螺旋菌門的相對豐度[35];酸桿菌門的細菌多屬于寡營養類群[36],土壤的富營養狀態不利于該類菌群的生長[37],生物炭施用改善了土壤養分狀況,雖未達到富營養狀態,但土壤環境較之前發生了變化,也抑制了該類群微生物的生長,另一方面,酸桿菌門細菌更喜歡酸性環境[38],生物炭本身具有較高的pH值,施加生物炭改變了土壤環境的酸堿度,所以酸桿菌門的相對豐度降低.本研究中綱水平優勢菌群變化與門水平類似.其中,生物炭施用降低了-變形菌綱、-變形菌綱的相對豐度,且-變形菌綱的相對豐度大于-變形菌綱,這與前人研究結果一致[39].變形菌綱細菌可在酸性環境中利用難降解的碳源并轉化為中間小分子物質為其他微生物提供養分[40],而該半干旱區土壤呈堿性且生物炭也增加了土壤pH值,阻礙了-變形菌綱菌群的發展,從而使-變形菌綱的相對豐度降低.此外,本研究還發現生物炭添加改變了部分功能屬水平細菌的相對豐度,如生物炭增加了芽孢桿菌屬、硝化螺菌屬、氣微菌屬的相對豐度,降低了土微菌屬、細桿菌屬、根瘤菌屬的相對豐度.研究表明,芽孢桿菌屬是一種能將植物難以吸收利用的磷轉化為可吸收利用的形態[41]、促進植物生長、防治植物病害[42]的一種根際菌屬.水分和植被是影響芽孢桿菌的兩個重要生態因子[43],生物炭施用增加了土壤的含水量,促進了植物生長,間接增加了芽孢桿菌屬的相對豐度,這與Yao等[11]的研究結果相似.慢生根瘤菌屬和根瘤菌屬一般與草本豆科植物關聯在一起,Zahran等[44]研究發現草本植物在生長過程中易與相容的根瘤菌屬菌株形成有效的根瘤共生體,具有很好的固氮作用.慢生根瘤菌屬與根瘤菌屬變化趨勢不同,因為其與根瘤菌屬在系統發育關系中并不屬于同一個分支[45];鞘氨醇單胞菌屬具有很強的生物降解能力且對芳香化合物具有高代謝能力,該菌株被廣泛應用于污染物修復方面[46].生物炭添加對鞘氨醇單胞菌屬相對豐度的影響可能與生物炭的種類相關,槐樹皮生物炭促進了該菌群的生長,相反地,鋸末生物炭對鞘氨醇單胞菌屬產生了抑制作用,相對豐度稍有降低,這與陳澤斌[47]的研究結果相似.功能菌群的改變對土壤生態功能和土壤環境健康狀況的影響值得注意,但生物炭影響細菌菌群相對豐度變化的原因復雜且各不相同,還需更進一步研究. 細菌的群落結構的變化會影響生態系統的穩定和健康[31].本研究中,冗余分析圖的RDA1軸將施用生物炭處理和未施用生物炭處理明顯地分隔為兩部分,說明生物炭添加對土壤細菌群落結構有明顯的影響,這與前人的研究結果一致[28].生物炭連續施用4a對土壤的理化性質產生了顯著的影響.研究表明,生物炭能有效提高土壤pH值、總有機碳、全氮含量[27,31],與本研究結果相似.而有機碳顯著提高一方面可能是由于供試生物炭HB和JB的有機碳含量較高且HB大于JB,混入土壤后提高了土壤總有機碳含量,提高的幅度與施用量呈正相關,另一方面生物炭在施入土壤后可能改變土壤有機碳的有效性,增加土壤對有機碳的固持作用,從而有利于土壤有機碳的固定[48].土壤的基本特性又與土壤微生物生長密切關聯.Yao等[11]研究發現土壤pH值最大程度的解釋了細菌群落結構的變化.Huang等[49]研究發現土壤含水量、有機碳可以增強或削弱樹種對土壤微生物的影響,是土壤微生物群落結構組成的決定因素.但在本研究中,微生物量碳,含水率、銨態氮、有機碳對細菌群落結構的影響較大,這與劉洋等[32]、zhang等[50]的研究結果相似,只是在重要性排序上略有不同,可能因為不同土壤類型的生物組成不同,環境因子對不同類群微生物的影響程度存在差異.優勢細菌菌群與環境因子的相關性熱圖顯示,銨態氮與放線菌門、綠彎菌門這兩種主要類群存在顯著相關關系,表明銨態氮是影響細菌群落結構的關鍵因子.本試驗結果表明,生物炭施用量和種類對細菌群落結構的影響無明顯規律,環境因子對微生物群落結構的影響也各不相同,這種群落結構的差異是否會導致土壤肥力、健康狀況變化有待于進一步研究. 4.1 生物炭施加對黃土高原半干旱區4a后的土壤細菌群落有明顯影響,提高了細菌多樣性.主要表現為:ACE指數和Chao1指數隨著生物炭添加量的增大而先增后減,在3%達到最大,香農指數在施加生物炭后呈現增加趨勢,且3%JB對細菌群落的多樣性影響最佳為 6.22, JB優于HB. 4.2 生物炭添加改變了部分細菌菌群的相對豐度,在不同分類學水平上均有體現.其中,主要的優勢門為放線菌門(Actinobacteria)、變形菌門(Proteobacteria)、綠彎菌門(Chloroflexi)、酸桿菌門( Acidobacteria)和Saccharibacteria;主要的優勢綱為放線菌綱(Actinobacteria)、α-變形菌綱(Alphaproteobacteria)、酸桿菌綱(Acidobacteria);部分屬水平功能細菌的相對豐度也發生了變化,施用生物炭增加了芽孢桿菌屬()、硝化螺旋菌屬()的相對豐度,降低了土微菌屬()、根瘤菌屬()的相對豐度. 4.3 生物炭施用4a對土壤理化性質產生了顯著影響,理化性質改變進一步影響了細菌群落結構.微生物量碳、含水率、銨態氮、有機碳對細菌群落結構的影響較大,銨態氮與放線菌門、綠彎菌門呈顯著相關性,銨態氮是影響細菌群落結構的關鍵因子. [1] Reddy K R. Characteristics and applications of biochar for environmental remediation: A review [J]. Critical Reviews in Environmental Science & Technology, 2015,45(9):939-969. [2] Gunes A, Inal A, Sahi N O, et al. Variations in mineral element concentrations of poultry manure biochar obtained at different pyrolysis temperatures, and their effects on crop growth and mineral nutrition [J]. Soil Use & Management, 2016,31(4):429-437. [3] Gul S, Whalen J K, Thomas B W, et al. Physico-chemical properties and microbial responses in biochar-amended soils: mechanisms and future directions [J]. Agriculture Ecosystems & Environment, 2015, 206:46-59. [4] 高超群.生物炭在土壤環境中的應用[J]. 安徽農學通報, 2017, 23(23):55-57. Gao C Q. Application of biochar in soil environment [J]. Anhui Agricultural Science Bulletin, 2017,23(23):55-57. [5] Falkowski P G, Fenchel T, Delong E F. The microbial engines that drive Earth's biogeochemical cycles [J]. Science, 2008,320(5879): 1034-1039 [6] Zhong W H, Gu T, Wang W, et al. The effects of mineral fertilizer and organic manure on soil microbial community and diversity [J]. Plant and Soil, 2010,326(1/2):523. [7] Zheng J, Chen J, Pan G, et al. Biochar decreased microbial metabolic quotient and shifted community composition four years after a single incorporation in a slightly acid rice paddy from southwest China [J]. Science of the Total Environment, 2016,571:206-217. [8] Xu N, Tan G, Wang H, et al. Effect of biochar additions to soil on nitrogen leaching, microbial biomass and bacterial community structure [J]. European Journal of Soil Biology, 2016,74:1-8. [9] Cui E, Wu Y, Zuo Y, et al. Effect of different biochars on antibiotic resistance genes and bacterial community during chicken manure composting [J]. Bioresource Technology, 2016,203:11-17. [10] Luo L, Gu J D. Alteration of extracellular enzyme activity and microbial abundance by biochar addition: Implication for carbon sequestration in subtropical mangrove sediment [J]. Journal of Environmental Management, 2016,182:29. [11] Yao Q, Liu J, Yu Z, et al. Changes of bacterial community compositions after three years of biochar application in a black soil of northeast China [J]. Applied Soil Ecology, 2017,113:11-21. [12] Liao N, Li Q, Zhang W, et al. Effects of biochar on soil microbial community composition and activity in drip-irrigated desert soil [J]. European Journal of Soil Biology, 2016,72:27-34. [13] Chen J, Li S, Liang C, et al. Response of microbial community structure and function to short-term biochar amendment in an intensively managed bamboo (Phyllostachys praecox) plantation soil: Effect of particle size and addition rate [J]. Science of the Total Environment, 2017,574:24-33. [14] Hu L, Cao L, Zhang R. Bacterial and fungal taxon changes in soil microbial community composition induced by short-term biochar amendment in red oxidized loam soil [J]. World Journal of Microbiology & Biotechnology, 2014,30(3):1085. [15] Dai Z, Barberán A, Li Y, et al. Bacterial community composition associated with pyrogenic organic matter (biochar) varies with pyrolysis temperature and colonization environment [J]. Msphere, 2017,2(2):e00085-17. [16] Andert J, Mumme J. Impact of pyrolysis and hydrothermal biochar on gas-emitting activity of soil microorganisms and bacterial and archaeal community composition [J]. Applied Soil Ecology, 2015, 96:225-239. [17] Lehmann J, Rillig M C, Thies J, et al. Biochar effects on soil biota – A review [J]. Soil Biology & Biochemistry, 2011,43(9):1812-1836. [18] Wang T T, Stewart C E, Sun C C, et al.Effects of biochar addition on evaporation in the five typical Loess Plateau soils [J]. CATENA, 2018,162:29-39. [19] 郭艷亮,王丹丹,鄭紀勇,等.生物炭添加對半干旱地區土壤溫室氣體排放的影響[J]. 環境科學, 2015,36(9):3393-3400.Guo Y L, Wang D D, Zheng J Y, et al. Effect of biochar on soil greenhouse gas emissions in semi-arid region [J]. Environmental Science, 2015,36(9):3393-3400. [20] 鮑士旦.土壤農化分析.3版[M]. 中國農業出版社, 2000. Bao Shidan. Soil agro-chemistrical analysis 3 [M]. China Agriculture Press, 2000. [21] Vance E D, Brookes P C, Jenkinson D S. An extraction method for measuring soil microbial biomass C [J]. Soil Biology & Biochemistry, 1987,19(6):703-707. [22] Mcbain A J, Bartolo R G, Catrenich C E, et al. Microbial characterization of biofilms in domestic drains and the establishment of stable biofilm microcosms [J]. Applied & Environmental Microbiology, 2003,69(1):177-85. [23] 楊亞東,王志敏,曾昭海.長期施肥和灌溉對土壤細菌數量、多樣性和群落結構的影響[J]. 中國農業科學, 2018,51(2):290-301.Yang Y D, Wang Z M, Zeng Z H.Effects of long-Term different fertilization and irrigation managements on soil bacterial abundance, diversity and composition [J].Scientia Agricultura Sinica, 2018,51(2): 290-301. [24] 王亞男,曾希柏,王玉忠,等.設施蔬菜種植年限對氮素循環微生物群落結構和豐度的影響[J]. 應用生態學報, 2014,25(4):1115-1124.Wang Y N, Zeng X B, Wang Y Z, et al. Effects of vegetable cultivation years on microbial biodiversity and abundance of nitrogen cycling in greenhouse soils [J].Chinese Journal of Applied Ecology, 2014,25(4): 1115-1124. [25] 劉根林.轉基因大豆對根際土壤微生物群落的影響及其多樣性指數的度量[D]. 南京:南京大學, 2012.Liu G L. Effects of transgenic soybeans on the rhizospheric soil microbial communities and their diversity indices measurement [D]. Nanjing: Nanjing University, 2012. [26] Doan T T, Bouvier C, Bettarel Y, et al. Influence of buffalo manure, compost, vermicompost and biochar amendments on bacterial and viral communities in soil and adjacent aquatic systems [J]. Applied Soil Ecology, 2014,73(2):78-86. [27] Nguyen T T N, Wallace H M, Xu C Y, et al. The effects of short term, long term and reapplication of biochar on soil bacteria [J]. Science of the Total Environment, 2018,636:142-151. [28] 烏英嗄,張貴龍,賴 欣,等.生物炭施用對華北潮土土壤細菌多樣性的影響[J]. 農業環境科學學報, 2014,33(5):965-971.Wu Y G, Zhang G L, Lai X, et al. Effects of biochar applications on bacterial diversity in fluvor-aquic soil of north china [J]. Journal of Agro-Environment Science, 2014,33(5):965-971. [29] 張玉潔,吳 婷,趙 娟,等.生物炭添加對秸稈還田土壤細菌群落結構和多樣性影響[J]. 環境科學學報, 2017,37(2):712-720.Zhang Y J, Wu T, Zhao J et al. Effect of biochar amendment on bacterial community structure and diversity in straw-amended soils [J]. Acta Scientiae Circumstantiae, 2017,37(2):712-720. [30] 李 力,劉 婭,陸宇超,等.生物炭的環境效應及其應用的研究進展[J]. 環境化學, 2011,30(8):1411-1421.Li L, Liu Y, Lu Y C, et al. Review on environmental effects and applications of biochar [J]. Environmental Chemistry, 2011,30(8): 1411-1421. [31] 陳 坤,徐曉楠,彭 靖,等.生物炭及炭基肥對土壤微生物群落結構的影響[J]. 中國農業科學, 2018,51(10):1920-1930.Chen K, Xu X N, Peng J, et al. Effects of biochar and biochar-based fertilizer on soil microbial community structure [J]. Scientia Agricultura Sinica, 2018,51(10):1920-1930. [32] 劉 洋,黃懿梅,曾全超.黃土高原不同植被類型下土壤細菌群落特征研究[J]. 環境科學, 2016,37(10):3931-3938.Liu Y, Huang Y M, Zeng Q C. Soil bacterial communities under different vegetation types in the loess plateau [J]. Environmental Science, 2016,37(10):3931-3938. [33] Lauber C L, Hamady M, Knight R, et al. Pyrosequencing-based assessment of soil pH as a predictor of soil bacterial community structure at the continental scale [J]. Applied & Environmental Microbiology, 2009,75(15):5111. [34] Zeng J, Liu X, Song L, et al. Nitrogen fertilization directly affects soil bacterial diversity and indirectly affects bacterial community composition [J]. Soil Biology & Biochemistry, 2016,92:41-49. [35] 張 星,張晴雯,劉杏認,等. 施用生物炭對農田土壤氮素轉化關鍵過程的影響[J]. 中國農業氣象, 2015,36(6):709-716.Zhang X, Zhang Q W, Liu X R, et al. Effects of biochar on the key soil nitrogen transformation processes in agricultural soil [J]. Chinese Journal of Agrometeorology, 2015,36(6):709-716. [36] Bergmann M J, Driscoll D C, Emerson D T. Indium gallium nitride-based Ohmic contact layers for gallium nitride-based devices: US, US7943924 [P]. 2011. [37] 韓東東,郝振宇,高廣海,等.寡營養細菌及其生態作用和應用的研究進展[J]. 微生物學通報, 2012,39(4):526-535.Han D D, Hao Z Y, Gao H G, et al. Ecological function of oligotrophic bacteria and their Applications in the environment [J]. Microbiology China, 2012,39(4):526-535. [38] Wu C, Shi L Z, Xue S G, et al. Effect of sulfur-iron modi?ed biochar on the available cadmium andbacterial community structure in contaminated soils [J]. Science of the Total Environment, 2019,647: 1158–1168. [39] Zhang T, Shao M F, Ye L. 454pyrosequencing reveals bacterial diversity of activated sludge from 14sewage treatment plants [J]. Isme Journal, 2012,6(6):1137-1147. [40] Campbell B J, Polson S W, Hanson T E, et al. The effect of nutrient deposition on bacterial communities in Arctic tundra soil [J]. Environmental Microbiology, 2010,12(7):1842. [41] Zaidi A, Khan M S, Ahemad M, et al. Plant growth promotion by phosphate solubilizing bacteria [J]. Acta Microbiologica Et Immunologica Hungarica, 2009,56(3):263. [42] Chen Z Y, Zhang J. Research progress on antimicrobial mechanism and genetic engineering of Bacillus for plant diseases biocontrol [J]. Acta Phytopathologica Sinica, 2003. [43] 張華勇,李振高.土壤芽孢桿菌及其資源的持續利用[J]. 土壤, 2001,33(2):92-97. Zhang H Y, Li Z G. Sustainable use of Bacillus and its resources in soil [J]. Soil, 2001,33(2):92-97. [44] Zahran H H. Rhizobium-Legume symbiosis and nitrogen fixation under severe conditions and in an arid climate [J]. Microbiology & Molecular Biology Reviews, 1999,63(4):968-989. [45] 聶 剛,陳衛民,韋革宏.神木地區耐旱灌木和草本豆科植物根瘤菌遺傳多樣性[J]. 應用生態學報, 2014,25(6):1674-1680. Nie G, Chen W M, Wei G H. Genetic diversity of rhizobia isolated from shrubby and herbaceous legumes in Shenmu arid area, Shaanxi, China [J]. Chinese Journal of Applied Ecology, 2014,25(6):1674- 1680. [46] 徐慧敏,閆 海,馬 松,等.鞘氨醇單胞菌USTB-05對微囊藻毒素的生物降解[J]. 中國環境科學, 2014,34(5):1316-1321.Xu H M, Yan H, Ma S, et al. Biodegradation of microcystins by Sphingopyxis sp. USTB-05 [J].China Environmental Science, 2014, 34(5):1316-1321. [47] 陳澤斌,高 熹,王定斌,等.生物炭不同施用量對煙草根際土壤微生物多樣性的影響[J]. 華北農學報, 2018,33(1):224-232.Chen Z B, Gao X, Wang D B, et al. Effects of different biochar application rates on rhizosphere soil microbial diversity of tobacco [J].Acta Agriculturae Boreali-Sinica, 2018,33(1):224-232. [48] 羅 梅,田 冬,高 明,等.紫色土壤有機碳活性組分對生物炭施用量的響應[J]. 環境科學, 2018,(9):4327-4337.Luo M, Tian D, Gao M, et al. Soil organic carbon of purple soil as affected by different application of biochar [J]. Environmental Science, 2018,(9):4327-4337. [49] Huang Z, Wan X, He Z, et al. Soil microbial biomass, community composition and soil nitrogen cycling in relation to tree species in subtropical China [J]. Soil Biology & Biochemistry, 2013,62(5):68- 75. [50] Zhang X F, Zhao L, Xu S J, et al. Soil moisture effect on bacterial and fungal community in Beilu River (Tibetan Plateau) permafrost soils with different vegetation types [J]. Journal of Applied Microbiology, 2013,114(4):1054-1065. 致謝:本研究的現場采樣工作由馬江波碩士和譚連帥碩士協助完成,在此表示感謝. Effects of biochar addition on soil bacterial community in semi-arid region. WANG Ying1, SUN Ceng-ceng1, ZHOU Ji-hai2, WANG Tong-tong1, ZHENG Ji-yong1,3* (1.College of Natural Environment and Resources, Northwest A&F University, Yangling 712100, China;2.Nanchang Institute of Technology, Nanchang 330099, China;3.State Key Laboratory of Soil Erosion and Dryland Farming on the Loess Plateau, Northwest A&F University, Yangling 712100, China)., 2019,39(5):2170~2179 Using surface soils collected from Guyuan Ecological Experiment Station in Semi-arid Region restored by biochar for 4years as research objects, th effects of two types of biochar (locust tree bark biochar、sawdust biochar) at three different mass percentages (1%、3% and 5%) on soil bacterial community diversities and structures were investigated by using Miseq high-throughput sequencing technology. The results showed that the diversity of soil bacterial community was increased with biochar addition, and the diversities were higher in the treatment with sawdust biochar than that in the treatment with locust tree bark biochar. Further, 3% sawdust biochar addition had the best effect on bacterial community diversities, and its Shannon index was 6.22. The dominant taxa at phyla level were Actinobacteria, Proteobacteria, Chloroflexi, Acidobacteria and Saccharibacteria,with relative abundance of 76.80%~85.31%. The dominant taxa at class level were Actinobacteria, Alphaproteobacteria, Acidobacteria, with relative abundance of 48.13%~57.08%. At genus level, the relative abundance of the,increased, while the relative abundance of the,decreased after biochar treatmetns. Hierarchical clustering and redundancy analysis (RDA) indicated that the application of biochar greatly affected the bacteria community structures. The microbial biomass carbon, water content, ammonium nitrogen (NH4+-N), organic carbon contents had great influences on the variations of bacterial community structure. The heatmap of the correlation between taxa of dominant bacteriaand environmental factors further indicated that NH4+-N concentration was significantly correlated with Actinobacteria and Chloroflexi. Therefore, the change in NH4+-N concentration caused by biochar addition was the key factor leading to the variations of soil bacterial communities. biochar;soil bacteria;diversity;community structure;high-throughput sequencing X172 A 1000-6923(2019)05-2170-10 王 穎(1994-),女,寧夏石嘴山人,西北農林科技大學碩士研究生,研究方向為土壤污染與修復.發表論文2篇. 2018-10-23 國家自然科學基金資助項目(41571225,41661065);“十三五”國家重點研發計劃項目(2016YFC0501702,2017YFC0504504);中國科學院STS項目(KFJ-STS-ZDTP-012) *責任作者, 副研究員, zhjy@ms.iswc.ac.cn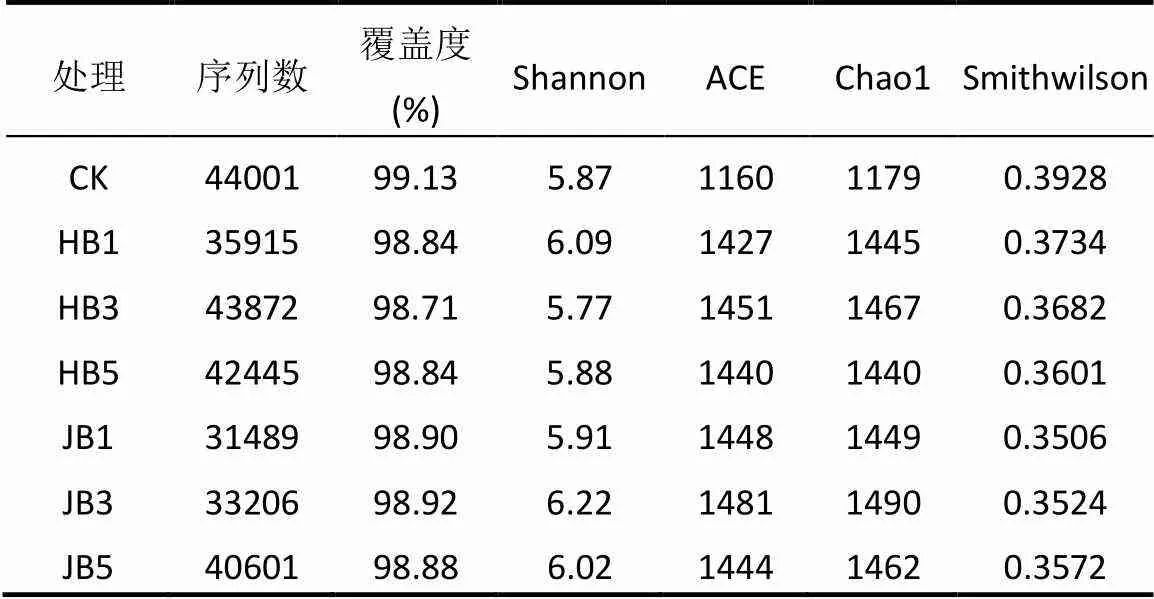
2.3 細菌群落組成

2.4 不同生物炭處理的細菌群落結構及其與環境因子的關系


3 討論
4 結論
猜你喜歡
天天愛科學(2022年9期)2022-09-15 01:12:54
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
科學大眾(2021年9期)2021-07-16 07:02:54
當代陜西(2021年2期)2021-03-29 07:41:24
軍事文摘(2020年20期)2020-11-28 11:42:50
航空世界(2020年10期)2020-01-19 14:36:20
媽媽寶寶(2017年3期)2017-02-21 01:22:28
中國塑料(2016年3期)2016-06-15 20:30:00
