大米淀粉結構與特性研究進展
2019-06-08 02:12:58劉傳菊李歡歡湯尚文聶榮祖豁銀強
中國糧油學報 2019年12期
關鍵詞:結構
劉傳菊 李歡歡 湯尚文 聶榮祖 豁銀強
(湖北文理學院,襄陽 441053)
淀粉是大米的主要組分,占其干物質的80%以上。大米淀粉具有易消化、清淡無味、顆粒小、白色、淀粉糊凍融穩定性好、抗酸解及支鏈/直鏈淀粉比例差異大、消費者易接受等特征。這些特性使大米淀粉在食品和藥品領域具有廣泛的應用。淀粉的特性取決于淀粉的粒徑大小分布、直鏈/支鏈淀粉比、淀粉顆粒形態及分子結構等結構特征,淀粉中殘留的蛋白質等成分對淀粉特性也有重要的影響,而淀粉的特性又是影響淀粉開發與應用的主要因素。
1 大米淀粉的制備
制備純化大米淀粉的關鍵是去除大米蛋白,目前工業生產中主要利用堿法和機械法,一些科研機構開展了酶法制備大米淀粉的研究。
1.1 堿法
堿法是工業上制備大米淀粉的傳統方法。80%左右的大米蛋白是堿溶性谷蛋白,在堿法工藝中,常用0.3%~0.5%的氫氧化鈉溶液浸泡大米12~24h,浸泡溫度從室溫至50℃不等。經濕磨粉碎浸泡軟化的大米,釋放出淀粉形成淀粉乳,隨后將淀粉乳保持懸浮狀態10~24h以促進蛋白質溶解,過濾去除細胞壁物質,淀粉乳再經水洗、中和及干燥得相應的大米淀粉產品[1-2]。該法具有淀粉回收率高、蛋白殘留低等特點(通常低于1%)[3,4]。副產物蛋白質的純度較高,但由于特殊的氣味及產生有毒的賴丙氨酸[5-7],而不適于用作食品原料。
1.2 酶法
堿法制備大米淀粉會產生大量的堿性廢液及高濃度鹽的廢水,一些學者研究了蛋白酶消化處理制備大米淀粉的效果。蛋白酶在pH6.5條件下50℃處理18h,所得淀粉含量與堿法制備的相當或更高,酶法降低了破損淀粉量,產品的特性與堿法制備的相當[8]。食品級堿性蛋白酶和中性蛋白酶去除蛋白的效果比其他蛋白酶的效果更好,當米粉用纖維素酶進行預處理后,蛋白的去除率更高[9]。在中性條件下米粉懸液經纖維素酶預處理以降解纖維素組織,隨后用蛋白酶水解緊密結合在淀粉顆粒表面的蛋白質,所制備大米淀粉的得率是76.5%,蛋白含量比堿法制備淀粉高,通過堿液或SDS溶液洗滌能使蛋白含量降低到堿法制備淀粉的水平[10]。中性蛋白酶聯合高強度超聲制備淀粉的得率為79.8%~86.7%,蛋白殘留量0.50%~0.96%[11]。利用碎米為原料,凍融浸漬-酶法制備大米淀粉的得率達69.31%,所得淀粉得率比酶法及堿法高[12]。酶法主要還處于實驗室研究階段,目前還沒能應用于工業生產。
1.3 機械法
機械法制備大米淀粉是一種濕磨技術,通過機械方法將胚乳中的蛋白質與淀粉團粒解離,根據密度間差異,通過物理方法將兩者分離。經該法所得的淀粉團粒位于團塊或直徑10~20μm的聚集體中,所得淀粉通常殘留0.25%~7%的蛋白質。通過該法制備的淀粉外觀上與傳統堿法制備的相似,但兩者的糊化等功能特性不同。與堿法不同,機械法產生的米蛋白質是一種價值較高的副產品,其具有良好的風味且不會產生有毒的副產物,在食品工業中具有廣泛的應用范圍。采用該法從糙米中生產的淀粉含有一定量的蛋白質和米糠油,這些成分賦予米淀粉獨特的物化特性,該類淀粉已作為原料應用于食品工業[2]。
2 淀粉的組成
淀粉的主要組分是直鏈淀粉和支鏈淀粉。支鏈淀粉是米淀粉的主要組分,組成淀粉顆粒的65%~85%,不過蠟質突變系的支鏈淀粉含量可以達到100%。直鏈淀粉通常形成螺旋結構,其能與碘等非極性物質形成復合物。淀粉樣品表觀直鏈淀粉含量通常就是根據淀粉-碘形成的藍色復合物進行分析測試[13]。由于支鏈淀粉長分支能與碘形成有色復合物,表觀直鏈淀粉含量并不能完全代表真實的直鏈淀粉含量。大米表觀直鏈淀粉含量范圍是0.6%~27.8%[14],根據表觀直鏈淀粉含量常將大米分為蠟質(0%~2%)、非常低(2%~12%)、低(12%~20%)、中等(20%~25%)及高(高于30%)直鏈淀粉含量五種類型[15]。AE(Amylose extender,直鏈淀粉擴增)突變體的表觀直鏈淀粉含量達35%~40%,目前還沒有稻米品種的表觀直連淀粉含量同高直鏈玉米淀粉那樣達到70%。不同水稻亞系的表觀直連淀粉含量不同,秈稻通常高于粳稻。秈稻包含四類直鏈淀粉含量類型,而普通的粳稻僅有低至中等表觀直鏈淀粉含量類型。粳稻和秈稻均包含蠟質品種。表觀直連淀粉含量也受生長地、氣候及土壤條件等環境因素的影響[3]。
大米胚乳中的蛋白質緊密的結合在淀粉顆粒的表面或者淀粉顆粒孔道腔而很難去除。不同方法制備大米淀粉殘留的蛋白質含量不同,堿法制備大米淀粉的蛋白質含量通常低于1%,機械法和酶法制備淀粉的蛋白質含量稍高[2]。淀粉中殘留的蛋白質主要是一些淀粉顆粒結合蛋白,這些蛋白大部分是參與淀粉合成的酶,通常認為這些蛋白位于淀粉孔腔內而難以去除[16]。
除了蛋白質之外,大米淀粉中含有少量的脂質、磷及礦物質[17]。非蠟質大米淀粉含0.3%~0.4%的結合脂質,而蠟質大米的脂質較低。大米淀粉中的脂質大部分是游離脂肪酸和溶血磷脂,內部脂質與直鏈淀粉含量呈顯著的正相關。非蠟質大米淀粉中的脂質由32%的游離脂肪酸和68%的溶血磷脂組成。直鏈淀粉能夠與脂質形成包合物,直鏈淀粉和內源性或外源添加的脂質在淀粉發生凝膠化時發生包合,天然淀粉顆粒內也存在直鏈淀粉-脂質復合體。
淀粉中存在相對較少的礦物質。淀粉中的磷對其加工特性有重要影響,其影響淀粉的黏度、糊透明度及穩定性等。淀粉中的磷主要以磷酸單脂和磷脂2種形式存在,非蠟質大米淀粉中主要是磷脂[18],蠟質大米淀粉中主要是磷酸單脂選擇性與直鏈淀粉結合。淀粉磷酸單脂是淀粉磷酸化的產物,主要存在支鏈淀粉內,而直鏈淀粉中的含量非常低,大約80%~90%磷酸是與葡萄糖殘基的C6結合[18]。
3 淀粉的結構
直鏈淀粉和支鏈淀粉以不同尺度形成了淀粉顆粒的進而形成多級結構體系(見圖1),主要包括顆粒結構、生長環結構、Blocklets小體、層狀結構、結晶結構以及鏈結構。
3.1 顆粒結構
淀粉以顆粒形式儲存在植物體內,不同植物來源淀粉的大小和形狀不同。淀粉顆粒直徑從不足1~100μm不等,形態有多面形、橢圓形、圓形、球形或不規則形等。大米單個淀粉顆粒直徑大約3~8μm,在谷類中最小,呈多面形或不規則形,淀粉顆粒通常聚集為復合淀粉顆粒[20]。大米淀粉復合顆粒直徑達150μm以上,通常為多面體,每個淀粉復合顆粒含20~60個淀粉顆粒[21]。如圖2所示,大米淀粉顆粒表面呈現較多的微孔結構,玉米和高粱等谷物淀粉表面也觀察到類似的微孔結構,而在木薯、馬鈴薯等根莖、塊莖類淀粉中并未觀察到此類微孔[22,23]。微孔結構是淀粉顆粒表面至內部臍點空腔的通道,為淀粉酶等作用于淀粉分子提供了作用位點,從而使大米等谷物淀粉顆粒較根莖、塊莖類淀粉顆粒更易被酶解。

注:a 淀粉顆粒,b 無定形和半結晶生長環,c無定形和結晶層,d Blocklets小體,e 支鏈淀粉雙螺旋結構,f 淀粉納米晶,g 支鏈淀粉,h 直鏈淀粉。

圖2 大米淀粉顆粒結構
3.2 生長環結構
采用高倍光學顯微鏡可以比較清晰地觀察到淀粉顆粒由半結晶和無定形交替形成的生長環結構[24],見圖3。生長環以淀粉顆粒臍點為同心環向外生長,其數目及大小與淀粉的生物來源和成熟程度有關,如蠟質玉米淀粉的生長環結構約為100~400nm,而大米淀粉的生長環大小接近400nm[25]。
3.3 Blocklets小體與片層結構
掃描電鏡、透射電鏡、原子力顯微鏡及酶解結果顯示,結晶層和半結晶層組裝為重復的結晶與無定形片層交替的 Blocklets小體結構,模式圖見圖4。結晶片層主要由支鏈淀粉側鏈簇組裝成晶格,而支鏈淀粉分支點、直鏈淀粉及無定形態直鏈淀粉分子組成了淀粉止水塞的無定形片層。依據淀粉的來源和在淀粉顆粒內的位置不同,Blocklets小體結構的大小和形狀不同,但其近似為直徑20~500nm的不透明球狀體,含較少的結晶結構。大米淀粉的Blocklets小體結構平均大小是100nm,由大約280個支鏈淀粉側鏈簇構成[27]。
3.4 結晶結構
淀粉顆粒是由結晶區和無定形區組成的半結晶體。支鏈淀粉的外鏈和直鏈淀粉能形成雙螺旋結構,排列成有序的結晶區,結晶區主要由支鏈淀粉的側鏈組成。富含支鏈淀粉的淀粉中,結晶區是由支鏈淀粉分子外鏈通過氫鍵形成的雙螺旋結構。直鏈淀粉對普通和蠟質大米淀粉結晶度影響較小,但其對高直鏈淀粉含量淀粉的結晶度有重要影響。淀粉顆粒在偏光顯微鏡下呈雙折射現象,表現出典型的馬爾文十字,淀粉顆粒的雙折射顯示微晶主軸呈徑向排列。常用X射線衍射分析淀粉的結晶度及淀粉的晶體類型。大米淀粉呈典型的A晶型,蠟質大米淀粉的結晶度比普通大米淀粉高。Sugary(糖質)突變體淀粉也呈A晶型,AE突變體與馬鈴薯和玉米淀粉類似[29]。大米淀粉XRD的最強反射角出現在15°、17°、18°和23°,結晶度在29.2%~39.3%[30]。表觀直鏈淀粉含量高的淀粉的結晶度32.3%,而凝膠化溫度高的蠟質大米淀粉的結晶度為45.5%。2種Sugary突變體淀粉的結晶度分別是22.9%和18.3%。淀粉結晶度可能受直鏈淀粉含量和直鏈淀粉結構的影響,表觀直鏈淀粉含量與結晶度呈負相關[31]。
3.5 淀粉分子結構
淀粉是一類具有分支的葡聚糖,主要包括直鏈淀粉和支鏈淀粉兩大類。直鏈淀粉是葡萄糖殘基通過α-1,4糖苷鍵連接的線性聚合物,部分直鏈淀粉含有少量的長分支(圖5)。不同大米直鏈淀粉的分子質量不同,平均分子質量是3.73×105~4.89×106g/mol[32],數均聚合度(DPn)和平均鏈長分別是847~1118和230~370[1]。6種水稻的直鏈淀粉經脫支酶水解后的數均分子量和重均分子質量是5.1~6.9×105和1.4~1.8×105g/mol,重均聚合度(DPw)和數均聚合度(DPn)分別是2569~4273和847~1118[34]。
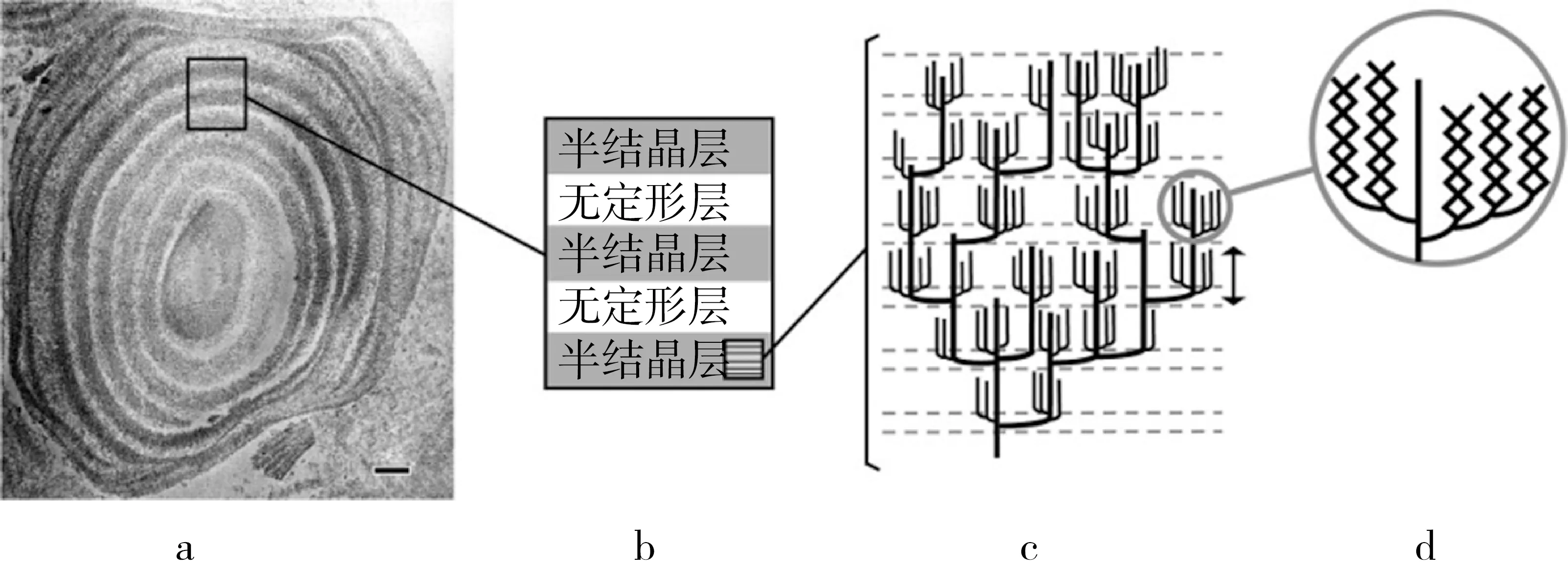
注: a 生長環結構,b 交替排列的結晶片層和無定形片層,c 支鏈淀粉簇狀結構,d 螺旋結構。

圖4 淀粉Blocklets小體與片層結構示意圖[28]
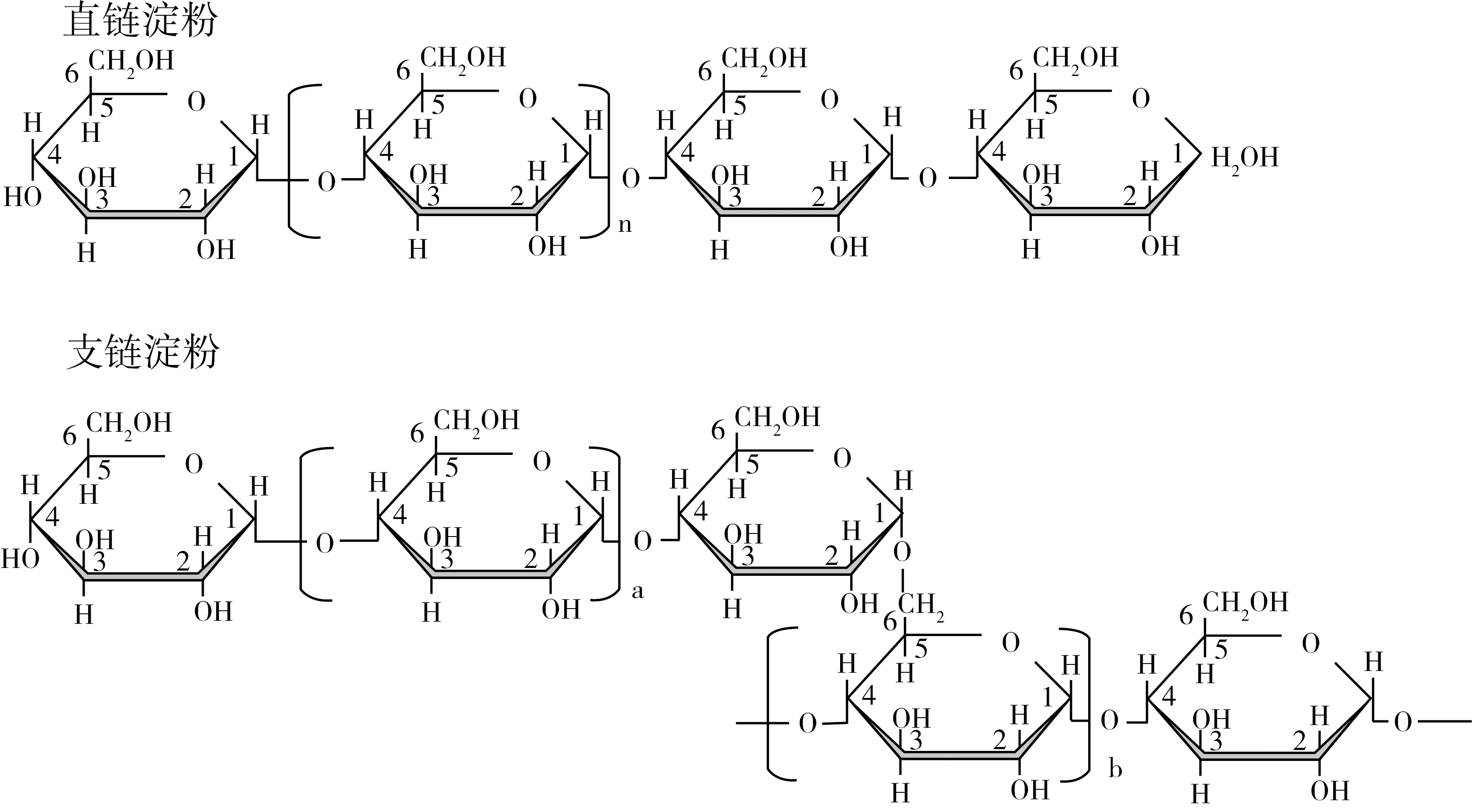
圖5 直鏈淀粉及支鏈淀粉的結構[33]
支鏈淀粉具有較多的分支結構,主要由α-1,4糖苷鍵鏈接,且具有較多以α-1,6糖苷鍵連接的短分支(圖5)。不同植物來源支鏈淀粉的數均分子量是7.0×107~5.7×109g/mol,蠟質和非蠟質淀粉的數均分子量分別為2.7×109和5.7×109g/mol。秈米、粳米和蠟質大米支鏈淀粉的數均聚合度范圍是8200~12900[21]。支鏈淀粉的分支可分為A、B和C鏈。外層的A鏈最短(DP6~12),其通過α-1,6糖苷鍵鏈接到B鏈上,B鏈上鏈接1個或多個A鏈或B鏈,依跨越簇的數量及長度不同,B鏈可進一步分為B1、B2、B3和B4(跨越1~4個簇),典型的B1和B2鏈長分別為DP13~24和DP25~36,B3和B4鏈長為DP>37,每個支鏈淀粉的單個C鏈含唯一的還原性末端[21,30]。
4 大米淀粉的物化特征
4.1 凝膠化特征
凝膠化是淀粉的重要功能特性之一,其受淀粉組成、支鏈淀粉分子結構、顆粒結構、顆粒形貌及顆粒大小分布等因素的影響[34]。通常采用DSC分析淀粉的凝膠化起始溫度(To)、峰值溫度(Tp)、終止溫度(Tc)及焓值(ΔH)等凝膠化特征參數,ΔH主要反映淀粉顆粒中分子的有序結構[35],而凝膠化溫度與結晶完整性有關[36]。大米淀粉的凝膠化特征受稻米品種的影響,非蠟質大米淀粉的ΔH、To、Tp及Tc分別是3.7~19.2J/g、53.3~75.9℃、61.8~80.0℃和70.9~85.4℃;蠟質大米淀粉的相應值是3.4~16.7J/g、5.5~63.0℃、1.6~75.8℃和4.9~88.3℃;長粒大米淀粉的相應值為8.7~21.6J/g、50.9~77.0℃、62.3~80.8℃和72.1~89.6℃[34]。蠟質大米淀粉的Tp比普通大米的低,直鏈淀粉含量對蠟質大米淀粉的凝膠化溫度沒有影響[37],有關直鏈淀粉含量對大米淀粉ΔH影響的報道不一[21]。較多的支鏈淀粉的長鏈對凝膠化產生延遲作用,支鏈淀粉的短支鏈促進淀粉凝膠化[37]。
4.2 糊化特征
在過量水分情況下連續加熱并不斷攪拌,淀粉顆粒吸水膨脹、隨后由于淀粉結構瓦解引起淀粉顆粒破裂。直鏈淀粉滲出及淀粉顆粒破裂形成黏性糊。淀粉的糊化特性對其應用有重要影響。常用布拉班德黏度儀和快速黏度儀(RVA)等分析淀粉的糊化特性。RVA通常測定淀粉的糊化溫度(PT)、峰值黏度(PV)、最終黏度(FV)、崩解值(BD)、熱糊黏度(HPV)、冷糊黏度(CPV)、回落值(SB)等特征參數。大米淀粉的糊化特性主要受Wx(蠟質)基因的控制,因此大米淀粉的糊化特征參數與淀粉的表觀直鏈淀粉含量有關,支鏈淀粉的分子結構也對淀粉的糊化特征有一定的影響[29]。支鏈淀粉的長分支和短分支分別與崩解值呈正相關和負相關[38]。大米淀粉的糊化特性也受氣候溫度、大氣二氧化碳含量、光照、灌溉用水量及土壤營養的影響。CPV、SB及BD主要受大米基因型的影響,而PV和HPV主要受環境因素的影響[39]。
4.3 回生特征
淀粉回生是加熱的淀粉糊在溫度降低到晶體熔點以下時,膨脹淀粉顆粒內的直鏈淀粉和支鏈淀粉分子重新聚集、結合成有序的結構,導致黏度增加、凝膠變硬、質構陳化。淀粉回生包括直鏈淀粉的快速再結晶和支鏈淀粉的慢速結晶。直鏈淀粉回生引起淀粉儲存早期的變化,支鏈淀粉回生需要幾周甚至幾個月的時間,造成長期儲存淀粉流變和結構的變化。
通常采用DSC分析淀粉的回生特征。蠟質大米淀粉長分支的量與回生焓及凝膠硬度呈正相關,短鏈/長鏈比值幾乎對淀粉所有的回生參數均有影響[40,41]。非蠟質稻米的表觀直鏈淀粉含量與回生焓及回生百分比呈正相關[42]。蠟質大米淀粉回生特征主要受SSIIa(可溶性淀粉合成酶)基因影響[43],非蠟質大米淀粉回生特征主要受Wx及SSIIa和ISAI(異淀粉酶)基因的調節[42]。儲藏淀粉凝膠中支鏈淀粉回生形成的有序性比天然淀粉低,不同品種稻米淀粉的回生焓值是2.16~3.71J/g[44]。
4.4 消化特性
根據在體外的消化速率不同,通常將淀粉分為快速消化淀粉(RDS)、慢速消化淀粉(SDS)和抗性淀粉(RS)。大米淀粉中RDS的質量分數為78.5%~87.5%,蠟質大米淀粉RDS比普通大米淀粉高,大米淀粉中SDS和RS質量分數分別為1.2%~6.0%和10.1%~18.0%[45]。由于采用的分析方法不同,不同研究所RDS、SDS及RS含量會有所不同。
直鏈淀粉含量、淀粉分子精細結構、殘留的蛋白及脂質均對大米淀粉的消化特性產生一定的影響。有關直鏈淀粉含量與淀粉消化率之間關系的報道結果并不一致,Kong等[31]報道RS含量隨大米淀粉中直鏈淀粉含量增加而增加,而RDS含量隨直鏈淀粉含量增加而降低,淀粉酶首先對不定形區的直鏈淀粉產生水解,水解物又重新結合而抗性增加,但Van Hung等[46]報道天然蠟質大米淀粉的RS含量比高直鏈淀粉含量大米淀粉和普通大米淀粉高。根據消化速率,有研究報道直鏈淀粉含量與SDS含量[47]和消化速率[48]呈負相關,Cai 等[48]認為直鏈淀粉與碘形成的復合體不容易水化,進而不利于酶浸入淀粉顆粒內部,不過也有研究認為直鏈淀粉含量與RDS或消化動力學參數無關[49]。由于大多數研究都是比較不同基因型大米淀粉,因而導致大米淀粉直鏈淀粉含量與消化性之間的關系比較復雜。Lin等[50]研究了相同遺傳背景但直鏈淀粉含量不同的大米淀粉,發現支鏈淀粉結構相同而直鏈淀粉含量與消化率間存在顯著的負相關。因此,大米淀粉消化率主要受直鏈淀粉含量的影響,同時支鏈淀粉精細結構對其也有一定的貢獻。
此外,由于降低了淀粉酶與淀粉的作用,殘留在淀粉顆粒表面的蛋白質和脂質會降低淀粉的消化率[51]。
5 結論
不同方法制備大米淀粉中殘留的蛋白質和脂質等成分的含量不同,這些殘留成分會一定程度上影響淀粉的物化特征。不過,淀粉的物化特征主要由淀粉的顆粒結構、生長環結構、層狀結構、結晶度、支鏈/直鏈淀粉比例及淀粉分子精細結構等不同層級結構所決定。隨著對大米淀粉多尺度結構理解的加深,人們可以更有針對性地對淀粉結構進行調控,利用物理、酶法及化學方法修飾淀粉結構而改善大米淀粉的物化特性及擴大其應用范圍。
猜你喜歡
小獼猴智力畫刊(2023年4期)2023-04-23 08:49:58
哲學評論(2021年2期)2021-08-22 01:53:34
中華詩詞(2019年7期)2019-11-25 01:43:04
模具制造(2019年3期)2019-06-06 02:10:54
中學生數理化·高一版(2018年1期)2018-02-10 05:20:03
影視與戲劇評論(2016年0期)2016-11-23 05:26:01
七彩語文·寫字與書法(2016年7期)2016-07-28 21:40:22
七彩語文·寫字與書法(2016年6期)2016-07-15 19:36:34
人間(2015年21期)2015-03-11 15:23:21
現代企業(2015年9期)2015-02-28 18:56:50
