海洋酸化與重金屬、有機污染物和人工納米顆粒的聯合毒性效應研究進展
2019-06-10 02:19:06王昊趙信國陳碧鵑夏斌孫雪梅朱琳曲克明
生態毒理學報 2019年1期
王昊,趙信國,陳碧鵑,夏斌,*,孫雪梅,朱琳,曲克明
1. 上海海洋大學海洋科學學院,上海 201306 2. 中國水產科學研究院黃海水產研究所 農業部海洋漁業資源可持續發展重點實驗室 山東省漁業資源與生態環境重點實驗室,青島 266071 3. 青島海洋科學與技術國家實驗室海洋生態與環境科學功能實驗室,青島 266237
自18世紀工業革命以來,在化石燃料燃燒等人類活動的影響下,大氣CO2濃度不斷增加,目前已達到409.65 ppm,增長了約45%。由于大氣和海洋之間的氣體交換作用,人類活動排放的CO2中約有1/3被海洋吸收,從而造成海水酸度的增大和碳酸鹽平衡體系的變化,即“海洋酸化”現象(Ocean Acidification)[1-2]。當前表層海水的pH值為8.1~8.2,根據政府間氣候變化專門委員會(Intergovernmental Panel on Climate Change, IPCC)的預測,到2100年,表層海水pH值將下降至7.8~7.7;到2300年,表層海水pH值將下降至7.4~7.3[1,3-4]。海洋酸化勢必會引起海洋環境的變化,直接影響到海洋生物賴以生存的環境條件,嚴重威脅海洋生態系統的安全與穩定[5]。
隨著全球經濟的高速發展,大量的污染物(重金屬、有機污染物和人工納米材料等)不斷進入到海洋環境中。重金屬作為一類海洋環境中常見的污染物,在沿海、河口以及沉積物中高濃度富集。Cu、Pb等是許多沿海與河口的水體和沉積物中最常見的重金屬污染物[6],可以在海洋生物的脂肪組織中發生累積,并通過食物鏈的營養傳遞對人類健康產生潛在的危害[7]。溢油事故的頻繁爆發及農藥化肥的不合理使用,使得大量有機污染物最終匯入海洋,對海洋生態系統造成嚴重威脅[8]。進入21世紀以來,隨著納米技術的迅猛發展,納米材料在工業、能源、醫療和消費品中得到了廣泛的應用[9],不可避免地導致大量人工納米顆粒(engineered nanoparticles, NPs)通過地表徑流、意外泄露等方式進入到海洋環境中,從而對海洋環境造成潛在的風險[10]。已經有越來越多的研究表明NPs會對浮游植物、浮游動物、游泳動物、底棲生物等產生毒性效應[11]。
海洋酸化與環境污染物(重金屬、有機污染物和NPs等)已成為全球海洋生態系統面臨的重大環境問題,它們同時存在,共同影響著海洋生態系統(圖1)。研究表明海洋酸化會改變污染物的環境行為,如改變重金屬離子的存在價態[12],抑制有機污染物的降解[13],改變人工納米顆粒的懸浮性和離子釋放能力[14],從而影響污染物對海洋生物的毒性效應(圖2)。本文回顧了海洋酸化的生態效應以及重金屬、有機污染物和NPs對海洋生物的毒性效應,重點分析了海洋酸化對污染物環境行為的影響及產生的聯合毒性效應,對近年來的相關報道進行了總結歸納(表1),指出了當前研究的特點和不足,并對未來的研究進行了展望。
1 海洋酸化對海洋生物的影響(The impact of ocean acidification on marine life)
海洋酸化引起的海水化學參數變化對從自養生物到頂級捕食者的所有營養水平的生物都有影響,主要有:(1)大型藻:海水中無機碳濃度和形態的變化會影響大型藻光合作用的速率,并改變其群落結構[38]。已有研究表明,在碳酸鹽飽和度較低的狀態下,夏威夷常見的殼狀珊瑚藻的鈣化率有所下降[39];(2)浮游植物:海洋酸化會導致鈣化浮游植物赫氏顆石藻(Emilianiahuxleyi)的鈣化量下降,遭受紫外線輻射的損害增加,威脅鈣化浮游植物的生存[40];而對于非鈣化浮游植物三角褐指藻(Phaeodactylumtricornutum),海洋酸化可降低其耐受高光脅迫的能力,加大光抑制,且增加其呼吸作用[41];(3)浮游動物:海洋酸化會延遲哲水蚤(Calanusglacialis)的孵化時間,并降低其孵化率[42]。海洋酸化會對日本虎斑猛水蚤(Tigriopusjaponicus)產生氧化脅迫,繼而對細胞生物大分子造成氧化損傷,最終影響其發育和繁殖力[43];(4)雙殼貝類:海洋酸化會阻礙貝類生物殼體和碳酸鈣骨架的生長,還會嚴重影響殼體和碳酸鈣骨架的內部晶體結構,甚至引起殼體和碳酸鈣骨架的消融[44-45]。海洋酸化可以降低櫛孔扇貝(Chlamysfarreri)的鈣化率和呼吸作用[46],顯著抑制紫貽貝(Mytilusedulis)碳酸酐酶活性和貝殼的生長速率[47];(5)魚類:由于魚類早期階段鰓等器官發育不完善,pH調節能力差[48],所以海洋酸化會導致魚類體液的pH值迅速下降,從而對魚類精子和卵子以及胚胎的發育具有較大的影響[49]。此外,海洋酸化對魚類的嗅覺、化學感覺和聽覺等產生不同程度的影響[50-51],進而會影響魚類的洄游、攝食和繁殖等生理行為。
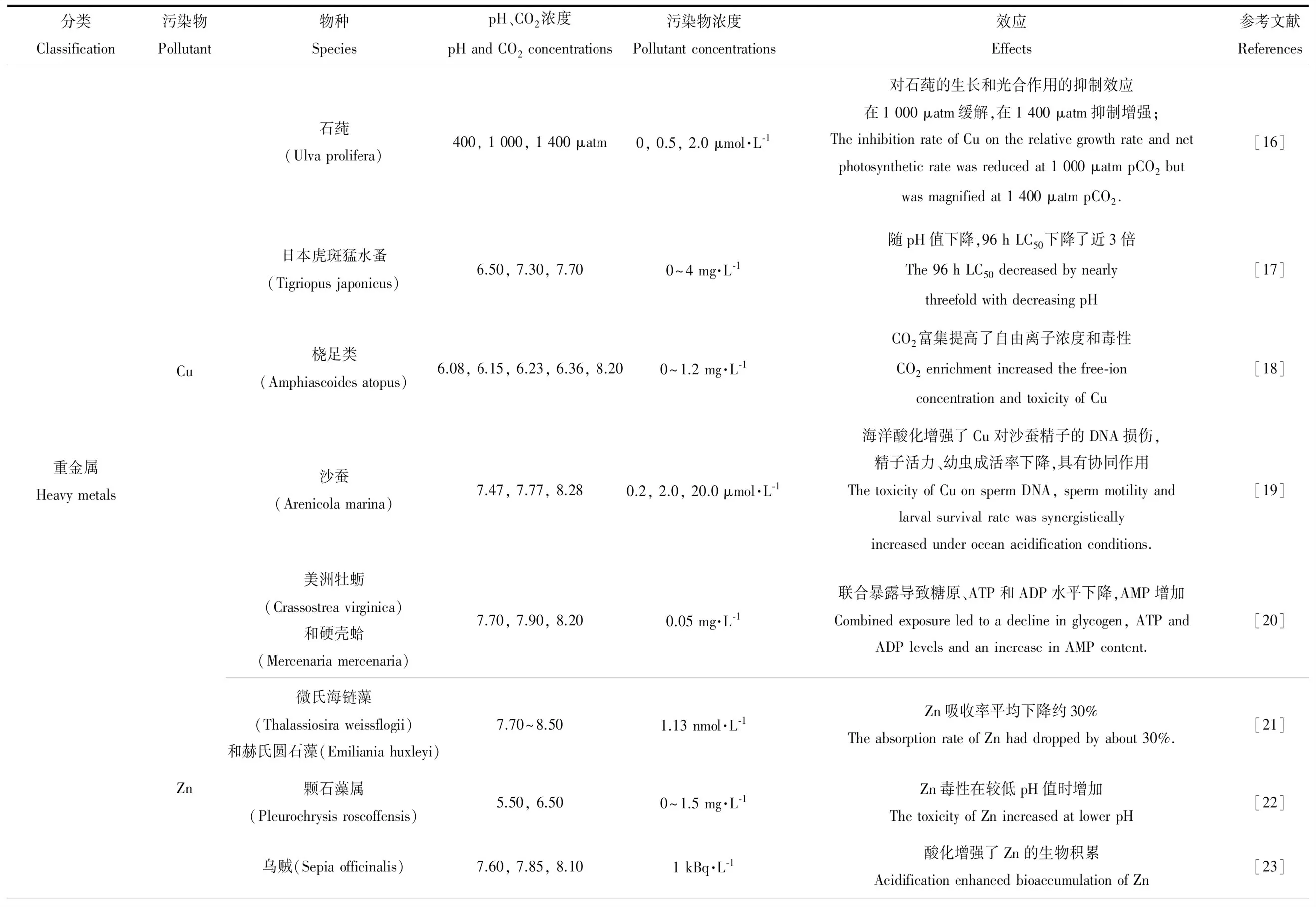
表1 海洋酸化與重金屬、有機污染物和NPs的聯合毒性Table 1 The interactions between ocean acidification and environmental pollutants
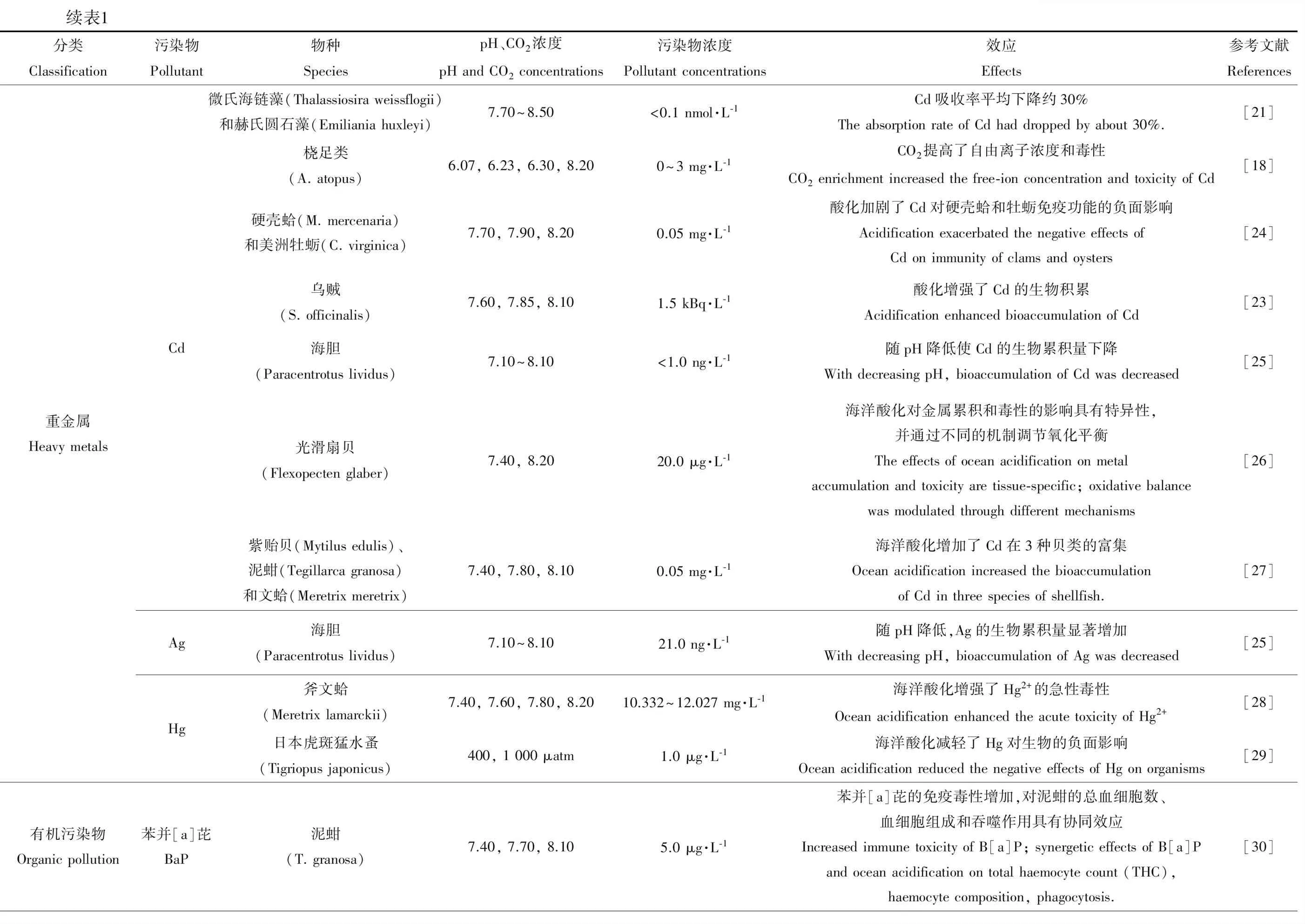
續表1分類Classification污染物Pollutant物種SpeciespH、CO2濃度pH and CO2 concentrations污染物濃度Pollutant concentrations效應Effects參考文獻References重金屬Heavy metalsCdAgHg微氏海鏈藻(Thalassiosira weissflogii)和赫氏圓石藻(Emiliania huxleyi)7.70~8.50<0.1 nmol·L-1Cd吸收率平均下降約30%The absorption rate of Cd had dropped by about 30%.[21]橈足類(A. atopus)6.07, 6.23, 6.30, 8.200~3 mg·L-1CO2提高了自由離子濃度和毒性CO2 enrichment increased the free-ion concentration and toxicity of Cd[18]硬殼蛤(M. mercenaria)和美洲牡蠣(C. virginica)7.70, 7.90, 8.200.05 mg·L-1酸化加劇了Cd對硬殼蛤和牡蠣免疫功能的負面影響Acidification exacerbated the negative effects of Cd on immunity of clams and oysters[24]烏賊(S. officinalis)7.60, 7.85, 8.101.5 kBq·L-1酸化增強了Cd的生物積累Acidification enhanced bioaccumulation of Cd[23]海膽(Paracentrotus lividus)7.10~8.10<1.0 ng·L-1隨pH降低使Cd的生物累積量下降With decreasing pH, bioaccumulation of Cd was decreased[25]光滑扇貝(Flexopecten glaber)7.40, 8.2020.0 μg·L-1海洋酸化對金屬累積和毒性的影響具有特異性,并通過不同的機制調節氧化平衡The effects of ocean acidification on metal accumulation and toxicity are tissue-specific; oxidative balance was modulated through different mechanisms[26]紫貽貝(Mytilus edulis)、泥蚶(Tegillarca granosa)和文蛤(Meretrix meretrix)7.40, 7.80, 8.100.05 mg·L-1海洋酸化增加了Cd在3種貝類的富集Ocean acidification increased the bioaccumulation of Cd in three species of shellfish.[27]海膽(Paracentrotus lividus)7.10~8.1021.0 ng·L-1隨pH降低,Ag的生物累積量顯著增加With decreasing pH, bioaccumulation of Ag was decreased[25]斧文蛤(Meretrix lamarckii)7.40, 7.60, 7.80, 8.2010.332~12.027 mg·L-1海洋酸化增強了Hg2+的急性毒性Ocean acidification enhanced the acute toxicity of Hg2+[28]日本虎斑猛水蚤(Tigriopus japonicus)400, 1 000 μatm1.0 μg·L-1海洋酸化減輕了Hg對生物的負面影響Ocean acidification reduced the negative effects of Hg on organisms[29]有機污染物Organic pollution苯并[a]芘BaP泥蚶(T. granosa)7.40, 7.70, 8.105.0 μg·L-1苯并[a]芘的免疫毒性增加,對泥蚶的總血細胞數、血細胞組成和吞噬作用具有協同效應Increased immune toxicity of B[a]P; synergetic effects of B[a]P and ocean acidification on total haemocyte count (THC), haemocyte composition, phagocytosis.[30]

續表1分類Classification污染物Pollutant物種SpeciespH、CO2濃度pH and CO2 concentrations污染物濃度Pollutant concentrations效應Effects參考文獻References人工納米顆粒NPsZnO NPsTiO2 NPsFe2O3 NPs多壁碳納米管MWCNTs厚殼貽貝(Mytilus coruscus)7.30, 8.100, 2.5, 10.0 mg·L-1低pH值和ZnO NPs在貽貝中誘導抗氧化反應,鰓比血細胞更敏感Low pH and ZnO NPs induce antioxidative responses, and gills were more sensitive than hemocyte[31]厚殼貽貝(M. coruscus)7.30, 8.100, 2.5, 10.0 mg·L-1高濃度的ZnO NPs和低pH值對貽貝產生負面效應,且具有協同作用High-concentration ZnO NPs and low pH had negative and synergistic effects on thick shell mussel[32]鯽魚(Carassius auratus)400 ppm, 600 ppm80 mg·L-1二氧化碳濃度的升高增強了ZnO NPs的Zn離子釋放量和生物有效性Elevated concentrations of CO2 enhance the release of Zn ions and bioavailability of ZnO NPs[33]小球藻(Chlorella vulgaris)7.47, 7.77, 8.200, 10 mg·L-1酸化增強了NPs的懸浮性,導致酸化和NPs的聯合作用是協同效應Acidification enhanced the suspension of NPs, resulting in a synergistic effect of acidification and NPs[14]厚殼貽貝(M. coruscus)7.30, 8.100, 2.5, 10.0 mg·L-1TiO2 NPs和低pH影響貽貝的血細胞,具有遺傳損傷和相互作用TiO2 NPs and low pH affected hemocyte responses of the mussels with interactive and genetic effects[34]厚殼貽貝(M. coruscus)2 500~2 600 μatm2.5, 10.0 mg·L-1TiO2 NPs與高pCO2對貽貝毒性具有協同效應Synergistic effects of TiO2 NPs and high pCO2 on mussel were observed[35]地中海貽貝(Mytilus galloprovincialis)6.00, 6.90, 8.100, 100, 1 000, 10 000 μg·L-1Fe2O3 NPs作為緩沖劑降低了酸化對地中海貽貝的損害Fe2O3 NPs act as a buffer to reduce acidification damage to Mytilus galloprovincialis[36]菲律賓蛤仔(Ruditapes philippinarum)7.60, 7.900, 0.1, 1.0 mg·L-1MWCNTs會影響氧化應激、生物體代謝和神經毒性,但海洋酸化不會改變其毒性MWCNTs affected oxidative stress, metabolism and neurotoxicity, butocean acidification did not change its toxicity[37]
注:LC50表示半致死濃度;ATP表示三磷酸腺苷;ADP表示二磷酸腺苷;AMP表示一磷酸腺苷。
Note: LC50is median lethal concentration; ATP is adenosine triphosphate; ADP is adenosine diphosphate; AMP is adenosine monophosphate.
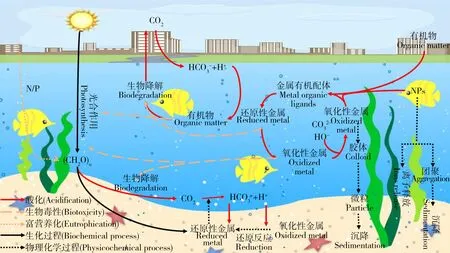
圖1 海洋酸化與環境污染物重金屬、有機污染物和人工納米顆粒(NPs)的相互作用注:根據參考文獻[15]改編。Fig. 1 Interactions between ocean acidification and heavy metals, organic pollutants and engineered nanoparticles (NPs)Note: Adapt according to reference [15].
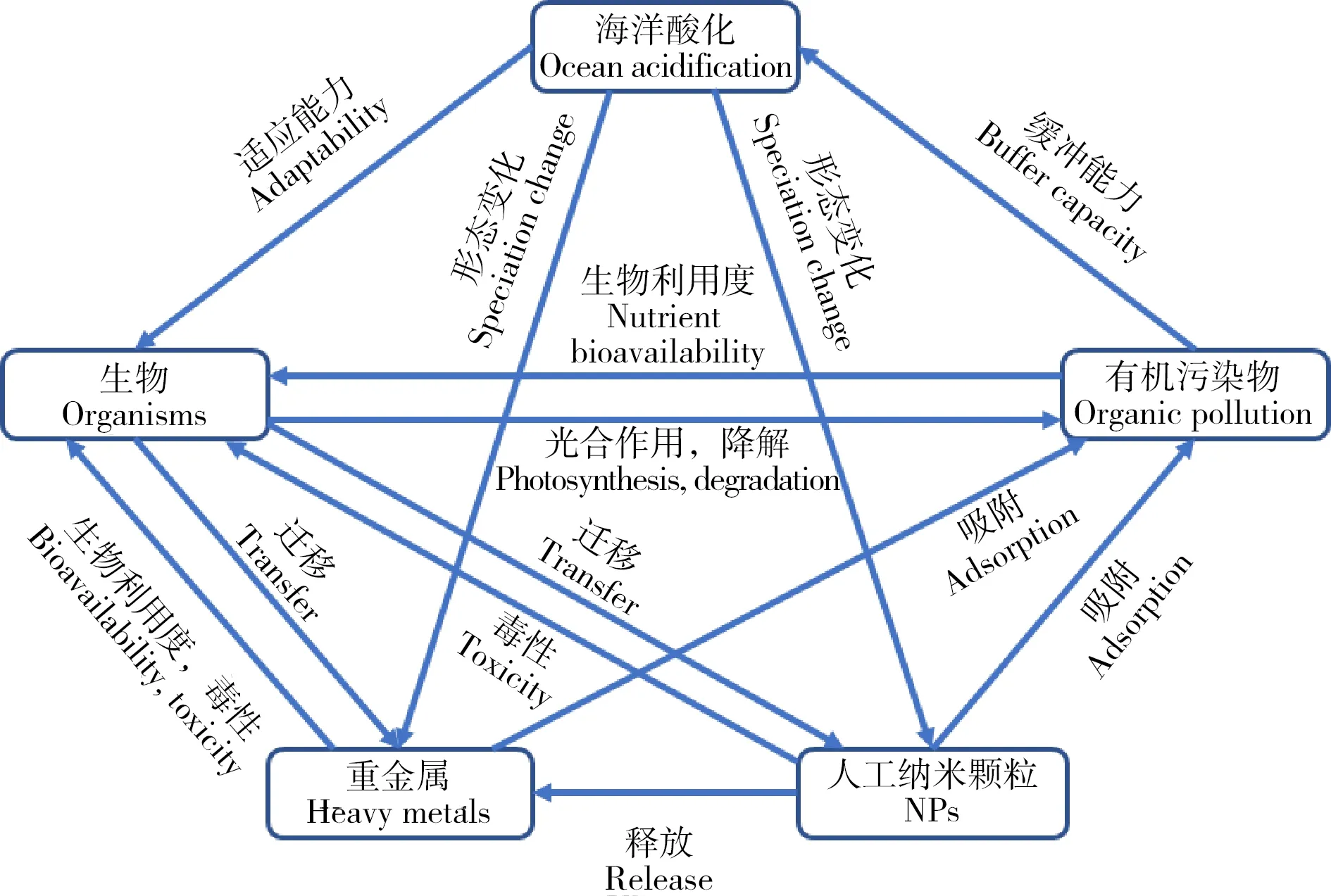
圖2 重金屬、有機污染物、NPs和海洋酸化的相互作用機制注:根據參考文獻[15]改編。Fig. 2 Interactive mechanisms of ocean acidification with heavy metals, organic pollutants and engineered nanoparticlesNote: Adapt according to reference [15].
綜上所述,不同營養級的生物個體對海洋酸化表現出不同的生理響應,長此以往會對海洋生態系統的平衡及對人類的服務功能產生影響[52]。
2 海洋酸化與重金屬的聯合毒性(Combined toxicity of ocean acidification and heavy metals)
2.1 重金屬對海洋生物的毒性效應
重金屬是海洋環境中最常見的污染物之一,主要是通過大氣輸入、陸源徑流、工業傾倒等方式進入海洋環境中,在許多近岸與河口的水體和沉積物中濃度較高[53-54]。一些金屬(如Cu和Zn)是許多生化過程中不可缺少的輔助因子,但是當金屬含量累積一定濃度時便會對生物體產生毒害作用:(1)大型藻:Han等[55]報道當Cu2+濃度達到25和100 μg·L-1時能分別抑制綠藻Ulvaarmoricana和Ulvapertusa的生長,當達到250 μg·L-1時2種藻Chl-a和Chl-b的含量均顯著降低;李欽等[56]研究發現,壇紫菜(Porphyrahaitanensis)葉狀體DNA、RNA含量和過氧化物酶(POD)活性隨Cu2+暴露劑量的增加表現出先上升后下降的時間效應關系。所以,重金屬主要是通過抑制藻細胞分裂、減少光合色素含量、降低光合放氧率、產生活性氧自由基(ROS)導致細胞氧化損傷和脂質過氧化等方式來影響藻類的生長發育過程[55-56];(2)浮游植物:低濃度的重金屬脅迫能刺激浮游植物的生長,表現出明顯的毒性興奮效應[57],高濃度時則能有效抑制其生長[58-59]。有研究表明,Cu2+脅迫能導致浮游植物體內代謝作用受阻,淀粉類物質消失,還可以通過干擾營養鹽代謝以及光合作用等對海洋浮游植物的生長產生影響[60];Hadjoudja等[61]研究證實,隨著Cu2+濃度的增加,銅綠微囊藻(Microcystisaeruginosa)和小球藻(Chlorellavulgaris)的細胞分裂率和自發熒光均減少,并且暴露時間的不同,其敏感性也有顯著差異;(3)浮游動物:浮游動物積累的重金屬相當一部分是吸附在甲殼表面的,在冠偽鏢水蚤(Pseudodiaptomuscoronatus)甲殼上的吸附量高達40%~50%,當重金屬從體表轉入體內后才會表現出毒性效應[62];海洋橈足類湯氏紡錘水蚤(Acaritatonsa)和哈氏紡錘水蚤(Acartiahudsonica)在溶解態Ag的作用下有急性致死效應,并且會導致其產卵量下降[63];高濃度的Cu和Cd會使中華哲水蚤(Calanussinicus)氨基酸含量下降,同時也抑制其攝食和排泄[64-65];(4)雙殼貝類:蔣國萍等[28]研究了海洋酸化條件下Cd2+和Hg2+對斧文蛤(Meretrixlamarckii)幼貝急性毒性效應的影響,發現單一的海洋酸化對斧文蛤幼貝的存活沒有顯著性影響,但是海洋酸化顯著增強了Cd2+和Hg2+的急性毒性;李玉環等[66]研究了Cd對海灣扇貝(Argopectenirradiams)的急性毒性影響,結果表明Cd2+對海灣扇貝是劇毒性的毒物,隨著濃度的升高,海灣扇貝會對Cd2+的毒性更加敏感;Shi等[27]研究發現,海洋酸化增加了Cd在紫貽貝、泥蚶(Tegillarcagranosa)和文蛤(Meretrixmeretrix)中的富集;周一兵等[67]的研究顯示,Hg、Cu、Zn這3種重金屬及其不同組合的聯合作用都會對菲律賓蛤仔(Ruditopesphiliprinatum)的耗氧率、排氨率及氧氮比值產生明顯的影響;(5)魚類:水體中的重金屬在魚類腎臟、肌肉等不同組織中不斷蓄積,最終可以使魚類發育畸形甚至中毒死亡。陳其晨等[68]的研究表明Zn在魚體中最容易蓄積,其次為Cu、Pb、Cd,從重金屬在魚體內不同組織的分布分析,以腎、肝、脾等部位的含量居高,肌肉、骨骼等部位則較低;同時,重金屬還會對魚類生殖細胞、胚胎、仔魚等不同生命階段的發育、存活和生長產生負面影響,從而降低子代的數量和質量[69]。
2.2 海洋酸化對重金屬環境行為的影響
海洋酸化對重金屬環境行為的影響主要歸結為3個方面:
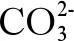
(2)海洋酸化影響重金屬的氧化還原反應。pH的變化也影響金屬的氧化和還原速率,通常,酸化對還原速率的提升高于氧化速率,因為后者對pH的依賴性較小。例如,當pH從8.1降至7.4時,海水中Fe半衰期從1 min增加到24 min[12],這說明海洋酸化可以增加還原態金屬的量[71]。
(3)海洋酸化影響與沉積物結合的金屬的行為。海洋酸化可以間接改變底泥沉積物的條件和性質,影響來自受污染沉積物的金屬通量,從而引起沉積物和海水界面主要微量元素的地球化學通量變化[72]。pH和氧化還原電位(Eh)降低可能會影響氧化還原敏感性元素的溶解度,對海洋生態系統產生負面影響[73]。
2.3 海洋酸化對重金屬生物毒性的影響
海洋酸化可以將重金屬轉化為對生物毒性更強的形態,影響海洋生物中金屬的毒性和累積及其在細胞內的結合[19]。建模研究預測,由于海洋pH值的下降,在未來100年內,河口的平均Cu2+濃度可能會增加115%[74]。已經有研究表明,Cu2+濃度為6.4×10-8mg·L-1便會對海洋浮游植物產生毒性,當pH降低時,Cu毒性將影響浮游植物的生長[75]。Nardi等[26]研究發現,相對于海洋酸化或Cd單獨暴露,光滑扇貝(Flexopectenglaber)暴露于低pH和Cd聯合作用的條件下其組織中Cd的濃度增加,并且聯合作用對扇貝的生物學毒性具有協同效應;Pascal等[18]研究也發現二氧化碳分壓(pCO2)升高,pH降低,增加了Cu2+濃度,進而導致其對橈足類Amphiascoidesatopus的毒性增強,同時,海洋酸化增強了Cu對沙蠶精子的DNA損傷,對幼體成活率抑制存在協同作用[19]。
海洋酸化的酸化程度會影響重金屬的毒性效應。海洋酸化會對石莼的生理性能產生一定的影響,pCO2濃度為1 000 μatm時會降低Cu對石莼(Ulvaprolifera)的毒性,而當pCO2增加到1 400 μatm時,Cu對石莼的毒性則會增強,這主要是因為當pCO2濃度為1 000 μatm時,H+和Cu2+的胞外競爭使得海洋酸化與重金屬出現了拮抗作用[16,76];Xu等[21]研究發現當CO2濃度升高使pH從8.5降至7.9或7.7時,浮游植物對Zn和Cd富集量平均下降約30%,從而影響其生長;通過對不同酸化條件下Zn對浮游植物半效應濃度(EC50)值影響研究發現,當pH為6.0時測得的EC50值最低,這說明隨著pH的降低,Zn對浮游植物的毒性增加[22]。
不同的重金屬在環境中的存在狀態也不同,所以海洋酸化對重金屬的毒性的影響也有一定的差異。Dorey等[25]研究了海洋酸化與放射性重金屬對地中海海膽幼體的影響,發現CO2導致的pH值變化可以改變海膽(Paracentrotuslividus)幼蟲體內重金屬的生物積累速率。而且由于重金屬的種類不同,會導致其對生物體的累積效應不同;韋曉慧等[17]研究了海洋酸化條件下Cu、Cd對日本虎斑猛水蚤的急性毒性效應,發現隨著海水pH值降低,Cu和Cd對日本虎斑猛水蚤的96 h半致死濃度(LC50)都呈減小趨勢,但是在海洋酸化條件下Cu與Cd相比,其急性毒性效應更加顯著,這是因為Cd2+在海洋中常形成氯復合物,而Cl-對pH的變化不敏感,Cu2+在海洋中常形成碳酸鹽復合物,對海洋酸化敏感,隨pH值下降釋放更多的游離Cu2+,游離Cu2+的毒性要遠遠高于復合物形態;Wang等[29]研究發現,海洋酸化顯著降低了每一代日本虎斑猛水蚤中的Hg生物積累,而且每一代的生殖抑制作用都會明顯逐漸減弱,這是因為Hg可以與氯化物形成強復合物,并且其濃度不會受到海水pH降低的影響。因此,重金屬的種類不同會導致其在海洋酸化條件下出現不同的差異[19]。
海洋酸化還會以物種特異性方式改變金屬對生物體的生理后果,最終影響其適應性和生存性。Ivanina等[24]發現pCO2升高與Cd聯合作用導致硬殼蛤(Mercenariamercenaria)和美洲牡蠣(Crassostreavirginica)血細胞中吞噬活性、粘附能力以及凝集素和熱休克蛋白(HSP70)的mRNA表達降低,從而使生物體對病原體和致病菌敏感性增強。CO2濃度升高不會影響硬殼蛤和牡蠣中的蛋白酶和和能量儲存,而且還有可能減輕微量金屬對這些生物細胞的負面影響,但是CO2濃度升高會影響Cd和Cu對海洋雙殼類的毒性,這是由于CO2濃度升高時Cu和Cd的積累增加以及高碳酸鹽和金屬聯合脅迫對能量狀態的負面影響,并可能影響雙殼類群體的健康以及受污染河口的金屬營養轉移[20]。
3 海洋酸化與有機污染物的聯合毒性(Combined toxicity of ocean acidification and organic pollutants)
3.1 有機污染物對海洋生物的毒性效應
有機污染是世界上最普遍的污染類型之一,也是驅動海洋生態系統變化的主要人為因素。環境中的持久性有機污染物(POPs)具有長期殘留性、生物蓄積性、半揮發性和高毒性等特征,會對海洋生物產生不利的影響:(1)浮游植物:Huang等[77]研究發現,高濃度的石油污染降低了浮游植物的葉綠素a含量和細胞密度,極大抑制了其生長;研究表明當全氟辛烷磺酸鹽(PFOS)的濃度高于50 mg·L-1時,羊角月牙藻(Selenastrumcapricornutum)和小球藻的生長會被明顯抑制[78]。Bopp等[79]的研究發現,多環芳烴(PAHs)可以影響假微型海鏈藻(Thalassiosirapseudonana)的脂肪酸代謝和二氧化硅殼層的形成;(2)雙殼貝類:當有機污染物被生物機體吸收后,在雙殼軟體動物體內會產生一系列的生物化學反應,并產生大量的ROS,從而產生氧化損傷[80]。蔡文貴等[81]通過研究鄰苯二甲酸二乙基己酯(DEHP)對翡翠貽貝(Pernaviridis)生化指標的影響,發現DEHP可以影響翡翠貽貝的內臟和外套膜抗氧化防御系統酶,并且短期內無法消除;(3)魚類:叢偉[82]研究了PAHs對海洋魚類胚胎發育的神經毒性,發現苯并[a]芘能引起仔魚出現一系列異常癥狀并誘導CYPIA與AHR2的表達。采用三環的菲(PHE)暴露后,雄性褐菖鲉魚(Sebastiscusmarmoratus)性體比指數和精子比例隨著濃度升高呈現U形曲線變化,同時還會抑制雌性黑點青鳉(Oryziasmelastigma)卵巢發育,產卵頻率和產卵數量均減少,影響卵子質量和F1代胚胎發育心率的異常,降低F1代仔魚的出膜率和出膜后的正常游動[83]。
3.2 海洋酸化對有機污染物降解的影響
海洋酸化會改變金屬形態及其生物有效性,進而影響有機污染物的降解(圖2)。Fe是降解碳氫化合物的重要因素,它可能會影響有機污染物的催化和油類氧化分解所需的酶活性[84]。在惡劣條件下,幾種關鍵酶在以Fe作為限制條件下會導致芳烴甲苯降解的活性降低[85-86]。Shi等[87]預測,到2100年的pCO2水平可能會將生物學上可用的Fe3+降低10%~20%,這可能是由于pH降低引起的Fe與有機配體的結合。因此,海洋酸化而導致Fe的生物利用度下降會間接對有機污染物的降解產生潛在的影響。
氮是促進PAHs和其他有機污染物降解的關鍵因素[87]。作為海洋中最重要的氮源之一,固定氮是限制光合生物生長的營養物質,對于藻類和海洋細菌來說,固定氮是海洋細菌和古細菌的能源或氧化劑[21],海洋酸化降低了在海洋模型中藍藻主導的N2固定的效率[21,87],從而影響了PAHs和其他有機污染物的降解。此外,隨著NH3底物從體系中消失,硝化速率可以在pH為6.0~6.5時下降至零[88],也會對有機污染物的降解產生影響。據報道,大氣CO2升高導致海洋含氧量下降。O2既是微生物降解有機物中最重要的元素之一,也是有機物初始分解所必需的,與正常情況相比較,較低的含氧量降低了微生物活性,導致降解率降低甚至厭氧降解[89]。
3.3 海洋酸化對有機污染物毒性的影響
海洋中的有機污染物可以被膠體有機物質吸收或結合并固定在有機金屬礦物復合物中,超過50%的有機質被約束在海洋的有機金屬礦物復合物中[90]。海洋酸化引起的金屬離子溶解可能會破壞有機金屬配合物,從而增強有機污染物的釋放和毒性。同時由于酸化會抑制有機污染物的生物降解,導致有機污染物長期存在于海洋環境中[89,91]。在無脊椎動物中的研究表明,海洋酸化和苯并[a]芘均可誘導長牡蠣(Crassostreagigas)血細胞凋亡,因此可能導致總血細胞數減少和血細胞組成改變[92-93]。Su等[30]研究發現,在海洋酸化條件下,苯并[a]芘的免疫毒性增加導致文蛤總血細胞減少、血細胞組成發生改變以及吞噬作用受阻,這表明海洋酸化與苯并[a]芘具有協同作用。在pCO2升高的壓力下,海洋生物可能改變能量分布,并為基礎代謝和體內調節等關鍵生命過程分配更多能量[94]。因此在未來海洋酸化條件下,可用于污染物防御和處理能量的減少也可導致苯并[a]芘增加生物體的免疫毒性。此外,海洋酸化可能損害海洋生物的表皮組織,從而導致更多的有機污染物如苯并[a]芘富集到海洋生物體內,增強對海洋生物的毒性效應[27,95]。
4 海洋酸化與NPs的聯合毒性(Combined toxicity of ocean acidification and engineered nanoparticles)
4.1 NPs對海洋生物的毒性效應
納米材料的大量生產和廣泛應用勢必會導致大量NPs進入到海洋環境中,進而對海洋生物產生毒性效應:(1)浮游植物:NPs對浮游植物的影響與其材質,粒徑和浮游植物的種類有關。ZnO NPs (≥10 mg·L-1)會顯著抑制硅藻(角毛藻和海鏈藻)的生長,并在海鏈藻中發生明顯富集[96]。TiO2NPs可以進入到正常新月菱形藻(Nitzschiaclosterium)的細胞內,對藻細胞產生氧化損傷,導致微藻脂質過氧化,影響藻細胞膜的穩定性。胞內ROS是TiO2NPs對海洋微藻產生氧化損傷的主導因素[97];(2)浮游動物:浮游動物營懸浮生活,這一生活習性決定其很容易受到懸浮NPs的影響。浮游動物在攝食時能夠同時過濾大量的海水,這就可能導致大量的NPs進入到浮游動物體內,對浮游動物的正常生長造成影響。Snell和Hicks[98]報道經過2 d暴露后,聚苯乙烯NPs(≥83 nm)主要富集在臂尾輪蟲(Brachionusmanjavacas)的胃和腸部,而37 nm聚苯乙烯NPs可以穿過輪蟲的腸壁孔徑富集到生物的不同組織。這表明粒徑是影響輪蟲對NPs吸收的重要因素,并會因此產生不同的毒性效應;(3)雙殼貝類:Shi等[99]研究了泥蚶暴露于TiO2NPs環境濃度的免疫毒性,結果表明在濃度為10和100 mg·L-1的TiO2NPs暴露30 d后,血細胞總數、吞噬活性和紅細胞比例都顯著降低。此外,當暴露于高濃度的TiO2NPs時,泥蚶體內神經遞質濃度顯著增加,并且抑制乙酰膽堿酯酶(AChE)活性和下調神經遞質相關基因的表達,這表明TiO2NPs對泥蚶具有神經毒性[100]。Canesi等[101]發現,當TiO2NPs濃度為1~10 mg·L-1時可對貽貝引起毒性作用。厚殼貽貝(Mytiluscoruscus)暴露在TiO2NPs中會導致其粘液中吸附大量的TiO2NPs,這可能會影響貽貝的呼吸[34]。CuO NPs進入水體中可以釋放一定量的Cu2+,納米材料和其釋放的Cu2+均能夠對櫛孔扇貝的血細胞造成一定的損傷[102];(4)魚類:Cong等[103]研究了ZnO NPs對黑點青鳉的胚胎毒性,結果表明,暴露20 d后,黑點青鳉胚胎的死亡率和心率增加,孵化率降低,孵化時間增長,畸形率增加。此外,ZnO NPs對黑點青鳉死亡率和心率的影響要大于釋放的Zn2+,這表明ZnO NPs的毒性主要來自尺寸效應。Vignardi等[104]報道了鯧鲹(Trachinotuscarolinus)對TiO2NPs的富集以及NPs產生的基因毒性和細胞毒性,結果表明,NPs主要富集在生物體的肝臟、腎臟和鰓部,少量進入到肌肉部位,并且NPs會對生物產生DNA損傷,降低紅細胞數量。
4.2 海洋酸化對NPs環境行為的影響
NPs進入海洋環境后,會發生一系列復雜的物理化學過程(團聚、溶解以及生物轉化等)。NPs的懸浮性能以及離子釋放能力與海水環境介質參數(pH、離子強度和天然有機物等)緊密相關。海洋酸化對NPs環境行為的影響主要有:
(1)NPs的懸浮性。當納米膠體顆粒粒徑<100 nm時,由于布朗運動而導致膠體顆粒物之間碰撞并在短時間內發生團聚。隨著團聚體尺寸越來越大,其擴散系數變得越來越小,團聚速度逐漸變慢,最后達到平衡狀態。此外,在海洋環境中,由于海水的高離子強度,NPs更易于發生團聚,而pH等條件對NPs的懸浮性能有著很大的影響[35]。有研究表明在pH=7.77和pH=7.47的海水中TiO2NPs的水力直徑要小于在pH=8.20的海水中,分散性增強,這表明海洋酸化能夠抑制TiO2NPs的團聚,增強其懸浮性能[14]。Mohd Omar等[105]研究發現,當pH小于等電點時,隨著pH的升高,NPs的團聚程度逐漸增大。當pH等于等電點的時候NPs團聚程度最大。當pH大于等電點時,NPs隨pH的增加而開始分散。所以當海洋酸化導致pH降低時,NPs的懸浮穩定性會發生變化。
(2)金屬NPs的離子釋放能力。海洋酸化可以導致pH下降,進而改變金屬NPs的離子釋放能力。Zhao等[106]的研究發現pH值變化促進了Cu NPs的離子釋放(公式1),在對ZnO和CuO NPs的研究中發現,pH值可以改變ZnO和CuO NPs的物理化學性質,在海洋酸化條件下,海水中CO2濃度升高,導致海水中H+增加、pH降低,進而導致公式1向右移動,使得Zn2+和Cu2+的釋放量增加。但是有研究表明,當pH大于7時,ZnO NPs在較高pH條件下的溶解度較低,這可能是由于高pH條件下在ZnO NPs表面形成了氫氧化物層,從而抑制了其在堿性環境下的溶解(公式2)[33,103,107]。已有研究表明,金屬NPs的離子釋放是導致其產生毒性的主要原因之一,因此研究海洋酸化對金屬NPs離子釋放能力的影響十分必要[108-109]。
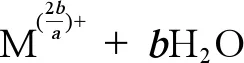
(1)

(2)
4.3 海洋酸化對NPs毒性效應的影響
NPs的毒性效應一般可以分為2種,即直接毒性效應和間接毒性效應。前者是指NPs本身物理性質(尺寸、形狀、電荷等)產生的毒性反應,而后者是指由于NPs的其他特性,如金屬NPs能夠溶解產生離子對生物造成的毒性效應。因此,一方面海洋酸化通過促進NPs的分散,增強其懸浮性能,減弱其在海水中的團聚,從而使其作用于生物體的NPs的水力直徑減小,增強其毒性效應。Xia等[14]的研究發現,海洋酸化增強了TiO2NPs對小球藻生長的抑制作用,同時增加了TiO2NPs對藻細胞的氧化損傷,這是因為海水pH降低使TiO2NPs在海水中的水力直徑減小,懸浮性能增強,從而導致藻細胞中富集的TiO2NPs含量升高,進而增強了TiO2NPs對藻類的毒性效應。Huang等[34]研究了海洋酸化條件下TiO2NPs對厚殼貽貝血細胞免疫參數的綜合效應,結果表明海洋酸化與TiO2NPs聯合作用會使厚殼貽貝血細胞功能受損,血細胞死亡數和ROS升高,并且海洋酸化與高劑量的TiO2NPs(10 mg·L-1)對厚殼貽貝存在協同作用;Hu等[35]也發現海洋酸化條件下TiO2NPs對厚殼貽貝的生理功能的損害增大。但是海洋酸化條件下TiO2NPs對貝類毒性增強的原因未作探討。另一方面,海洋酸化促進了金屬NPs的離子釋放,從而增強了NPs的毒性效應。Yin等[33]研究發現,CO2升高使pH值降低,ZnO NPs在水中釋放Zn離子量是升高,增加了鯽魚(Carassiusauratus)肝臟、腦和肌肉中Zn的富集含量,從而增強了ZnO NPs對鯽魚的氧化損傷。Huang等[31]研究了海洋酸化與ZnO NPs對厚殼貽貝的聯合毒性效應,發現在低pH和ZnO NPs的單獨作用下,厚殼貽貝腮和血細胞超氧化物歧化酶(SOD)、過氧化氫酶(CAT)、谷胱甘肽過氧化物酶(GPx)、酸性磷酸酶(ACP)和堿性磷酸酶(ALP)活性均升高,但是海洋酸化與ZnO NPs沒有顯著的交互作用,即海洋酸化對ZnO NPs的毒性效應沒有顯著影響。但是,Wu等[32]研究發現海洋酸化與ZnO NPs對厚殼貽貝血細胞吞噬作用、酯酶活性、溶酶體含量、血細胞死亡率以及ROS含量具有顯著的協同效應。這表明海洋酸化與ZnO NPs對厚殼貽貝的聯合作用模式與測定的生化指標具有一定相關性。此外,Kadar等[36]研究發現海洋酸化增加了地中海貽貝(Mytilusgalloprovincialis)D形幼蟲的畸形率,而Fe2O3NPs對貝類胚胎發育沒有顯著影響,但是二者聯合作用下Fe2O3NPs作為緩沖劑降低了海洋酸化對地中海貽貝的損害,這可能是由Fe2O3NPs的毒性較小所致。對于非金屬NPs與海洋酸化的聯合毒性效應也進行了研究。如De Marchi等[37]研究了海洋酸化條件下多壁碳納米管(MWCNTs)對菲律賓蛤仔的毒性效應,發現在低pH和正常pH條件下MWCNTs均可以誘導菲律賓蛤仔的氧化應激,影響其能量代謝,并產生神經毒性,但是低pH條件下MWCNTs的毒性與正常pH無顯著差異,表明海洋酸化對MWCNTs的毒性沒有顯著影響。基于以上研究,海洋酸化與NPs的聯合毒性作用及其機制尚不清楚,還需進一步研究。
5 研究展望(Research prospect)
全球氣候變化條件下污染物的海洋環境效應是一個新興的交叉性科學問題,特別是海洋酸化與污染物對海洋環境的聯合效應更加值得關注。本文系統梳理了海洋酸化與典型污染物(重金屬、有機污染物)和新型污染物(NPs)的聯合毒性效應,表明海洋酸化會影響重金屬、有機污染物和NPs的環境行為,從而改變其對海洋生物的毒性效應。但是目前海洋酸化與污染物的聯合毒性效應的研究還很少,其致毒機制尚不明確。所以,對下一步開展的研究工作展望如下:
(1)目前的研究主要集中于海洋酸化與重金屬的聯合毒性效應,而與有機污染物、NPs及其他污染物的研究還很少,特別是海洋酸化條件下污染物在海洋食物鏈上的營養傳遞及生物毒性研究還沒有報道。海洋酸化與污染物的聯合毒性作用模式的影響因素及調控機制還不確定,機理性的研究還需進一步加強;
(2)目前的報道大多是研究IPCC預測的海洋酸化pH值與環境污染物的聯合毒性效應,但是CO2濃度升高造成海洋酸化是一個漸進的過程,所以應設置更多的pH梯度來研究這種pH漸進情況下海洋污染物的環境行為及其對海洋生態系統的影響,從而更加客觀地反映海洋酸化過程中污染物的海洋環境效應;
(3)目前的研究主要集中于室內模擬實驗,研究對象也主要是單一的生物物種,不能全面反應實際海洋環境中酸化與污染物的聯合作用對海洋生態系統的真實影響,所以下一步應采用微宇宙(Microcosm)和中宇宙(Mesocosm)生態系統原位研究海洋酸化與污染物的聯合毒性,對海洋生物群落結構的影響也應加以關注;
(4)除海洋酸化外,海水升溫、海洋低氧、UV-B輻射等都是全球氣候變化下海洋環境的改變,它們會與環境污染物共同作用于海洋生態系統[110]。所以需要更加系統地研究全球氣候變化耦合海洋污染對海洋生物的毒性效應,為準確評估全球氣候變化下的海洋生態風險提供科學依據。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
核科學與工程(2021年4期)2022-01-12 06:30:26
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
當代陜西(2021年2期)2021-03-29 07:41:24
今日農業(2020年19期)2020-12-14 14:16:52
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
中學物理·高中(2016年12期)2017-04-22 11:53:03
中國塑料(2016年3期)2016-06-15 20:30:00
