不同生境下的象頭花光合生理特性探究
2019-07-02 07:32:58陳金珠黎云祥楊佳何理權(quán)秋梅
生態(tài)科學(xué) 2019年3期
陳金珠,黎云祥,楊佳,何理,權(quán)秋梅,*
1.西華師范大學(xué)生命科學(xué)學(xué)院,南充 637002
2.西華師范大學(xué)環(huán)境科學(xué)與工程學(xué)院,南充 637002
0 前言
光合作用的研究歷史悠久,1772年P(guān)riestly 便揭示了光合作用就是呼吸作用的逆反應(yīng)[1]。光合作用作為植物生命的維系,是所有植物生存和發(fā)展最為關(guān)鍵的生理過程。同時,光合作用受到生態(tài)環(huán)境的影響,這使得植物形成了適應(yīng)環(huán)境的形態(tài)結(jié)構(gòu)和生理調(diào)節(jié)特征[2]。目前,對環(huán)境因子影響植物光合作用的理解來自植物光合作用對光的響應(yīng)[3-4]。通過恰當?shù)墓夂夏P蛿M合植物光響應(yīng)曲線可得到許多具有參考價值的光合生理參數(shù)(諸如最大凈光合作用速率Pmax、光飽和點LSP、光補償點LCP、表觀量子效率AQY以及暗呼吸速率Rd),可用以描述植物光合作用能力及環(huán)境適應(yīng)性能評價[5-6]。國內(nèi)外已建構(gòu)的多種光合模型,不同模型對不同植物的適應(yīng)性各不相同[7-11]。因此,對于模型的選取應(yīng)當對比驗證,需考量模型的準確性以及與實際測量值的相符程度。除模型擬合外,當前進行了諸多控制性試驗,通過人工控制外部環(huán)境因子水分、溫度、光照等,從而改變植物氣孔導(dǎo)度、蒸騰與光合作用速率,影響植物的生長狀況[12-15]。但對于自然生境中植物的光合速率變化的探索仍相對匱乏。
在對天南星屬(Arisaema)植物的研究和文獻調(diào)查中顯示象頭花(Arisaema franchetianumEngl.)為我國特有物種。具藥用功能,以塊莖入藥,有解毒消腫,散結(jié)止痛之效[16-17]。中藥質(zhì)量評價當中象頭花抗腫瘤和鎮(zhèn)咳祛痰效果較佳,現(xiàn)已能從其塊莖中分離出18 種化合物主要是生物堿及酰基甘油糖苷[18-19]。現(xiàn)已知對該屬植物的研究主要集中在化學(xué)成分提取及分析[19-20]、藥理作用研究[21-22]、種植生產(chǎn)等方面[23]。為合理的開發(fā)象頭花資源,我們探究不同的自然環(huán)境因子下象頭花光合生理特性,為象頭花的生理生態(tài)和栽培繁殖研究提供參考。
1 材料與方法
1.1 研究區(qū)域概況
唐家河國家級自然保護區(qū)(32°32′—32°41′N,104°36′—104°56′E),隸屬四川省青川縣,岷山山系,總面積40000 hm2。該區(qū)雨量充沛,溫暖濕潤,屬亞熱帶季風氣候。1月平均溫度1.8 ℃,最高溫度5.3 ℃,最低溫度-11 ℃,6—8月氣候溫涼,平均溫度13.1— 14.2 ℃,最高為22.6—24.7 ℃。年降雨量1100—1300 mm,無霜期180 d[25]。整個保護區(qū)植被類型包括山地常綠闊葉林、常綠與落葉闊葉混交林、針闊混交林、亞高山針葉林和高山灌叢與高山草甸[26],土壤類型有山地黃壤、山地黃棕壤、山地灰棕壤和山地棕色灰化土等[25]。其豐富的植被與土壤資源,孕育了大量的野生動植物。
1.2 實驗方法
實驗于8月22 號至8月24 號,晴朗天氣的上午進行。因其地下塊莖多年生,地上部分每年死亡[16],我們在林緣、林下兩種常見生境各選取3 株長勢相近的健康植株,測量其生長旺盛的新葉的光響應(yīng)曲線。具體而言,使用Li-6400XT(LI-COR,Lincoh,USA)便攜式光合作用測量系統(tǒng),紅藍LED 光源,Light curve2自動測量系統(tǒng),CO2濃度400 μmol·mol-1,葉室溫度25 ℃,空氣流速500 μmol·s-1,光合有效輻射梯度為2000、1800、1600、1400、1200、1000、800、500、250、120、60、30、15、0 μmol·m-2·s-1。而后,通過模型(表2)擬合光合有效輻射PAR與光合速率Pn,繪制曲線圖,獲取光合參數(shù),篩選出最適模型;分析光響應(yīng)曲線同步輸出的光合因子參數(shù)(氣孔導(dǎo)度Cs、蒸騰速率Tr、胞間CO2濃度Ci)和環(huán)境因子參數(shù)(樣品室CO2濃度CO2S、空氣溫度Tair、樣品室相對濕度RH-S、葉室外光合有效輻射PARO)與Pn的相關(guān)性并繪制曲線圖。為初步比較林緣、林下兩種生境環(huán)境因子的差異,在測量期間使用GARMIN GPS 導(dǎo)航儀定經(jīng)緯度與海拔定位;利用FIELDSCOUT3413 照度計、KIMO HD100 溫濕度計分別測量兩種生境中各10 個個體周圍的光照、溫度、濕度三個環(huán)境因子。最后,為探討飽和光強下不同生境植株這些因子的差異顯著性以及對植株長勢的影響,我們根據(jù)光響應(yīng)曲線得出的飽和光強,將光合輻射設(shè)為500 μmol·m-2·s-1其余設(shè)定同光響應(yīng)曲線,測量飽和光強下兩種生境中各10 個個體的光合因子,并記錄每個測量個體株高、直徑、葉長、葉寬。每次實驗至少3 次重復(fù),最后取用均值。
2 數(shù)據(jù)分析
2.1 數(shù)據(jù)分析模型及檢驗公式
在曲線擬合的過程當中,選取了最為常見的五種模型如表1進行擬合。各式相同的字符含義相同。(1)式中Pn:凈光合速率;φ:初始量子效率;Pmax:最大凈光合速率;I:光合有效輻射;Rd:暗呼吸速率;(2)式中θ:光響應(yīng)曲角,反應(yīng)光響應(yīng)曲線彎曲程度,取值0<θ≤1;公式(1)、(2)計算時需要估算植物的飽和光強,為了估算植物的飽和光強,就需要用直線方程擬合弱光強條件下 (PAR≤200 μmol·m-2·s-1)的光響應(yīng)數(shù)據(jù)得到表觀量子效率(AQY),然后解直線方程:Pmax=AQY×LSP-Rd可以得到光飽和點(LSP)[26-27]。(3)式α:I=0 和I=Ic 時,兩點連線的斜率;β:修正系數(shù);γ:為光飽和項,光響應(yīng)曲線初始斜率與最大凈光合速率之比,即γ=α/Pmax;LCP:光補償點[9]。公式(4)估算植物的飽和光強時需要假定光合速率為0.9Pmax或 0.99Pmax(5)式中當I=0 時,Rd=0;當Pn=0 時,光補償點當I=0 時,光響應(yīng)曲線在該點的斜率為內(nèi)稟量子效率φ0=Pn′(I=0=-αβ+γε);當αβ=γε時,Pn′=0,Pn存在最大值;公式(3)、(5)可以直接計算植物的飽和光強和最大凈光合速率[10-11]。
1.4 數(shù)據(jù)處理
數(shù)據(jù)基本處理、光合參數(shù)計算、作圖均由Microsoft Excel 2010 完成,模型公式基本參數(shù)計算、皮爾遜相關(guān)性檢驗、單因素方差分析均由SPSS 23.0 完成。
3 結(jié)果與分析
3.1 五種模型光響應(yīng)曲線擬合
5 種模型的象頭花凈光合速率光響應(yīng)曲線如圖1所示。由圖1A 看出林緣象頭花的凈光合速率Pn隨著光合有效輻射PAR的增加先迅速增加(PAR<250 μmol·m-2·s-1),而后緩慢增大至光飽和點LSP。達到500 μmol·m-2·s-1時,隨著PAR增強Pn呈現(xiàn)出略微下降的趨勢,故林緣象頭花的光響應(yīng)曲線在強光階段(PAR>500 μmol·m-2·s-1)的表現(xiàn)為弱度抑制型[28]。圖1B 中林下象頭花的光響應(yīng)曲線在PAR<500 μmol·m-2·s-1階段的走勢與林緣象頭花走勢相近,而后隨著PAR增強Pn呈現(xiàn)出明顯的下降趨勢,因而林下象頭花的光響應(yīng)曲線在強光階段表現(xiàn)出強度抑制型[28]。
光合生理參數(shù)常用于衡量植物的光合作用能力,但諸如光補償點LCP、最大凈光合速率Pmax等無法進行直接的測量,模型模擬計算逐漸的成為了計算光合生理參數(shù)主要手段[5]。象頭花模型擬合參數(shù)見表2。其中,決定系數(shù)R2反應(yīng)了擬合的精確程度,其越接近1 表示相關(guān)方程的參考價值越高[29]。林緣、林下兩種生境中的R2均大于0.9,說明5個模型均滿足擬合的精度要求。從R2來判斷,非直角雙曲線模型、直角雙曲線修正模型、指數(shù)模型、改進指數(shù)模型對林緣的光響應(yīng)曲線擬合效果均較好;林下的光響應(yīng)曲僅有改進指數(shù)模型具有較好的擬合效果。無論是在林緣還是林下5 個模型均能一定程度的擬合出光響應(yīng)過程中的生理參數(shù),但其中4 個模型擬合的LSP均遠低于實測值,僅有改進指數(shù)模型能較好的擬合出LSP。此外,改進指數(shù)模型能較好的擬合出最大凈光合速率Pmax、光補償點LCP、暗呼吸速率Rd與表觀量子效率AQY,其擬合值均與實測值較為相符。鑒于此,改進指數(shù)模型為林緣、林下兩種生境象頭花光響應(yīng)曲的最適擬合模型。

表1 五種光響應(yīng)模型 Table1 Five kinds of light response models
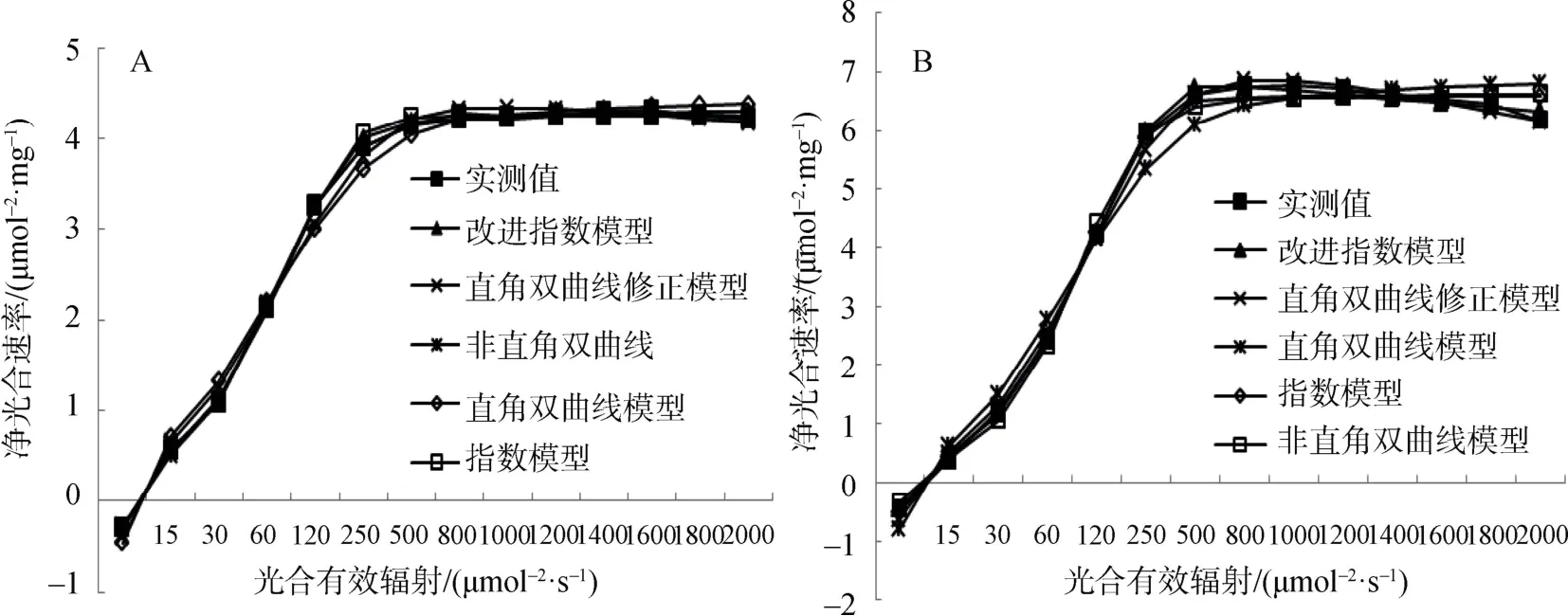
圖1 五種模型擬合不同生境象頭花的光響應(yīng)曲線,林緣(A)、林下(B)) Figure1 Photosynthesis-light response curve of five fitted models of A.franchetianum Engl.in different habitats under-canopy (A) and gap edge (B)
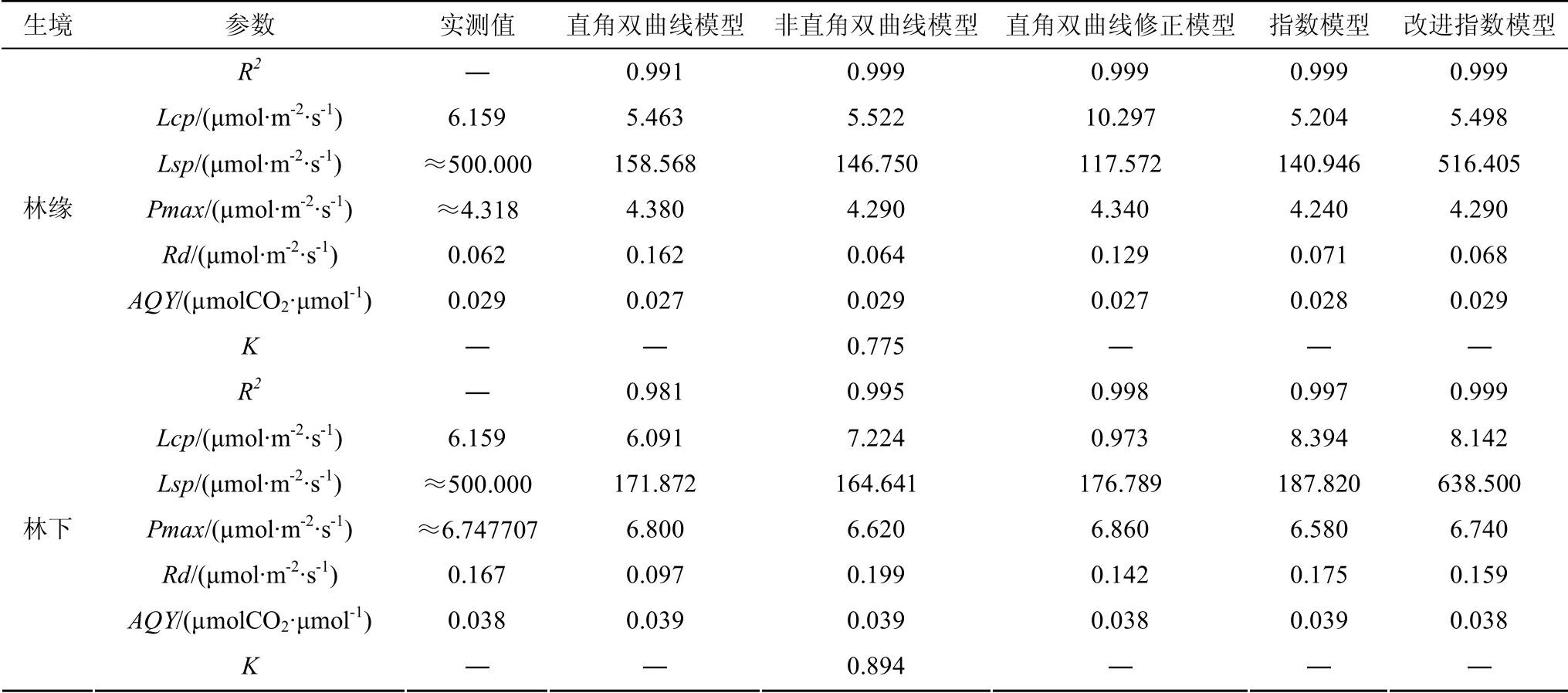
表2 5 個模型各項光合參數(shù)與實測值的比對 Table2 Comparison of the photosynthetic parameters of five models with measured values
3.2 凈光合速率與光合因子的相關(guān)性
由表3可知,林緣象頭花的凈光合速率Pn與氣孔導(dǎo)度Cs、蒸騰速率Tr、空氣溫度Tair、和樣品室相對濕度RH-S的相關(guān)系數(shù)分別為 0.902**、0.914**、0.814**、0.894**,呈顯著正相關(guān)關(guān)系(P<0.01);而與胞間CO2濃度Ci、樣品室CO2濃度CO2S的相關(guān)系數(shù)分別為-0.951**、-0.981**,呈顯著負相關(guān)關(guān)系(P<0.01);林下象頭花的Pn與Cs、Tr、Tair、RH-S和PARO的相關(guān)系數(shù)分別為0.931**、0.907**、0.883**、0.903**、0.725**,呈顯著正相關(guān)關(guān)系(P<0.01);而與Ci的相關(guān)系數(shù)分別為-0.972**,呈顯著負相關(guān)關(guān)系(P<0.01);與CO2S無明顯的相關(guān)性。
3.3 光合因子隨光照輻射的變化
如圖2A 所示,隨光照輻射的增強,林緣、林下象頭花葉片的氣孔導(dǎo)度Cs均呈現(xiàn)出上升趨勢,在PAR為800μmol·m-2·s-1時達到峰值;且林下生境中象頭花葉片的Cs始終高于林緣生境中象頭花葉片Cs。圖2B 中林緣、林下兩種生境中的胞間CO2濃度Ci均隨著光照輻射的增強呈現(xiàn)出明顯下降的趨勢,且林緣的Ci在光照輻射的增強的整個過程中幾乎均低于林下的Ci。圖2C 中,林緣、林下象頭花葉片的蒸騰速率Tr與Cs的走勢一致,均隨光照輻射的增強呈現(xiàn)出上升趨勢,且在PAR為800 μmol·m-2·s-1時達到峰值,林緣象頭花葉片的Tr在相同的光照輻射下總是低于林下象頭花葉片的Tr。圖2D 中的水分利用效率WUE來自光合速率與蒸騰速率的比WUE=Pn/Tr,其反應(yīng)了植物對水分的利用情況[30]。由圖2D可知,林緣、林下象頭花葉片的WUE均隨光照輻射的增強呈現(xiàn)出先上升(PAR<250 μmol·m-2·s-1)而后處于3.5 μmolC02·molH2O-1水平呈上下波動,波動范圍在34 μmolC02·molH2O-1。最后,圖2E 中的林緣、林下象頭花葉片的Pn隨光照輻射的增強呈現(xiàn)出先上升(PAR<250 μmol·m-2·s-1)的趨勢,而后林緣Pn趨于穩(wěn)定水平,僅林下Pn呈現(xiàn)出略微下降趨勢;但自(PAR>120 μmol·m-2·s-1)開始,同等光照強度下林下Pn總是大于林緣Pn。
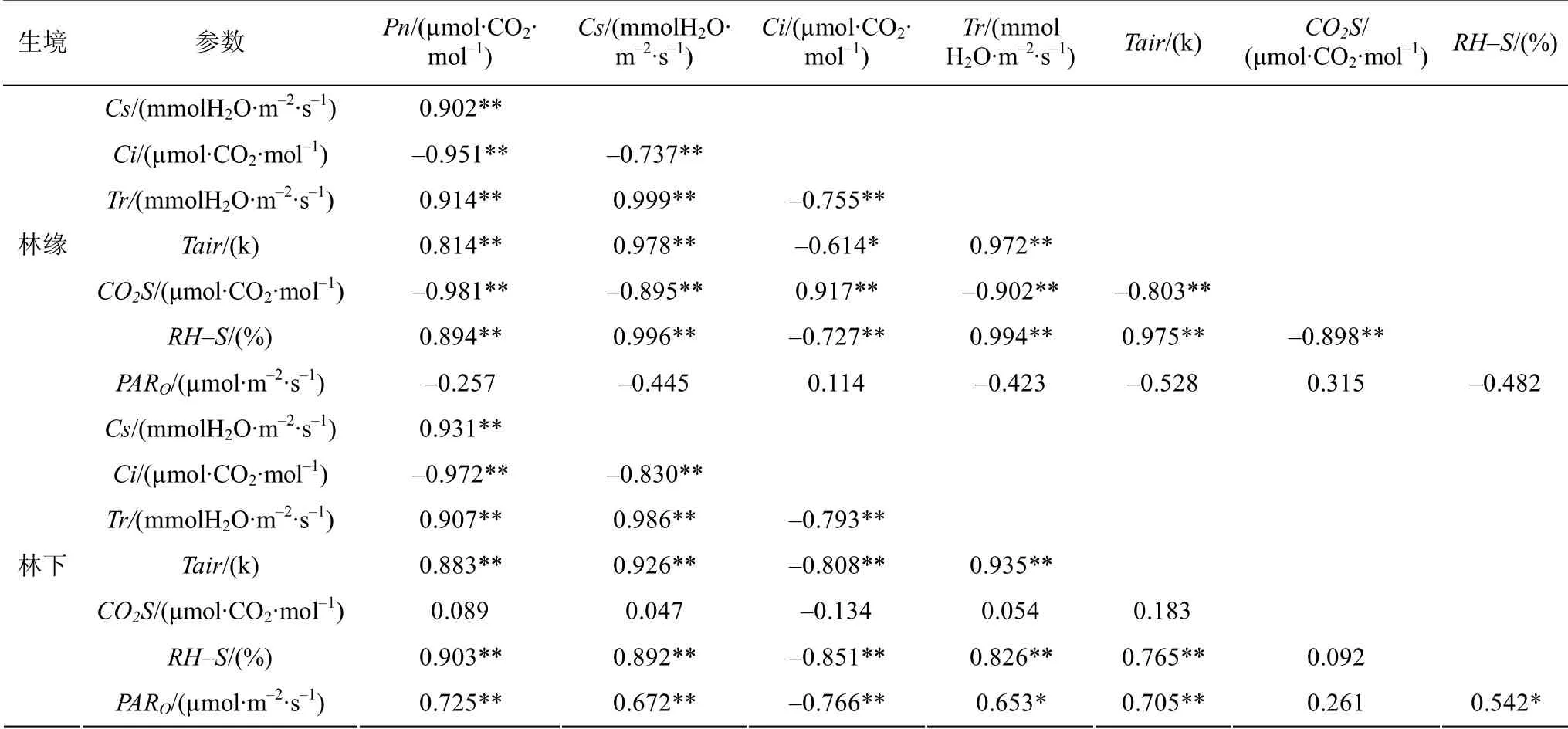
表3 不同生境凈光合速率參數(shù)相關(guān)性分析 Table3 Correlation analysis of Pn parameter in different habitats
3.4 飽和光強下光合因子對比
由表4可見,溫度、濕度在林緣和林下兩種生境中均未見明顯差異,光強差異顯著(P<0.05),即林緣的光強高于林下的光強。表5中光合速率Pn、氣孔導(dǎo)度Cs和蒸騰速率Tr差異顯著(P<0.05),且林下的這三個指標均高于林緣。其余參數(shù)均未見明顯差異;兩種生境的光強均設(shè)置在了飽和點水平,結(jié)合光響應(yīng)曲線中的Ci的情況來看,兩種生境中Cs與Tr對象頭花Pn的影響更大。
3.5 植株長勢對比
對比林緣、林下象頭花植株的長勢(表6),可知林緣、林下的象頭花的株高、葉長、葉寬的呈顯著差異(P<0.05),而象頭花直徑未見顯著差異(P> 0.05)。同時林下象頭花株高、葉長、葉寬均大于林緣象頭花。林下株高、葉長、葉寬分別達到了(10.27±1.01) cm、(14.4±1.0) cm、(69.87±0.61) cm;而林緣分別為(7.93±3.93) cm、(11.6±0.52) cm、(8.43±0.06) cm。由此可見,處于林緣生境當中的象頭花長勢更好,這也與林下環(huán)境中象頭花的光合作用能力強有助于象頭花對于有機物的積累相對應(yīng)。
4 討論
在象頭花最適模型的篩選過程中,改進指數(shù)模型為林緣、林下兩種生境中象頭花光響應(yīng)曲線的最適模型。LSP反應(yīng)了植物Pmax的最大需求光照,根據(jù)最適模型擬合的參數(shù)可見,林緣的LSP為516.405 μmol·m-2·s-1,林下的LSP為638.500 μmol·m-2·s-1,這說明林下象頭花在達到最大光合速率時對光照的需求高于林緣的象頭花。而林下的LCP為8.142 μmol·m-2·s-1高于林緣的LCP為5.498 μmol·m-2·s-1,這說明林緣象頭花對環(huán)境的適應(yīng)性更強。研究表明植物的AQY范圍為0.03—0.07[34],而林緣的AQY為0.029 μmolCO2·μmol-1,林下的AQY為0.038 μmol CO2·μmol-1;林緣AQY處于最低值邊緣,林下的AQY也處于較低水平。因此,林下象頭花相對于林緣象頭花對光照的利用效率更高,但是從整體來說兩種生境中的象頭花對光照的利用效率均處于較低水平。其LSP、LCP、AQY均較低具有陰生植物的特性[35]。暗呼吸是植物新陳代謝重要的生理過程,其反應(yīng)了植物自身消耗情況;林緣的Rd為0.068 μmol·m-2·s-1明顯低于林下的Rd為0.159 μmol·m-2·s-1,林下象頭花的生理活性更強,并不利于有機物的積累。但是,林下象頭花的最大凈光合速率Pmax為6.740 μmol·m-2·s-1明顯高于林緣Pmax為4.290 μmol·m-2·s-1,那么林下象頭花的有機物的生產(chǎn)量也更多。這是因為不同生境中植物對資源的投資與分配會有所不同,這是植物與其生境長期適應(yīng)和進化(生境選擇)的結(jié)果[36]。

圖2 象頭花光合生理參數(shù)對不同光合有效輻射的響應(yīng) Figure2 Photosynthetic physiological factors of A. franchetianum Engl.response to different PARs

表4 不同生境的環(huán)境因子 Table4 Environment factor of in different habitats

表5 不同生境的凈光合速率參數(shù) Table5 Pn parameter in different habitats

表6 不同生境植株長勢 Table6 Growth potential of plant in different habitats
植物進行光合作用,或多或少的都因受到了外部環(huán)境的影響,研究表明影響植物光合作用的因子復(fù)雜且多變[37],而且這些因子對光合作用的影響不僅是綜合的,而且因子之間也會相互影響[8]。本實驗中,將兩種生境當中象頭花的凈光合速率與光合因子進行相關(guān)性分析,結(jié)果表明林緣象頭花的Pn與氣孔導(dǎo)度Cs、蒸騰速率Tr、空氣溫度Tair、和樣品室相對濕度RH-S、胞間CO2濃度Ci和樣品室CO2濃度CO2S均相關(guān)性極為顯著(P<0.01);而林下象頭花的Pn除與Cs、Tr、Tair、RH-S、Ci相關(guān)性極為顯著(P<0.01)外,還與PARO相關(guān)性極為顯著(P<0.01),這說明光照對林下象頭花的影響大于對林緣生境中象頭花的影響。結(jié)合兩種生境當中光合因子隨光照輻射的變化看來,在同等的光照條件下林下的象頭花葉片不管是Cs、Tr還是Ci均高于林緣生境中的象頭花;兩種生境當中象頭花葉片的水分利用效率WUE雖不相上下但是最終林下象頭花的光合作用能力更強。這與其同類植物半夏(Pinellia ternata)的生長特性研究結(jié)果一致[38]。
飽和光強是植物進行光合作用的理想光照條件,有研究指出生境和植物類型對氣孔響應(yīng)行為具有顯著影響[39],葉片對光照亦具有不同的適應(yīng)策略[40]。本實驗中,林緣、林下的自然環(huán)境因子除光強外溫度、濕度均未見明顯差異,僅林緣的光強高于林下的光強。在測量過程當中,將林緣、林下的象頭花葉片均置于飽和光強下,同時保障兩種生境空氣溫度Tair、樣品室CO2濃度CO2S、葉室外光合有效輻射PARO和胞間CO2濃度Ci均處于相似水平條件,即Tair、CO2S、PARO和Ci均未見明顯差異。研究發(fā)現(xiàn),林下的氣孔導(dǎo)度Cs和蒸騰速率Tr均高于林緣,那么林下的氣孔響應(yīng)更為積極,與之對應(yīng)的光合速率Pn也自然更大。象頭花對林下生境的適應(yīng),也造就了林下象頭花在相同的光照條件下其光合作用能力強于林緣象頭花,這與上文林緣、林下兩種生境光響應(yīng)曲線中凈光合速率的變化圖2E 相一致。林下高的Pn理因使得林下的象頭花植株長勢好于林緣的象頭花,從表6可見林下象頭花也的確實現(xiàn)了株高、葉長、葉寬均大于林緣象頭花。但象頭花為多年生草本,以塊莖入藥,是否高的地上莖、葉投入與地下塊莖的生長情況相匹配,還有待進一步探究。
綜上所述,5 種模型中改進指數(shù)模型是擬合兩種生境中象頭花光響應(yīng)曲線的最優(yōu)模型,更符合植物的生理學(xué)意義。Cs、Tr、Tair、RH-S、Ci等光合作用因子均與象頭花的Pn密切相關(guān)。林緣、林下兩種生境中的自然環(huán)境因子溫度、濕度未見明顯差異,僅林緣光照高于林下光照,但是由于象頭花葉片對林下生境的適應(yīng),林下象頭花葉片的光合作用能力強于林緣。林下象頭花的植株長勢也好于林緣。
本文只是對生長旺盛期林緣、林下兩種生境象頭花的光合光響應(yīng)特征進行了研究,為更加深入地了解象頭花的光合特性,還應(yīng)繼續(xù)開展更多生境中象頭花在不同生長期的光響應(yīng)、CO2響應(yīng)、日變化等響應(yīng)規(guī)律的研究,今后還需要做大量研究工作,以期為象頭花的生理生態(tài)研究以及開發(fā)利用提供依據(jù)。
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2020年10期)2020-11-26 08:24:50
數(shù)學(xué)物理學(xué)報(2020年2期)2020-06-02 11:29:24
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領(lǐng)巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
光學(xué)精密工程(2016年6期)2016-11-07 09:07:19
少兒科學(xué)周刊·兒童版(2015年10期)2015-11-07 03:42:03
核科學(xué)與工程(2015年4期)2015-09-26 11:59:03
