情緒記憶的神經機制探討
2019-07-03 03:12:40黃思淼
現代商貿工業 2019年10期
黃思淼
摘要:圍繞情緒記憶這一主題,綜合近些年的文獻以及研究成果全面論述了情緒記憶的神經機制。介紹了情緒記憶的理論模型中Bower的連結語義網絡模型,并對其優缺點進行了分析;以恐懼條件反射為研究范式,通過對杏仁核、內側顳葉、前額葉在情緒記憶中的作用討論了情緒記憶的特點和神經機制;說明了情緒記憶相關的激素、基因,并對個體差異的原因進行了解釋。回顧近幾年的研究成果,對情緒記憶機制的研究,情緒記憶機制和特點對于一些疾病治療的作用,以及其與科技結合的未來發展等方面做出了展望。
關鍵詞:情緒記憶;神經機制;個體差異
中圖分類號:F24文獻標識碼:Adoi:10.19311/j.cnki.1672-3198.2019.10.039
隨著人工智能的研究發展,計算機語言所編輯的智能程序不斷趨近于真正的人類大腦。然而,在人類記憶的機制中有一種很特別的方式——情緒記憶,這使得情緒性事件相對于中性事件更不容易被遺忘,從而不同于智能程序的記憶。
記憶根據記憶內容能否被有意識提取可以分為陳述性和非陳述性記憶,而情緒記憶是通過情緒因素對于記憶效率進行調節的一種記憶機制,主要體現在影響編碼、鞏固和提取等不同記憶加工階段(Dolcos,Labar,& Cabeza,2004a)。在情緒刺激的輔助下,記憶將更加強烈和持久。因此,情緒刺激的加工在大腦中享有優先權(Davidson,Maxwell,& Shackman,2004)。從進化的觀點看,情緒記憶作為重要的認知功能,對于生物生存和繁衍都具有重大影響。這就表明了情緒記憶在人類記憶中的顯著優勢地位,而其神經機制、相關激素和基因以及對于創傷后應激失調(Post-traumatic Stress Disorder,PTSD)等心理疾病的作用研究成為了當前研究的熱點。本文將通過對情緒記憶的理論模型、神經機制、激素研究、相關基因和個體差異方面進行全面的敘述。
1情緒記憶的理論模型
1.1Bower模型
連結語義網絡模型(Bowers Associative Semantic Network Model)(Bower,1981)——第一個情緒記憶模型在1981年由Bower提出。Bower情緒記憶連結語義網絡模型認為:各種基本情緒在記憶網絡中以結點的形式存在,而復雜情緒由這些基本情緒復合而成。基本情緒結點又與表情行為、自主反應模式等多個結點連接,因此當復雜情緒產生時,根據激活擴散原理,這些有關結點都會被激活。
Bower假定在特定情境下體驗到給定情緒時,這種情緒即會與事件一同存儲在記憶中,這種記憶表征模式包括兩個方面:一是心境一致性記憶(Mood-congruent Memory)和心境依賴性記憶(Mood-dependent Memory)。其中,心境一致性記憶是指人們在處理新信息的時候,對于那些情緒效價與當前心境一致的信息更加敏感,更容易被發現和深入加工——偏重于新信息;二是心境依賴性記憶是指在特定心境下編碼的信息,當再次處在同樣心境的時候更容易被提取——偏重于已存在的記憶。
1.2對Bower模型的分析
Bower的情緒記憶的連結語義網絡模型首次解釋了情緒記憶的存儲和表征方式,但仍有不足之處,比如某些記憶的提取并不會涉及情緒,或某些情緒的產生并不影響記憶本身;再比如按照Bower的情緒信息加工模型,某個情緒結點的觸發將會引起相關聯的所有結點激活,即當某情緒記憶被提取時必定引起相應的生理喚醒和主觀體驗,這是不符合實際的(王振宏,蔣長好,2008)。
2情緒記憶的神經機制
人們一般根據是否具有情緒維度和喚醒度,將記憶分為情緒記憶(Emotional Memory)和中性記憶(Neutral Memory)。情緒記憶在神經成像顯示中,除了相關感覺皮層外,內側顳葉、前額葉和杏仁核(Amygdala)也處于激活狀態(LaBar,2006),因此可以推測情緒記憶的優勢地位可能與內側顳葉、前額葉和杏仁核的相互作用有關。
2.1研究范式:恐懼條件反射
恐懼條件反射是情緒記憶的一部分,它經常被用于研究情緒記憶發生時大腦中的相關腦區活動。例如,Bechara等人在對腦損傷患者的對比實驗時發現:杏仁核損傷的患者雖然不能建立條件反射,但可以知道條件刺激伴隨著非條件刺激;海馬損傷的患者能建立條件反射,卻無法解釋原因;而杏仁核和海馬都損傷的患者,既無法建立條件反射,也不能解釋原因(Bechara,1995)。Bechara的實驗包括之后進行的一些動物研究都可以成為研究海馬、杏仁核等腦區功能的證據。
恐懼條件反射與人類生活中的恐懼和焦慮失調等心理疾病有密切聯系(Sotres-Bayon,Cain,LeDoux,2006)。當條件刺激多次單獨出現且未伴隨非條件刺激時,由條件刺激引起的條件反射將會逐漸減弱,這一過程被稱為條件反射中的消退(Extinction)階段。對消退階段的研究可能可以為減輕PTSD等情緒障礙提供思路。
2.2情緒記憶的特性
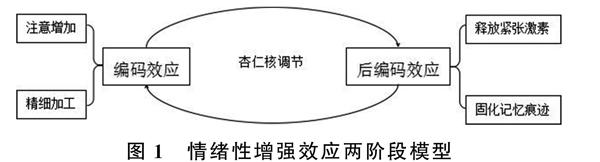
情緒記憶擁有兩個特性:一是情緒性增強效應(Emotion Enhancement Effect);二是可以進行自動加工。情緒性增強效應是指情緒性事件相比中性事件更容易被記住(Mori,Ikeda,Hirono,Kitagaki,Imamura,& Shimomura,1999)。Hamann(2001)曾根據其研究結果提出情緒增強效應的兩階段模型:編碼效應和后編碼效應,具體如圖1。
自動加工是指情緒記憶可以在意識下進行認知加工。意識上的情緒加工將外界信息通過丘腦—皮層—杏仁核通路進行傳遞(LeDoux,2003),這種皮層參與的加工方式比較復雜,導致速度也較慢。而意識下的情緒加工在皮層下進行傳遞,主要通路是丘腦和杏仁核的直接聯系,可以使人體對強烈的刺激做出最迅速的反應。并且在情緒增強效應的影響下,情緒性刺激使得情緒記憶加工的編碼階段注意資源分配增加,從而也促進了其自動加工。例如在注意分神實驗(Divided-attention Paradigm)中,同時給予被試高喚醒負性詞和聽覺辨別任務,然而被試在提取時仍能表現出喚醒詞成績優于中性詞和非喚醒情緒詞(Kensinger,Corkin,& Raichle,2004)。
2.3杏仁核與情緒記憶
大腦中與情緒記憶相關最重要的區域之一是杏仁核。杏仁核尤其在處理情緒性社會信號、建立情緒條件刺激和鞏固情緒記憶的過程中發揮關鍵作用(Dalgleish,2004)。并且在情緒加工的特性體現中,杏仁核也均扮演著不可或缺的角色。
杏仁核分為基底外側杏仁核(Basolateral Amygdala)和周邊附屬結構。其中,基底外側杏仁核分為外側核(Lateral Nuclei)、基底核(Basal Nuclei)和附屬基底核(Accessory Basal Nucle);周邊附屬結構分為中央核(Central Nuclei)、內側核(Medial Nuclei)和皮質核(Cortical Nuclei)。
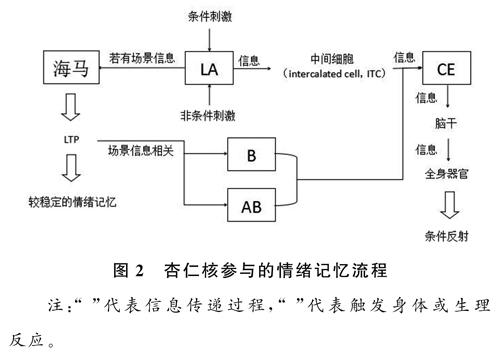
在條件反射建立的過程中,杏仁核的外側核(LA)、基底核(B)、附屬基底核(AB)和中央核(CE)起到了重要作用。LA具有較高的可塑性,是記憶中長時間增強效應(long-term potentiation,LTP)發生的主要區域。如果在條件反射建立的過程中附帶有場景信息,則會使LA在海馬誘發LTP,具體過程如圖2。
2.4其他腦區與情緒記憶
在Bechara(1995)的恐懼條件反射實驗中,已經發現海馬與情緒記憶有所關聯。在Richardson等(2004)的實驗中,使用fMRI檢測了左側海馬或杏仁核具有不同程度損傷的患者在進行詞匯編碼任務時的腦區活動,對比結果發現,在編碼階段,左側海馬的活性與左側杏仁核的損傷程度、杏仁核活性與左側海馬的損傷程度都有所關聯。Dolcos,Labar,& Cabeza(2004b)也在研究中發現,編碼階段杏仁核與內側顳葉的激活程度有強烈的關聯。
Dolcos等人(2004a)的另一研究表明,前額葉參與情緒記憶,且不同區域對應著不同情緒的認知,例如眶額葉與懲罰獎賞相關,背內側前額葉與情緒喚醒度有關等。Seigeriek(2006)的研究還顯示右背外側前額葉與非語言材料編碼相關,左背外側前額葉負責情緒和記憶的整合。
3情緒記憶的相關激素和基因
3.1情緒記憶相關激素
神經調節迅速、精準、持續時間短,但情緒記憶一般強烈而持久,可以發現人體中體液調節也具有類似的特點,因此在考慮情緒記憶的神經機制時,應考慮相關激素在其中的作用。杏仁核能夠調節腎上腺素(Epinephrine,E)和糖皮質激素(Glucocorticoids,G)等壓力相關激素,而杏仁核又是情緒記憶的“中樞”,因此腎上腺素和糖皮質激素與情緒記憶密不可分。實驗證明,這兩種激素都能促進記憶的鞏固。腎上腺素和糖皮質激素分別與其在大腦中對應的受體作用,來影響去甲腎上腺素的分泌,進而調節情緒記憶(McGaugh,2002)。
3.2情緒記憶相關基因
情緒記憶存在著多個方面的個體差異,這些差異的基礎可能是負責記憶編碼、鞏固和提取等功能的相關腦區及相關神經遞質受體的基因來決定的。近來的神經遺傳學實驗表明,兩種基因與情緒記憶存在一定聯系:一是腎上腺素ɑ2b受體基因(ADRA2B)對腎上腺素ɑ2b受體編碼,從而控制血管收縮和血壓調節(Muszkat,2005);腎上腺素ɑ2b受體作為自受體(Autoreceptors)抑制去甲腎上腺素的釋放,因此ADRA2B缺失會導致去甲腎上腺素增多,促進情緒記憶;實驗表明,ADRA2B缺失人群更易患PTSD(de Quervain,2007)。二是腦源性神經營養因子基因Val66Met(BDNFVal66Met),其編碼的腦源性神經營養因子(BDNF)可以調節突觸可塑性,影響學習和記憶(Bekinschtein,Cammarota,Lzquierdo,& Medina,2008)。
4情緒記憶的個體差異
4.1性別差異
大腦功能具有偏側化效應,例如大腦左半球主要負責語言,而右半球傾向著視覺和空間信息處理。在過往研究中,發現不同性別對于大腦半球的運用具有差異性,從而導致了男女在許多問題的處理上有不同的方式。同樣,在情緒記憶中也存在性別差異。
Canli等(2002)在研究中發現,編碼時杏仁核的激活程度主要體現為男性右側化優勢和女性左側化優勢。而因為左右腦功能的差異,有一種解釋為女性在情緒記憶的編碼過程中使用了大腦左半球的以語言為基礎的編碼機制,而男性更多使用右半球進行視覺空間信息處理,從而體現在左右側杏仁核激活的差異上。并且在另一研究中發現,情緒記憶的性別偏側化效應可發生于較長的延遲時間(2-3w)(王海寶,2009)。
4.2年齡差異
一般來說,隨著年齡的增長,大腦的認知記憶功能會發生一定的衰退。類似的,情緒記憶隨年齡的增長會出現偏向性。
Mather等人(2004;2005)在對不同年齡段人的行為學研究中,發現隨年齡的增加,對于正性和中性情緒圖片的再認準確度逐步高于負性情緒,而其進一步研究又表明老年人在觀看正性圖片時的杏仁核激活程度明顯高于負性圖片,年輕人則無此現象。文首提到情緒記憶在進化的觀點下看,是一種有利于生物生存繁衍的機制。而年齡差異正印證了這一點。“社會情緒選擇理論”(Socio-emotional Selective Theory,SST)認為隨年齡增長,人們認識到生存時間有限,因此具有更多調節情緒的動機偏向,來使情緒體驗最佳化。而由于情緒調節偏于正性情緒,使對正性情緒的注意資源分配增加,從而形成了情緒記憶的年齡差異。
5結論
本文介紹了情緒記憶的理論模型中Bower的連結語義網絡模型,并對其優缺點進行了分析;此外以恐懼條件反射為研究范式,通過對杏仁核、內側顳葉、前額葉在情緒記憶中的作用討論了情緒記憶的特點和神經機制;最后說明了情緒記憶相關的激素、基因,并對個體差異的原因進行了解釋。
盡管近幾年在情緒記憶方面的研究有很大進展,但在其研究領域中,仍有許多問題和方向有待探索,如杏仁核與其他腦區的相互作用對于情緒記憶的影響,情緒記憶的增強機制對于PTSD、抑郁癥等疾病的作用與可從情緒記憶的損害機制入手的治療方案,情緒記憶機制在人工智能方面的應用等。
參考文獻
[1]Bechara,A.,Tranel,D.,Damasio,H.,Adolphs,R.,& Rockland,C.Double dissociation of conditioning and declarative knowledge relative to the amygdala and hippocampus in humans[J].Science,1995,(269):1115-1118.
[2]Bekinschtein,P.,Cammarota,M.,Izquierdo,I.,& Medina,J.H.Bdnf and memory formation and storage.Neuroscientist A Review[J].Journal Bringing Neurobiology Neurology & Psychiatry,2008,(14):147.
[3]Bower,G.H.Mood and memory[J].Am Psychol,1981,(36): 129-148.
[4]Canli,T.,Desmond,J.E.,Zhao,Z.,& Gabrieli,J.D.E.Sex differences in the neural basis of emotional memories[C].Proceedings of the National Academy of Sciences of the United States of America,2002,(99):10789-10794.
[5]Dalgleish,T.The emotional brain[J].Nature Reiview Neuroscience,2004,(5):582-589.
[6]Davidson,R.J.,Maxwell,J.S.,& Shackman,A.J.The privileged status of emotion in the brain[C].Proceedings of the National Academy of Sciences of the United States of America,2004,(101):11915-11916.
[7]de Quervain,D.J.,Kolassa,I.T.,Ertl,V.,Onyut,P.L.,Neuner,F.,& Elbert,T.,et al.A deletion variant of the alpha2b-adrenoceptor is related to emotional memory in europeans and africans[J].Nature Neuroscience,2007,(10):1137-1139.
[8]Dolcos,F.,Labar,K.S.,& Cabeza,R.Dissociable effects of arousal and valence on prefrontal activity indexing emotional evaluation and subsequent memory:an event-related fmri study[J].Neuroimage,2004,(23):64-74.
[9]Dolcos,F.,Labar,K.S.,& Cabeza,R.Interaction between the amygdala and the medial temporal lobe memory system predicts better memory for emotional events[J].Neuron,2004,(42):855-863.
[10]Hamann,S.Cognitive and neural mechanisms of emotional memory[J].Trends in Cognitive Sciences,2001,(5):394-400.
[11]Kensinger,E.A.,Corkin,S.,& Raichle,M.E.Two routes to emotional memory:distinct neural processes for valence and arousal[C].Proceedings of the National Academy of Sciences of the United States of America,2004,(101):3310-3315.
[12]Labar,K.S.,& Cabeza,R.Cognitive neuroscience of emotional memory[J].Nature Reviews Neuroscience,2006,(7):54-64.
[13]Ledoux,J.The emotional brain,fear,and the amygdala[J].Cellular & Molecular Neurobiology,2003,(23):727-738.
[14]Mather,M.,Canli,T.,English,T.,Whitfield,S.,Wais,P.,& Ochsner,K.,et al.Amygdala responses to emotionally valenced stimuli in older and younger adults[J].Psychological Science,2004,(15):259-263.
[15]Mather,M.,& Carstensen,L.L.Aging and motivated cognition:the positivity effect in attention and memory[J].Trends in Cognitive Sciences,2005,(9):496-502.
[16]Mcgaugh,J.L.,& Roozendaal,B.Role of adrenal stress hormones in forming lasting memories in the brain[J].Current Opinion in Neurobiology,2002,(12):205-210.
[17]Mori,E.,Ikeda,M.,Hirono,N.,Kitagaki,H.,Imamura,T.,& Shimomura,T.Amygdalar volume and emotional memory in alzheimer's disease[J].Am J Psychiatry,1999,(156):216-222.
[18]Muszkat,M.,Kurnik,D.,Solus,J.,Sofowora,G.G.,Xie,H.G.,& Jiang,L.,et al.Variation in the alpha2b-adrenergic receptor gene (adra2b) and its relationship to vascular response in vivo[J].Pharmacogenet Genomics,2005,(15):407-414.
[19]Richardson,M.P.,Strange,B.A.,& Dolan,R.J.Encoding of emotional memories depends on amygdala and hippocampus and their interactions[J].Nature Neuroscience,2004,(7):278-285.
[20]Sergerie,K.,Lepage,M.,& Armony,J.L.A process-specific functional dissociation of the amygdala in emotional memory[J].J Cogn Neurosci,2006,(18):1359-1367.
[21]Sotresbayon,F.,Cain,C.K.,& Ledoux,J.E.Brain mechanisms of fear extinction:historical perspectives on the contribution of prefrontal cortex[J].Biological Psychiatry,2006,(60):329-336.
[22]王海寶.情緒記憶特異性認知神經機制[D].北京:中國科學技術大學,2009.
[23]王振宏,蔣長好.情緒記憶的理論模型[J].西北師范大學學報(社會科學版),2008,(45):112-117.
