超聲波輔助副干酪乳桿菌發酵脫脂乳粉制備多肽的動力學研究
2019-07-10 13:10:06陳蘇婉湯穎秀侯小珊何榮海馬海樂
食品工業科技 2019年11期
陳蘇婉,湯穎秀,邢 政,侯小珊,何榮海,馬海樂
(江蘇大學食品與生物工程學院,江蘇省農產品物理加工重點實驗室,江蘇鎮江 212013)
近年來,副干酪乳桿菌作為乳制品的發酵菌種被廣泛用于發酵生產,傳統副干酪乳桿菌發酵乳的功效成分主要是益生菌,其功能單一,對健康的促進效果有限[1]。傳統副干酪乳桿菌發酵乳發酵過程存在發酵周期長、菌體內外物質交換效率低等問題,導致發酵效率及產物得率都較低[2-4]。通過前期研究發現,以脫脂乳粉、葡萄糖等為底物,在微生物的發酵過程中施加適宜的超聲波處理,可有效提高發酵效率,加快發酵進程,提高發酵產物產量和功能特性[5];超聲波處理后的發酵乳中多肽含量顯著提高,同時,發酵乳的營養品質和活性功能得到了加強[5],但超聲波對脫脂乳粉發酵進程促進的原因還有待進一步的研究。
發酵動力學是指在整個發酵過程中,隨著發酵時間的延長,發酵液中微生物的活菌數、產物的生成量和底物的消耗量的變化規律以及各變量之間通過軟件建立的相應數學模型關系[6]。研究發酵動力學是為了了解發酵過程中不同變量之間的關系,找出各種變化和現象之間存在的潛在聯系,對不同參數進行非線性擬合確定相應模型參數的系數值[6]。在微生物發酵的過程中,微生物的生長代謝會影響發酵產物中產物的生成以及底物的消耗,微生物消耗的底物用于自身生長的維持和生產活性產物[7]。研究發酵動力學對于發酵過程的優化和調控、深入了解微生物的代謝規律、將發酵規模從實驗室向工廠化的轉變和放大有著十分重要的意義[8]。何榮海等[9]研究了枯草芽孢桿菌液態分批發酵菜籽粕的代謝特征,發現多肽濃度在32 h時達到穩定,對枯草芽孢桿菌正常液態發酵過程中的菌體生長規律、多肽生成和底物蛋白消耗進行了模型擬合,且模型驗證的計算值與實測值的相對誤差低于10%,較好地反映了枯草芽孢桿菌分批發酵過程的動力學特征。孟掉琴等[10]通過篩選復合益生菌,以蘋果濁汁為原料,構建了菌體生長、產物生成及底物消耗動力學模型,研究確立了模型的動力學參數,并進行驗證,發現模型的理論值與試驗值誤差均小于10%,擬合較好,建立的動力學模型能夠較好地預測復合益生菌發酵蘋果濁汁發酵過程的變化。對超聲波輔助發酵制備發酵乳過程進行發酵動力學研究,有助于對超聲處理促進發酵過程原因的分析。
為此,本文對超聲處理與未超聲處理的副干酪乳桿菌發酵脫脂乳制備多肽發酵過程分別建立發酵動力學模型,并進行比較分析,探討超聲波提高發酵效率的原因。
1 材料與方法
1.1 材料與儀器
副干酪乳桿菌(CICC編號:20241) 中國工業微生物菌種保藏管理中心;脫脂乳粉(蛋白質含量:32.9%) 新西蘭恒天然公司;食用葡萄糖 呼倫貝爾東北阜豐生物科技有限公司;MRS培養基 杭州百思生物技術有限公司;酪蛋白(AR)、1,1-二苯基-2-三硝基苯肼(DPPH,AR)、中性紅(AR)、百里酚酞(AR) 國藥集團化學試劑有限公司。
PB-10型pH計 賽多利斯科學儀器(北京)有限公司;T6新世紀紫外分光光度計 北京普析通用儀器有限責任公司;DL-5C型離心機 上海安亭科學儀器廠;YX280A型滅菌鍋 上海三申醫療器械有限公司;QYC-200型全溫空氣搖床 上海福瑪實驗設備有限公司;JJ-CJ-1FD型單人單面無菌超凈操作臺 蘇州市金凈凈化設備科技有限公司;脈沖多頻超聲試驗設備 江蘇大學食品與生物工程學院自主研制。
1.2 實驗方法
1.2.1 多肽發酵乳的制備 稱取脫脂乳粉12.6 g,葡萄糖6 g,100 mL蒸餾水溶解至250 mL錐形瓶中,攪拌至完全溶解,115 ℃高壓滅菌15 min后冷卻至室溫,以5%(v/v)的接種比例接種副干酪乳桿菌種子液,使用搖床進行發酵(發酵時間:24 h,搖床轉速:180 r/min,發酵溫度:37 ℃)。在發酵9 h時取出發酵液,在37 ℃用頻率為28 kHz,功率為100 W/L,每超聲100 s間歇10 s超聲35 min[11]。在超聲組進行超聲的同時,將對照組置于37 ℃水浴鍋中保溫,待超聲結束后將超聲組與對照組一同放回搖床中繼續培養。在發酵過程的不同時刻取樣,分別測定pH、葡萄糖含量、游離氨基酸含量、活菌數、多肽含量以及底物剩余蛋白質含量等指標。
每隔2 h取超聲組與對照組的發酵乳分別測定活菌總數和pH。另外取發酵乳按如下步驟制備樣液,用于測定除活菌數之外的指標:將發酵乳振蕩搖勻后,取10 mL置于50 mL離心管中,用1 mol/L HCl溶液調節發酵乳pH至3.4~3.6,4000 r/min離心20 min。取上清液用1 mol/L NaOH溶液調節pH至8.2~8.4,4000 r/min離心20 min獲得上清液進行相關指標的測定。
1.2.2 指標的測定
1.2.2.1 酸含量的變化 由于檢測發酵乳中乳酸的含量較為不便,故以pH的變化代替乳酸含量的變化作為動力學模型研究的監測指標,其中pH用pH計直接測定。
1.2.2.2 葡萄糖含量的測定 取1.2.1中的發酵待測樣液參照國標法[12]測定發酵乳中葡萄糖的含量。取5.0 g混勻后的發酵乳(精確至0.001 g),置于250 mL容量瓶中,加入50 mL水,緩慢加入乙酸鋅溶液5 mL和亞鐵氰化鉀溶液5 mL,加水至刻度,混勻,靜置30 min,用干燥濾紙過濾,棄去初濾液,取后續濾液備用。吸取堿性酒石酸銅甲液5.0 mL和堿性酒石酸銅乙液5.0 mL于250 mL錐形瓶中,加水10 mL,玻璃珠2~4粒,控制在2 min內加熱至沸,保持沸騰繼續以1滴/2 s的速度滴定,直至藍色剛好褪去即為終點,記錄樣液消耗體積,平行測定3次,得出平均消耗體積(V)。
X=(m1×25)/(m×V)
式(1)
式中:X:試樣中還原糖的含量(g/100 g);m1:堿性酒石酸銅溶液(甲、乙液各半)相當于某種還原糖的質量(mg);m:試樣質量(g);V:測定時平均消耗試樣溶液體積(mL)。
1.2.2.3 發酵乳中活菌數的測定 按照GB 4789.2-2016食品微生物學檢驗菌落總數的測定方法對發酵乳中的活菌數進行菌落計數[13]。將發酵乳搖勻,取5 mL于50 mL已滅菌的生理鹽水中,混勻,制成樣品溶液。取1 mL上述樣品溶液于裝有9 mL無菌生理鹽水的無菌試管中,稀釋6倍,取1 mL稀釋液于滅菌平板中,將MRS瓊脂培養基傾注于平板內,凝固后轉入37 ℃培養箱中培養48 h進行菌落計數[5]。
1.2.2.4 游離氨基酸含量的測定 取超聲組與對照組發酵待測樣液各5 mL分別于兩個250 mL錐形瓶中,每個錐形瓶中分別加入25 mL蒸餾水,一個錐形瓶中加入2~3滴0.1%中性紅的50%乙醇溶液指示劑(中性紅指示劑),用0.1 mol/L NaOH滴定至pH為7.4(顏色由紅色變為琥珀色),記錄消耗NaOH的體積為V1;向另一個錐形瓶中加入10 mL 40%甲醛溶液、2~3滴0.1%百里酚酞乙醇指示劑,充分搖勻后靜置1 min,用0.1 mol/L NaOH滴定至pH為10.0(顏色由無色變為淺藍色),記錄消耗NaOH的體積為V2。按下式計算出樣品中氨基酸態氮濃度,即可作為發酵乳中游離氨基酸的含量[14-16]。
氨基酸態氮濃度(mg/mL)=C×(V2-V1)×0.014×100
式(2)
式中:C:氫氧化鈉標準溶液的濃度(mol/L);V1:中性紅為指示劑消耗氫氧化鈉標準溶液的體積(mL);V2:百里酚酞為指示劑消耗氫氧化鈉標準溶液的體積(mL);0.014:氮的毫摩爾質量(g/mmoL);100:氨基酸態氮濃度轉換系數。
1.2.2.5 多肽含量的測定 準確稱取酪蛋白標準品12 g,用蒸餾水配制成12 mg/mL的母液,分別稀釋為10、8、6、4和2 mg/mL的酪蛋白標準溶液。取1 mL酪蛋白標準溶液與3 mL雙縮脲試劑混勻后靜置30 min,于540 nm處測定吸光度值[4],根據結果制作標準曲線,標準曲線方程為y=0.0629x+0.0022,R2=0.998。
參照任國譜等[17]的研究方法配制雙縮脲試劑。將5 mL按照1.2.1中方法制備得到的發酵液待測樣液與10%的三氯乙酸(TCA)溶液1∶1等體積混勻后靜置10 min,4000 r/min離心15 min。將離心上清液與雙縮脲試劑以體積比3∶2的比例混勻后靜置15 min,于540 nm處測定吸光度OD值,每個樣品平行測定三次,取平均值,通過標準曲線回歸方程換算得到多肽濃度C[18]。按照同樣方法測定制備的發酵0 h時的發酵樣液中多肽濃度C0。多肽含量的計算見下式:
多肽含量(mg/mL)=(C-C0)×2×5/3
式(3)
式中:C:發酵到某一時刻時發酵樣液多肽濃度(mg/mL);C0:起始多肽濃度(mg/mL);2、5/3:樣品的稀釋倍數。
1.2.2.6 底物剩余蛋白質含量計算 在發酵過程中,副干酪乳桿菌生長所消耗的蛋白質絕大部分都轉變成了牛乳多肽和游離氨基酸,因此,發酵乳中的底物剩余蛋白質含量可以通過脫脂乳粉發酵培養基中的總蛋白量除去乳源多肽和游離氨基酸的含量來計算。底物剩余蛋白質含量按下式計算:
M=M總-M1-M2
式(4)
式中:M總:發酵液中總蛋白含量(mg/mL);M1:發酵液中多肽含量(mg/mL);M2:發酵液中游離氨基酸含量(mg/mL)。
1.2.3 菌體生長動力學模型建立及擬合曲線分析 在菌體生長動力學中,Logistic模型能夠很好地反映菌體濃度增加對菌體自身生長的抑制,表明微生物的生長速率不依賴物質的限制[19]。本文采用該模型來描述副干酪乳桿菌發酵制備多肽發酵乳中菌體細胞的生長,式(5)為Logistic方程。
式(5)
令t=0,X=X0,Logistic方程可以積分為代數方程式(6)。
式(6)
式中:X:發酵乳中的活菌數(106CFU/mL);μm:最大比生長速率(106個/h);Xmax:活菌數最大值(106CFU/mL)。
對超聲組和未超聲組的活菌數增長情況分別建立動力學模型分析。以式(6)為自定義函數,應用Matlab軟件進行非線性擬合并進行對比分析。
1.2.4 產物生成動力學模型建立及擬合曲線分析 Luedeking和Piret對產物生成動力學規律進行了總結,可描述發酵產物形成與細胞生長的關系[20]。
式(7)
將式(6)代入(7),并積分可得式(8):
式(8)
式中:P:多肽濃度(mg/mL);P0:初始多肽濃度(mg/mL);A:與菌體生長相關聯的產物合成常數(g/g);β:與菌體量相關聯的產物合成常數(g/g·h);X:發酵乳中的活菌數(106CFU/mL)。
對超聲組和未超聲組的多肽濃度變化情況分別建立動力學模型分析。以式(8)為自定義函數,應用Matlab軟件進行非線性擬合并進行對比分析。
1.2.5 底物消耗動力學模型建立及擬合曲線分析 在副干酪乳桿菌發酵產多肽的過程中,發酵培養基中奶粉蛋白的消耗主要用于菌株的生長、多肽和游離氨基酸的生成,乳源多肽和氨基酸的生成是由于菌株生長過程中釋放的酶作用于牛乳蛋白而生成,可以看作菌株生長的一部分,因此牛乳蛋白主要是用于菌株的生長和維持,故牛乳蛋白的消耗可用與Luedeking-Piret方程相似的方程式9表示。
式(9)
對式(9)進行求解得到積分式(10)。
式(10)
式中:S:底物蛋白剩余量(×100 mg);S0:初始底物蛋白剩余量(×100 mg);Yx/s:底物用于菌體生長的得率常數;ms:微生物維持常數。
對超聲組和未超聲組的底物蛋白剩余量分別建立動力學模型分析以式(10)為自定義函數,應用Matlab軟件進行非線性擬合并進行對比分析。
1.3 數據分析
每個實驗結果重復測定3次,結果以“平均值±標準差”的形式表示。使用SPSS 17.0進行單因素方差分析,顯著性水平α設定值為0.05,使用Origin 8.6進行圖形繪制。
2 結果與分析
2.1 副干酪乳桿菌發酵乳發酵過程主要指標的變化及分析
超聲組和對照組的pH基本保持一致(圖1),超聲組的葡萄糖消耗量略低于未超聲組(圖2),發酵乳中的游離氨基酸含量略高于未超聲組,且游離氨基酸含量在發酵20 h以后增加幅度變大(圖3)。如圖4、圖5,在發酵9 h對發酵乳進行超聲波處理后,隨著發酵時間的延長,超聲組的多肽含量和活菌數逐漸高于對照組。超聲組發酵底物中的剩余蛋白質含量要低于對照組(圖6)。

圖1 發酵過程中pH變化曲線圖

圖3 發酵過程中游離氨基酸含量變化曲線圖

圖4 發酵乳中菌體濃度變化曲線圖

圖5 發酵過程中多肽含量變化曲線圖

圖6 發酵過程中底物剩余蛋白質含量變化曲線圖
2.2 菌體生長動力學模型參數求解及擬合曲線分析
從圖4菌體濃度變化曲線可以發現超聲組與未超聲組菌體生長都呈S型曲線,符合Logistic模型反映的分批發酵過程中菌體的生長規律[20],超聲組發酵液中的活菌數高于未超聲組,以式(6)為自定義函數,應用Matlab軟件對未超聲組和超聲組副干酪乳桿菌數實驗值與模型方程分別進行非線性擬合。
利用Matlab軟件擬合獲得未超聲組副干酪乳桿菌生長實驗值與模型方程擬合曲線(圖7)、發酵乳中活菌數(X)隨時間變化的函數方程式(11)。

圖7 未超聲組副干酪乳桿菌生長實驗值與模型方程擬合曲線
式(11)
式(11)模型參數:X0=6.745,Xmax=48.6,μm=0.297,R2=0.9900。從圖8可以發現,式(11)擬合方程的曲線與實驗數據能較好地吻合。
利用Matlab軟件擬合獲得超聲組副干酪乳桿菌生長實驗值與模型方程擬合曲線(圖8)、發酵乳中活菌數(X)隨時間變化的函數方程(式12)。

圖8 超聲組副干酪乳桿菌生長實驗值與模型方程擬合曲線

式(12)
式(12)模型參數:X0=5.914,Xmax=55.503,μm=0.318,R2=0.9926。從圖8可以發現,式(12)擬合方程的曲線與實驗數據能較好地吻合,因此該方程較好地描述了超聲組副干酪乳桿菌發酵制備多肽過程中菌體副干酪乳桿菌的生長規律,為優化控制提供了參考依據。
比較超聲組和未超聲組的動力學模型可以發現,超聲組的最大活菌數高于未超聲組,這與圖4發酵乳中菌液濃度變化曲線的變化規律一致,超聲組的最大比生長速率高于未超聲組,由此可以解釋隨著發酵時間的延長,超聲組發酵液中的活菌數逐漸高于未超聲組的原因。
2.3 產物生成動力學模型參數求解及擬合曲線分析
從圖5超聲組和未超聲組的多肽含量變化曲線可以發現超聲組發酵液中的多肽生成量要高于未超聲組。以式(8)為自定義函數,應用Matlab軟件對未超聲組和超聲組發酵乳多肽濃度的實驗值和模型方程分別進行非線性擬合。
利用Matlab軟件擬合獲得未超聲組發酵乳多肽濃度實驗值與模型方程擬合曲線(圖9)、多肽濃度(P)隨時間變化的函數方程(式13)和模型參數:P0=0.0301,α=0.01312,β=-0.0005245,R2=0.9658。

圖9 未超聲發酵乳多肽濃度實驗值與模型方程擬合曲線
式(13)
利用Matlab軟件擬合獲得超聲組發酵乳多肽濃度實驗值與模型方程擬合曲線(圖10)、多肽濃度(P)隨時間變化的函數方程(式14)。

圖10 超聲發酵乳多肽生成動力學與模型方程擬合曲線
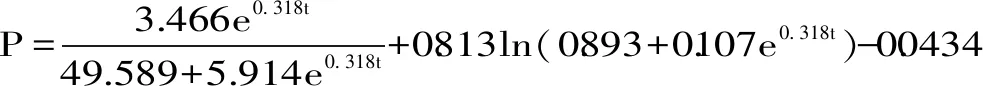
式(14)
式(14)模型參數:P0=0.0191,α=0.01056,β=-0.001047,R2=0.9658。從圖10可知,式(14)擬合方程的曲線與試驗所得數據在發酵24 h內能很好地擬合,較好地描述了超聲發酵乳發酵過程中乳源多肽的生成情況。
β為負值,說明副干酪乳桿菌發酵產生的多肽隨著菌體的生長,含量逐漸減少,分析其原因可能是由于副干酪乳桿菌分泌的蛋白酶將牛乳多肽進一步水解成游離氨基酸,導致多肽含量降低,比較超聲發酵乳和未超聲發酵乳的擬合方程可以發現,超聲發酵乳的β值(即與菌體量相關聯的產物合成常數)大于未超聲組的β值,說明超聲可以加快發酵過程中的物質代謝,使得發酵液中多肽含量增幅減緩,進一步導致游離氨基酸含量增多,最終產生的結果就是在發酵22 h以后,發酵乳的多肽含量趨于穩定甚至出現下降趨勢,同時,這一結論與圖3發酵液中游離氨基酸含量變化曲線也相對應。
2.4 底物消耗動力學模型參數求解及擬合曲線分析
從圖6發酵過程中底物剩余蛋白質變化曲線可以發現未超聲組的底物蛋白剩余含量高于超聲組。以式(10)為自定義函數,應用Matlab軟件對未超聲組和超聲組發酵乳底物蛋白消耗的實驗值與模型方程分別進行非線性擬合。
利用Matlab軟件擬合獲得未超聲組發酵乳底物蛋白消耗實驗值與模型方程擬合曲線(圖12)、底物蛋白消耗量(S)隨時間變化的函數多肽濃度(P)隨時間變化的函數方程式(15)和模型參數:S0=37.56,Yx/s=163.1,ms=-0.001451,R2=0.9778。

圖11 未超聲發酵乳底物蛋白消耗實驗值與模型方程擬合曲線

式(15)
利用Matlab軟件擬合獲得超聲組發酵乳底物蛋白消耗實驗值與模型方程擬合曲線(圖12)、底物蛋白消耗量(S)隨時間變化的函數多肽濃度(P)隨時間變化的函數方程式(16)和模型參數:S0=37.58,Yx/s=187.2,ms=-0.001908,R2=0.9836。
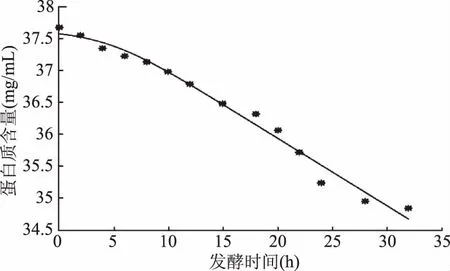
圖12 超聲發酵乳底物蛋白消耗實驗值與模型方程擬合曲線

式(16)
從圖12可知,式(15)擬合方程的曲線與試驗所得數據能很好地擬合,與圖6也相吻合,較好地描述了超聲發酵乳中底物蛋白的消耗情況。
比較未超聲和超聲發酵乳底物蛋白的消耗情況可以發現,超聲組和未超聲組的初始底物蛋白剩余量基本是相同的,但隨著發酵時間延長,超聲發酵乳的底物剩余蛋白含量逐漸小于未超聲組,同時,超聲組底物用于菌體生長的得率常數Yx/s大于未超聲組,說明超聲可以加快發酵乳中牛乳蛋白的消耗,提高發酵效率,有利于發酵過程的進行。
2.5 發酵乳動力學模型分析
根據試驗數據,通過Matlab軟件模擬未超聲組與超聲組副干酪乳桿菌發酵制備多肽發酵乳的動力學模型參數值如表1所示。表1表明,兩種發酵過程菌體生長均符合Logistic模型,其中超聲組模型中的最大比生長速率Xmax比未超聲組提高14.2%;兩種發酵過程的產物生成均符合Luedeking和Piret提出的動力學模型,其中超聲組模型中與菌體量相關聯的產物合成常數β比未超聲組提高99.62%;兩種發酵過程的底物消耗均符合Luedeking-Piret模型,其中超聲組模型中底物用于菌體生長的得率常數Yx/s比未超聲組提高14.78%。

表1 動力學模型參數
3 結論
本文建立了超聲波輔助發酵和無超聲發酵過程的菌體生長、產物生成和底物消耗動力學模型,對比超聲處理和未超聲處理的發酵動力學模型,發現兩種發酵過程均符合Logistic模型規律,且超聲組最大比生長速率Xmax為55.5,未超聲組最大比生長速率Xmax為48.6,超聲組的最大比生長速率比未超聲組提高14.2%;兩種發酵過程的產物合成生成均符合Luedeking和Piret提出的動力學模型,且超聲組產物合成常數β為-0.001047,未超聲組產物合成常數β為-0.0005245,超聲組產物合成常數比未超聲組提高99.62%;兩種發酵過程的底物消耗均符合Luedeking-Piret模型,且超聲組模型中底物用于菌體生長的得率常數Yx/s為187.2,未超聲組Yx/s為163.1,超聲組的菌體生長得率常數比未超聲組提高14.78%。因而在發酵乳的發酵過程中使用超聲波處理可以加快發酵過程中的物質代謝,提高發酵效率,利于發酵過程的進行,在今后的研究中可利用本文的分析結果研究發酵過程中產物的生成和底物的消耗變化規律,同時可對副干酪乳桿菌發酵制備多肽發酵乳的發酵過程變化規律進行監控和預測。
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
小讀者(2021年2期)2021-03-29 05:03:48
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
數學物理學報(2020年2期)2020-06-02 11:29:24
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
光學精密工程(2016年6期)2016-11-07 09:07:19
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
