沙棘7個亞種與26個重要品種的遺傳多樣性
2019-07-24 08:19:18王羅云何彩云羅紅梅張建國段愛國曾艷飛
浙江農林大學學報 2019年4期
王羅云,何彩云,羅紅梅,張建國,段愛國,曾艷飛
(1.中國林業科學研究院 林業研究所 國家林業和草原局林木培育重點實驗室,北京100091;2.中國林業科學研究院沙漠林業實驗中心,內蒙古 磴口015200)
沙棘Hippophae rhamnoides別名醋柳、沙棗,為胡頹子科Elaeagnaceae沙棘屬Hippophae多年生落葉灌木或小喬木。根據SUN等[1]和BARTISH等[2]的系統分類,沙棘屬共有7個種,11個亞種;其中有9個亞種廣泛分布于亞歐溫帶地區。沙棘果實中維生素C和黃酮含量較高,種子中富含脂肪油,枝葉富含高蛋白,這些生物活性物質是美容產品、抗敗血酸藥物和營養補充劑等的重要組成成分[3];沙棘的根系能產生根瘤,具有固氮功能,從而可達到改善土壤的作用;又由于其能適應干旱的環境,可以防風固沙,因此可用于生態恢復和土壤保護。因此沙棘是集生態、經濟和社會效益為一體的多用途樹種[3-4]。中國是沙棘屬植物類群分布最多的國家,包括7個種,5個亞種;其沙棘屬的資源蘊藏量也最大,沙棘天然林分布達67.54萬hm2[4];而中國的青藏高原地區被認為是沙棘的起源中心[5]。目前,中國沙棘種植總面積達200.00萬hm2,約占世界總面積的90%以上[6]。但當前廣泛用于栽培的大果沙棘品種主要是從俄羅斯引進的,人工選育自蒙古沙棘H.rhamnoidessubsp.mongolica亞種的天然遠緣雜交種[7]。中國沙棘自然資源的開發和利用對其人工培育有重要意義;而對沙棘遺傳多樣性的系統研究是沙棘新品種選育的基礎之一。目前,已有利用分子標記研究沙棘屬植物的系統發育關系的報道[1,8],探討了沙棘種下不同亞種的起源過程[5],分析了沙棘個別亞種,如中國沙棘H.rhamnoidessubsp.sinensis,云南沙棘H.rhamnoidessubsp.yunnanensis的遺傳多樣性,但是對于沙棘種下多個亞種的多樣性及分化的研究很少。本研究利用簡單序列重復(SSR)標記比較分析了沙棘中分布最廣的7個亞種的遺傳多樣性和遺傳分化,分析了當前廣泛栽培的26個大果沙棘品種的遺傳多樣性和可能來源,以期為沙棘新品種選育提供基礎。
1 材料與方法
1.1 分析材料和DNA提取
從野外采集中國沙棘,云南沙棘,中亞沙棘H.rhamnoidessubsp.turkestanica,蒙古沙棘,高加索沙棘H.rhamnoidessubsp.caucasica,海濱沙棘H.rhamnoidessubsp.rhamnoides和溪生沙棘H.rhamnoidessubsp.fluviatilis共7個沙棘亞種葉片樣品,其中:蒙古沙棘20個個體,其他沙棘亞種各10個個體;收集在中國林業科學研究院溫室播種的高加索沙棘幼苗10株;從黑龍江農業科學院漿果研究所、中國林業科學研究院沙漠實驗中心收集26個大果沙棘品種的葉片。樣品采集見表1。葉片采集后用變色硅膠干燥處理帶回實驗室,采用植物全基因組DNA提取試劑盒(北京天根生物公司)提取總DNA。

表1 7個沙棘亞種與26個大果沙棘品種采樣信息Table 1 Sampling information of the 7 subspecies and 26 cultivars of Hippophae rhamnoides
1.2 微衛星標記分析
所用的14個微衛星標記選自李珊珊等[9]從沙棘轉錄組和基因組中開發的,引物信息見表2。為簡化實驗,PCR擴增采用將2~4對攜帶不同熒光標記(Fam,Hex,Rox和Tamra,生工生物工程股份有限公司,上海),且擴增產物長度不同的微衛星引物混合后,在同一PCR管中進行。擴增反應體系為30 μL,分別包括2×T5 Super PCR Mix(PAGE)(擎科新業生物技術有限公司,北京),正向和反向引物分別0.2 μmol·L-1和模板 DNA約 50 ng。熱循環反應在 ABI System 9700PCR擴增儀(Applied Biosystems,Life Technologies)上進行,程序如下:94℃預變性4 min,10個循環的94℃變性30 s,60℃(每個循環降1℃)退火30 s和72℃延伸45 s,20個循環的94℃變性30 s,50℃退火30 s和72℃延伸45 s,最后72℃10 min。
PCR產物交由擎科新業生物技術有限公司進行變性[0.5 μL產物與9.0 μL的甲酰胺,0.5 μL的內標Liz500(Applied Biosystems,Life Technologies)混合后93℃變性5 min],然后在ABI 3730測序儀上進行毛細管電泳。最后利用Genemapper 4.0軟件(Applied Biosystems,Life Technologies)讀取電泳結果并分析得到每個個體基因型。每一批數據從PCR到毛細管電泳,都以大果沙棘品種 ‘向陽’為陽性對照。
1.3 遺傳多樣性和分化分析
采用軟件GenALEX 6.503[10],針對每個沙棘亞種和所有品種,計算如下遺傳多樣性指數:等位基因個數(NA), 有效等位基因個數(Ne), Shannon 信息指數(I), 觀測雜合度(HO), 期望雜合度(HE)和固定指數(F)。 利用微衛星分析軟件 MSA 4.05[11]計算亞種兩兩之間的遺傳分化系數(FST)[12], 并進行 10 000次重抽樣后統計顯著性,檢驗各亞種間分化是否顯著偏離于0。

表2 14個微衛星位點基本信息Table 2 Information of the 14 microsatellite loci
1.4 系統發育分析
基于個體多位點微衛星基因型,利用MSA軟件[11]計算個體之間的遺傳距離(DC)[13];然后基于此遺傳距離,采用PHYLIP 3.695[14]軟件包中的NEIGHBOR子程序構建系統發育樹,自展值設為500次。
1.5 貝葉斯聚類
利用貝葉斯聚類軟件STRUCTURE 2.3.4[15]確定沙棘各亞種的分組,并鑒定大果沙棘品種的來源。分析采用混合模型(admixture model),考慮各亞種的等位基因頻率相關(allele frequencies correlated),分組數K值設置為1~10,每個K值重復計算10次,每次的運算進行1 000 000次迭代和100 000次BURNIN。利用Structure Harvester[16]計算STRUCTURE運算結果中各K值所對應的似然值lnP(D)與模型值ΔK,根據lnP(D)和ΔK最大的原則[17]選擇合適的分組數。
2 結果與分析
2.1 沙棘亞種與品種的14個SSR位點及種間關系分析
沙棘7個亞種的14個微衛星位點平均NA為2.643~4.643,NE為1.869~3.065,I為0.559~1.053,HO為2.460~0.477,HE為0.293~0.523,F為0.002~0.300。其中:中國沙棘的遺傳多樣性為所有沙棘亞種中最高,有著最高的NE,I,HO和HE(表3),其次為云南沙棘,但云南沙棘的F遠高于中國沙棘,顯示云南沙棘個體間差異較大。高加索沙棘的NE,I,HE和F在所有沙棘亞種中最低,表明其遺傳多樣性水平最低,次低為海濱沙棘。總體來看,亞洲分布的中國沙棘、云南沙棘、蒙古沙棘和中亞沙棘遺傳多樣性普遍高于歐洲分布的溪生沙棘、海濱沙棘和高加索沙棘。26個大果沙棘品種的等位基因個數最多,可能與其個體數最多有關,但是其NE,I,HO,HE與各沙棘亞種的值相比都屬于中等。
沙棘亞種兩兩之間的FST都顯著大于0(表4),表明各亞種間分化明顯。其中:蒙古沙棘和中亞沙棘之間分化最低(FST=0.274),次低為中國沙棘和云南沙棘之間(FST=0.283);中國沙棘和高加索沙棘之間遺傳分化最高(FST=0.535)。亞洲分布的沙棘亞種與歐洲分布的沙棘亞種之間分化普遍高于這2個地區內部亞種之間的分化。
2.2 沙棘亞種與品種的系統發育分析
系統發育樹結果顯示:除大果沙棘品種外,所有沙棘個體分成7個分支,與亞種分類基本一致。僅1個中亞沙棘個體列到亞種分支以外,但各分支的支持率并不高。這7個分支可以分成2個大支:歐洲分布的高加索沙棘、溪生沙棘和海濱沙棘以77.6%的支持率聚為一大支;亞洲分布的中國沙棘、云南沙棘、中亞沙棘和蒙古沙棘聚為另一大支(圖1)。大果沙棘品種主要分成2個大支:大部分的品種個體聚成一支,然后與蒙古沙棘亞種野生個體分支聚在一起;另有5個品種(‘新俄2’ ‘Xine No.2’, ‘芬蘭’‘Fenlan’, ‘首都’ ‘Shoudu’, ‘小辣椒’ ‘Xiaolajiao’, ‘深秋紅’ ‘Shenqiuhong’) 以超過 50%的支持率聚為一支,位于歐洲分布的沙棘亞種分支和亞洲分布的沙棘亞種分支之間。大果沙棘品種‘中俄雜4’ ‘Zhongeza No.4’位于中國沙棘和云南沙棘分支的基部。
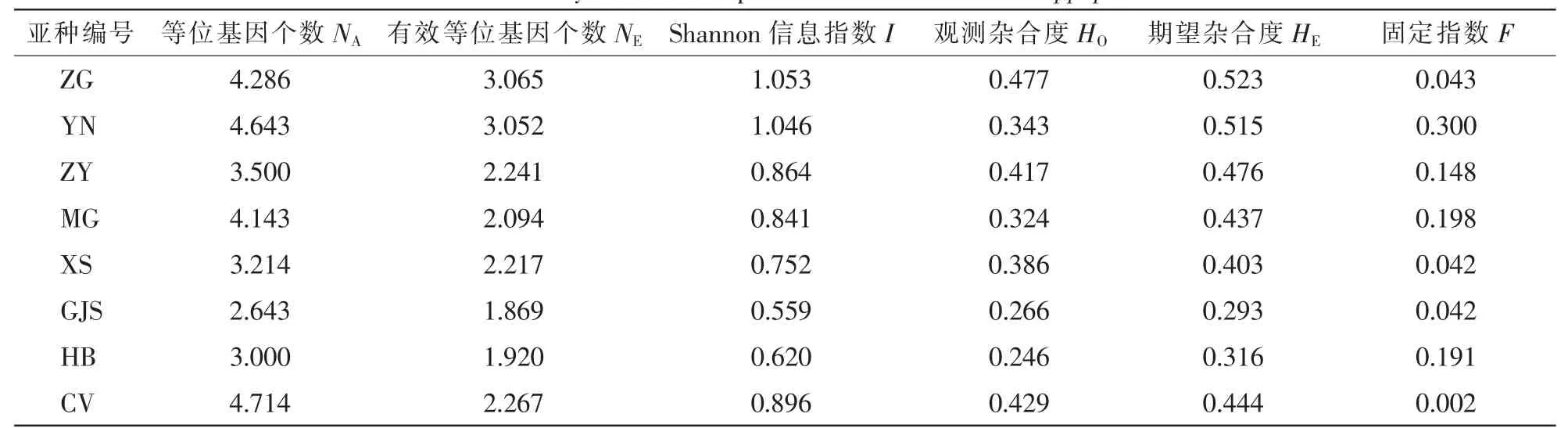
表3 7個沙棘亞種與26個沙棘品種的遺傳多樣性Table 3 Genetic diversity of the 7 subspecies and 26 cultivars of Hippophae rhamnoides
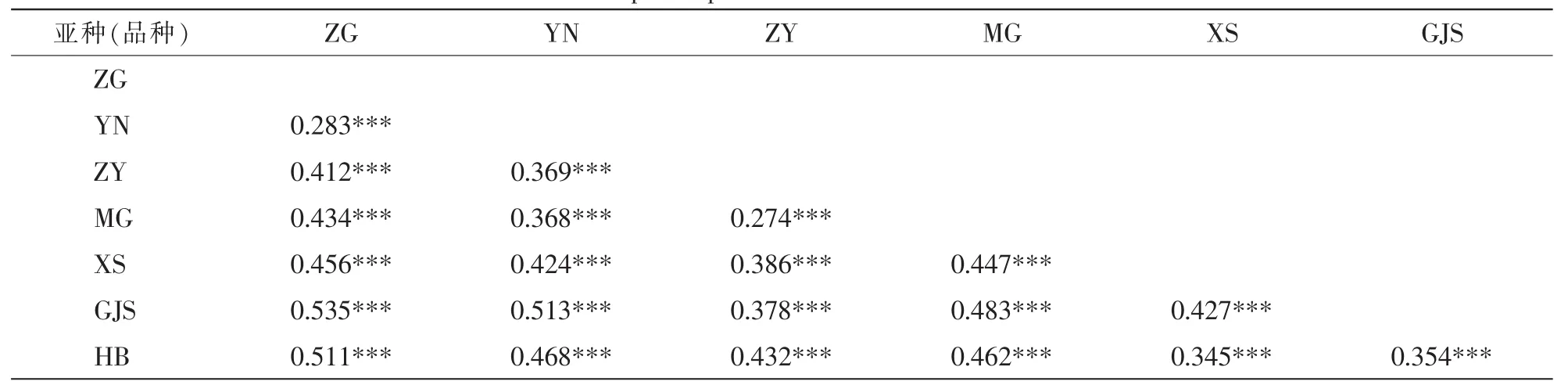
表4 基于14個SSR位點的沙棘亞種間遺傳分化系數Table 4 Subspecies pairwise FST based on 14 SSR loci

圖1 7個沙棘亞種和26個沙棘品種系統發育關系樹Figure 1 Phylogenetic tree of the 7 subspecies and 26 cultivars of Hippophae rhamnoides
2.3 沙棘亞種與品種的貝葉斯聚類分析
貝葉斯聚類分析發現:當K=7時,lnP(D)迅速增加;當K>7時lnP(D)的增加趨于緩慢。可見,K=7是一個明顯的拐點(圖2A)。因此,lnP(K)支持所有的沙棘亞種和品種分為7組。而從ΔK與K的關系可以看出:在K=3處,ΔK有1個顯著的峰值(圖2B),因而支持所有沙棘個體劃分為3組。第1組由中國沙棘和云南沙棘組成,第2組主要由蒙古沙棘和大部分大果沙棘品種組成,第3組由溪生沙棘、高加索沙棘和海濱沙棘組成。中亞沙棘的遺傳成分主要來源于第2組,并包含少量的第3組和更少的第1組遺傳成分。大果沙棘品種 ‘新俄2’ ‘芬蘭’ ‘首都’ ‘小辣椒’和 ‘深秋紅’的遺傳組成中第2組和第3組各約占50%;大果沙棘品種 ‘巨人’ ‘Juren’的遺傳組成主要來自第2組,第3組約占20%;大果沙棘 ‘中俄雜4’的遺傳組成中第1組和第2組各約占50%。當所有個體劃分為7組時,中國沙棘、云南沙棘、中亞沙棘、蒙古沙棘、溪生沙棘、高加索沙棘和大部分大果沙棘品種各自組成一組,但云南沙棘包含少量其他各組的成分,中亞沙棘和蒙古沙棘包含大果沙棘組的成分,溪生沙棘包含高加索沙棘和云南沙棘的遺傳成分;海濱沙棘由溪生沙棘和高加索沙棘的遺傳成分共同構成,大果沙棘品種 ‘新俄2’ ‘芬蘭’ ‘首都’ ‘小辣椒’和 ‘深秋紅’的遺傳成分一半來自大果沙棘組,另一半由溪生沙棘和高加索沙棘組共同構成,大果沙棘 ‘中俄雜4’則中國沙棘和大果沙棘組各約占50%。
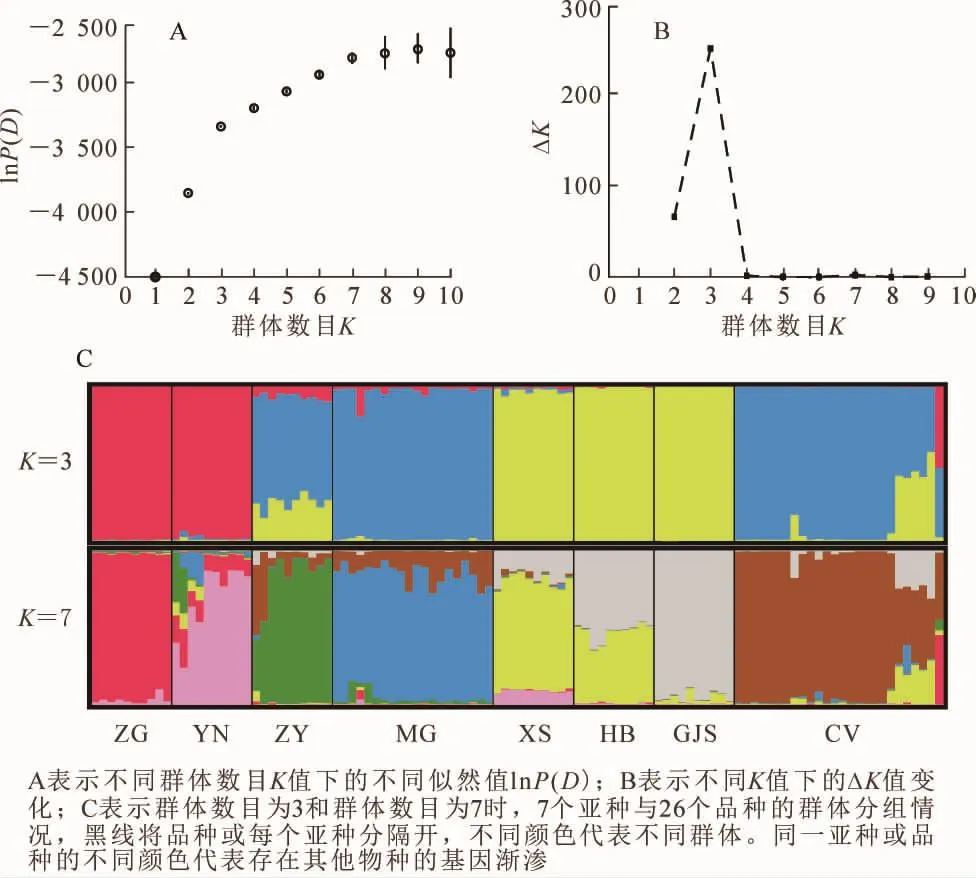
圖2 7個沙棘亞種和26個沙棘品種Structure聚類分析Figure 2 Structure cluster analysis of the 7 subspecies and 26 cultivars of Hippophae rhamnoides
3 討論
3.1 沙棘亞種的分化及親緣關系
廉永善等[18]根據形態特征分析認為:沙棘種下的9個亞種可歸為3組:中國沙棘、云南沙棘和臥龍沙棘H.rhamnoidessubsp.wolongensis分為第1組;蒙古沙棘、高加索沙棘和中亞沙棘為第2組;喀爾巴千山沙棘H.rhamnoidessubsp.carpatica,海濱沙棘和溪生沙棘為第3組。馬玉花等[8]基于ITS序列建立的系統樹認為沙棘9個亞種分為2個組:高加索沙棘、海濱沙棘、喀爾巴千山沙棘、中亞沙棘、云南沙棘和臥龍沙棘為一組;蒙古沙棘、溪生沙棘與中國沙棘為另一組。本研究基于14個微衛星位點的基因型構建的系統發育樹可以看出:7個沙棘亞種主要分成2個組:歐洲分布的海濱沙棘、溪生沙棘和高加索沙棘為一組;亞洲分布的中國沙棘、云南沙棘、蒙古沙棘和中亞沙棘為另一組。組間不同亞種之間的遺傳分化系數也明顯高于組內不同亞種間。這說明跨歐亞大陸的地理隔離已造成歐洲和亞洲分布的沙棘亞種之間更為明顯的分化。貝葉斯分析顯示:這7個沙棘亞種可分為3個組:中國沙棘和云南沙棘分為第1組;蒙古沙棘和中亞沙棘為第2組;海濱沙棘、溪生沙棘和高加索沙棘為第3組。該分組結果與廉永善等[18]根據形態特征的分組更為相近,只是在高加索沙棘的歸屬上稍有差別。高加索沙棘可能在歐洲分布的時間較短,導致高加索沙棘與溪生沙棘、海濱沙棘的基因交流更廣泛,但在形態學上,高加索沙棘還未能從蒙古沙棘和中亞沙棘區分開。
本研究還發現:在系統發育樹中,沙棘個體大都按亞種聚類,即來自同一亞種的個體聚為一支;在貝葉斯聚類分析中,當分7組時,除海濱沙棘由溪生沙棘組和高加索沙棘組共同構成外,其他亞種都各自分成一組。這說明沙棘各亞種間存在明顯的遺傳分化,與兩兩沙棘亞種間較高的遺傳分化系數相符。基因交流的限制[19]和對各自生境的適應性進化都可能造成沙棘亞種間明顯的遺傳分化。然而,貝葉斯聚類分析顯示:各沙棘亞種間可能存在少量的雜交和基因漸滲;系統發育樹中每個亞種的分支支持率并不高,也說明種間雜交可能存在。這與JIA等[5]利用cpDNA片段以及ITS序列對沙棘種下9個亞種的分析結果一致。然而,要進一步確認各亞種間雜交和基因漸滲的方向和程度,還需要更多樣品和基因組范圍內更多位點的詳細分析。
3.2 沙棘亞種的遺傳多樣性比較
JIA等[5]推測沙棘主要起源于中國青藏高原地區,中國東部和歐洲分布的沙棘亞種都是從該地區擴散而來。本研究發現:中國分布的中國沙棘和云南沙棘的遺傳多樣性水平遠高于歐洲分布的溪生沙棘、高加索沙棘和海濱沙棘,這與沙棘起源于中國青藏高原地區的假說相符。相比分布廣泛的中國沙棘,云南沙棘僅分布在西藏、四川、青海和云南交界的狹長地帶,分布范圍較小,較高的遺傳多樣性可能與其分布鄰近起源中心有關。云南沙棘分布區地形復雜,環境變化多端,因此,該亞種各個體也可能因適應不同的環境而積累了較高的遺傳變異[19]。云南沙棘有著最高的固定指數F,也說明其個體間遺傳變異較大。
分析樣品的數量和采集方式也可能會影響沙棘亞種遺傳多樣性的評估。在本研究中,除蒙古沙棘為20個個體外,其他沙棘均為10個個體。因此當亞種間進行比較時,不存在樣品數量的偏差。另外,除高加索沙棘取自種子的播種苗外,其他沙棘亞種的個體都采自2~4個野外種群,這也將減小地理種群的偏差。當然,要準確評估沙棘各亞種內的遺傳多樣性水平,還需整個分布區更全面的采樣。
3.3 大果沙棘品種的來源和多樣性比較
基于14個位點的微衛星基因型構建的系統發育樹中,大部分大果沙棘品種都和采自中國新疆阿勒泰地區的蒙古沙棘亞種聚在一起;貝葉斯聚類分析將所有沙棘個體劃分為3組時(K=3),大果沙棘品種也和蒙古沙棘亞種為一組,這與大果沙棘人工選育自蒙古沙棘亞種的記錄相符。然而,系統發育樹和貝葉斯聚類分析都能將大果沙棘品種和中國新疆阿勒泰地區的蒙古沙棘個體分開,說明用來品種選育的俄羅斯蒙古沙棘種群與中國新疆阿勒泰地區的蒙古沙棘種群存在明顯的遺傳分化。系統發育樹和貝葉斯聚類分析表明:本研究所分析的大果沙棘品種并非都選育自蒙古沙棘亞種,品種 ‘新俄2’ ‘芬蘭’ ‘首都’ ‘小辣椒’和 ‘深秋紅’可能是蒙古沙棘和歐洲分布的沙棘亞種雜交的結果。
本研究還發現:所收集的26個大果沙棘品種多樣性稍高于中國新疆阿勒泰地區的蒙古沙棘亞種野生種群,與本研究的7個沙棘亞種相比屬于中等偏上水平,表明用于栽培的大果沙棘品種具有豐富的遺傳變異。劉雨娜等[20]曾利用RAPD標記分析了24個大果沙棘品種得出同樣的結論。大果沙棘品種來源于蒙古沙棘的遠緣雜交后代,這自然會提高大果沙棘的多樣性水平;其次,本研究表明:少量大果沙棘品種具有歐洲分布的其他沙棘亞種的遺傳成分,說明這些沙棘可能是蒙古沙棘和歐洲其他沙棘種雜交的結果,這也提高了大果沙棘的多樣性水平。雖然大果沙棘品種有著豐富的遺傳變異,但其多樣性仍低于從野外采集的少量中國沙棘和云南沙棘個體。這說明,中國豐富的沙棘自然資源可為沙棘新品種的選育提供重要的遺傳基礎。
