短枝木麻黃幼苗對低溫脅迫的生理響應
2019-07-24 08:19:18李賀鵬葉碧歡陳友吾李海波
浙江農林大學學報 2019年4期
關鍵詞:質量
李 楠,李賀鵬,江 波,葉碧歡,陳友吾,李海波
(浙江省林業科學研究院,浙江 杭州310023)
短枝木麻黃Casuarina equisetifolia作為木麻黃科Casuarinaceae木麻黃屬Casuarina的代表樹種,是世界各國引種最早、人工栽培面積最大的木麻黃植物[1],也是中國東南沿海地區重要的防護林樹種[2],發揮著巨大的生態效益和經濟價值。木麻黃是典型的熱帶植物,生長適溫為22.1~26.9℃,耐寒性較差[3]。引種的木麻黃常遭受較嚴重凍害,給沿海防護林建設造成較大損失。因此,研究木麻黃對低溫的響應機制,開展耐寒品種的選育,對于擴大木麻黃種植范圍和推進沿海防護林建設尤為重要。在低溫誘導下,植物形成了其各自特有的生理生化特征,研究植物的耐寒生理機制,對于解析植物的耐寒性具有重要意義。目前,國內外對木麻黃耐寒生理研究大多集中在抗性生理指標的測定分析上[2,4-6],但從全面或更深入揭示木麻黃的低溫適應的機制還遠遠不夠。本研究針對木麻黃生產實踐中存在的低溫凍害問題,在前期對短枝木麻黃耐寒性評價的基礎上,以短枝木麻黃耐寒無性系 ZS7和不耐寒無性系 HN1幼苗為材料,通過測定這2種無性系幼苗在低溫脅迫下的葉綠素、滲透調節物質、酶類和非酶類抗氧化劑等耐寒相關生理指標,比較分析耐寒性不同無性系生理指標變化的趨勢與差異,為深入闡明短枝木麻黃耐寒機理,促進木麻黃耐寒品種選育奠定基礎。
1 材料與方法
1.1 試驗材料與方法
根據前期試驗結果,選定2個耐寒性差異較大的短枝木麻黃無性系:耐寒無性系ZS7和不耐寒無性系HN1,采用盆栽方式,于2016年10月在浙江省林業科學研究院溫室進行扦插繁殖。塑料盆尺寸為 13 cm(高)×20 cm(直徑)。盆栽基質為泥炭和蛭石混合物(4∶1),裝盆前進行滅菌處理。培養條件為光照16 h/黑暗8 h,溫度 25℃,光合光子照度180 mol·m-2·s-1,相對濕度 60%~70%。在溫室中培養 2個月后,扦插苗高度達到15~20 cm時,選取生長良好、長勢較一致的植株,開始低溫處理。低溫梯度處理設計:將ZS7和HN1無性系置于低溫人工氣候箱(PRXD-300,上海喬躍)中,從常溫預冷至10℃左右,之后逐步降溫至-2,-5,-8,-11℃,在降至每個低溫節點持續2 h后,各取出5株幼嫩枝條用錫箔紙包裹后迅速放入液氮中,置-80℃超低溫冰箱中保存,用于生理指標測定。以在常溫25℃下同步培養的ZS7和 HN1無性系作為2種低溫處理的對照(ck),取樣方法同上。每處理均設3次重復。連續低溫處理設計:將ZS7和HN1無性系各40株移入低溫人工氣候箱內,在-5℃下連續處理 1,2,5,8,16,24,48,72 h,并于每個時間點分別取ZS7和HN1的5株幼嫩枝條,用錫箔紙包裹后迅速放入液氮中,置于-80℃超低溫冰箱中保存備用。
1.2 生理指標測定
葉綠素質量分數的測定采用分光光度計法[7],過氧化氫(H2O2)質量摩爾濃度測定按照高洪波等[8]的方法,丙二醛(MDA)質量摩爾濃度測定采用硫代巴比妥酸(TBA)法[9],可溶性蛋白質測定采用考馬斯亮藍法[10],脯氨酸(Pro)質量分數測定采用酸性茚三酮比色法[11],超氧化物歧化酶(SOD)活性測定采用氮藍四唑(NBT)光化還原法測定[12], 過氧化氫酶(CAT)活性測定采用紫外吸收法[13], 過氧化物酶(POD)活性測定采用愈創木酚顯色法[14],抗壞血酸過氧化物酶(APX)活性測定參照程玉靜等[15]的方法,谷胱甘肽還原酶(GR)活性測定參照PINHEIRO等[16]方法。還原型谷胱甘肽(GSH)和氧化型谷胱甘肽(GSSG)質量摩爾濃度參照JIANG等[17]的方法。
1.3 數據處理
采用Excel 2010和SPSS 18.0對數據進行統計分析,采用鄧肯(Duncan)新復極差法進行5%水平的差異顯著性分析。
2 結果與分析
2.1 低溫脅迫對不同耐寒性短枝木麻黃幼苗葉綠素質量分數的影響
耐寒無性系ZS7幼苗在低溫梯度脅迫下葉綠素質量分數隨著脅迫的加劇,先升高后下降并趨于平緩。在降至-2℃時,ZS7的葉綠素質量分數迅速升高并達到峰值,升幅達63.2%;降至-11℃時,雖有所下降,但和對照相比差異不顯著。不耐寒無性系HN1幼苗的葉綠素質量分數則呈顯著下降趨勢(圖1A),至低溫脅迫結束,葉綠素質量分數降幅達45.7%,受到低溫脅迫的傷害相對較重。在各脅迫溫度下,ZS7的葉綠素質量分數相對穩定,始終顯著高于HN1。
在-5℃持續低溫脅迫下,ZS7和HN1的葉綠素質量分數變化趨勢如圖1B所示。ZS7葉綠素質量分數在1~5 h內與常溫對照沒有明顯差異,8 h后葉綠素質量分數開始顯著升高。HN1在5~72 h內,葉綠素質量分數呈逐漸下降趨勢,且始終顯著低于ZS7,說明-5℃的持續低溫脅迫對HN1的葉片產生了破壞作用,而在短期內對耐寒無性系ZS7葉片的葉綠體結構破壞不明顯。
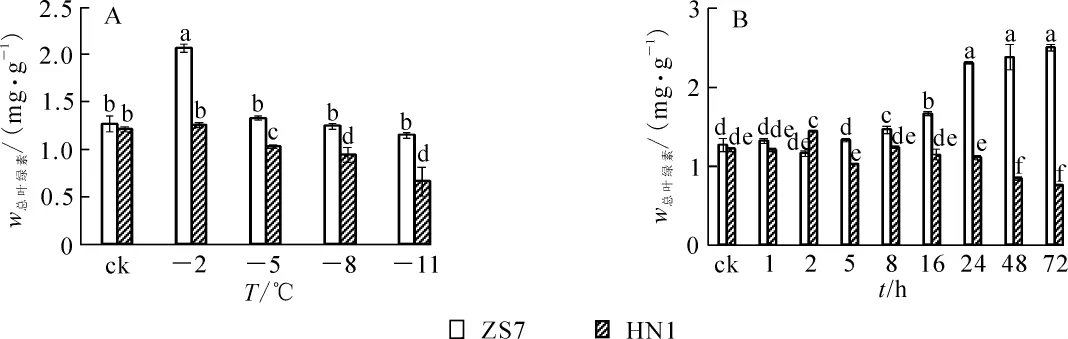
圖1 低溫梯度脅迫和連續低溫脅迫對短枝木麻黃幼苗葉綠素質量分數的影響Figure 1 Effect of successive low temperatures and low temperature (-5℃)conditions for 1-72 h on chlorophyll content in Casuarina equisetifolia
2.2 低溫脅迫對不同耐寒性短枝木麻黃幼苗活性氧水平及丙二醛質量摩爾濃度的影響
隨著低溫梯度脅迫的增強,ZS7和HN1幼苗葉片的H2O2質量摩爾濃度都呈現先升高后降低的變化趨勢(圖2A)。在降至 -5℃時,2種無性系的H2O2質量摩爾濃度均達到最高值,其中ZS7的H2O2質量摩爾濃度升高80.1%,HN1升高130.7%;在 -5~-11℃,ZS7和HN1的H2O2質量摩爾濃度呈下降趨勢,在低溫梯度脅迫期間HN1的H2O2質量摩爾濃度一直較高。ZS7和HN1的MDA質量摩爾濃度呈升降升的趨勢變化 (圖2B),ZS7的MDA質量摩爾濃度在低溫脅迫過程中始終顯著低于HN1。至脅迫結束,ZS7的MDA質量摩爾濃度增幅為11.2%,HN1的增幅為23%,結果顯示低溫脅迫對耐寒性強的無性系ZS7的質膜損傷程度較低。在-5℃持續低溫脅迫下,ZS7和HN1的H2O2,MDA質量摩爾濃度均隨處理時間的延長呈現波動變化,且在脅迫結束時達到最高值(圖2C和2D)。從低溫脅迫處理開始至結束,ZS7的H2O2和MDA質量摩爾濃度的增幅分別為142.4%和73.2%,HN1增幅分別為176.9%和84.7%。與不耐寒無性系相比,低溫下耐寒無性系葉片中MDA質量摩爾濃度一直較低,且差異顯著。
2.3 低溫脅迫對不同耐寒性短枝木麻黃幼苗可溶性蛋白質和脯氨酸質量分數影響
無論在常溫條件下還是在低溫梯度脅迫下,ZS7中可溶性蛋白質質量分數一直維持較高水平,并始終顯著高于HN1(圖3A和3B)。至低溫脅迫結束,ZS7和HN1的可溶性蛋白質質量分數的漲幅和降幅分別為13.9%和12.8%,和耐寒性強弱表現一致。在-5℃持續低溫脅迫下,ZS7和HN1的可溶性蛋白質質量分數總體先升后降。-5℃處理24 h后,ZS7和HN1的可溶性蛋白質質量分數均達到最高,增幅分別為80.8%和43.2%;72 h后,都達到最低值,降幅分別為5.7%和57.5%。
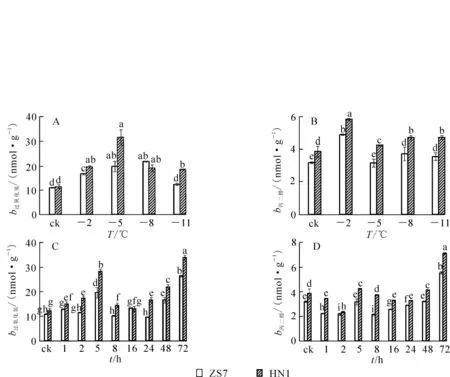
圖2 低溫梯度脅迫和連續低溫脅迫對短枝木麻黃幼苗H2O2和MDA質量摩爾濃度的影響Figure 2 Effect of successive low temperatures and low temperature (-5℃)conditions for 1-72 h on H2O2and MDA contents in Casuarina equisetifolia
在低溫梯度脅迫下,ZS7和HN1中脯氨酸質量分數先上升后下降(圖3C)。處理結束后,2種無性系的脯氨酸質量分數均有所降低,降幅分別為36.3%和16.5%。在-5℃持續低溫脅迫下,ZS7和HN1的脯氨酸質量分數呈波動變化(圖3D)。低溫處理72 h后,2種無性系中脯氨酸質量分數的漲幅分別為37.1%和37.3%。ZS7的脯氨酸質量分數在低溫梯度脅迫和連續低溫脅迫下始終顯著高于HN1。
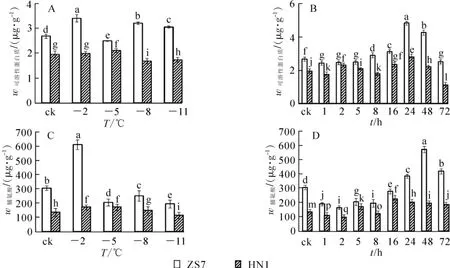
圖3 低溫梯度脅迫和連續低溫脅迫對短枝木麻黃幼苗可溶性蛋白質和脯氨酸質量分數的影響Figure 3 Effect of successive low temperatures and low temperature(-5℃)conditions for 1-72 h on protein and proline content of in C.equisetifolia
2.4 低溫脅迫對不同耐寒性短枝木麻黃幼苗無性系抗氧化酶活性的影響
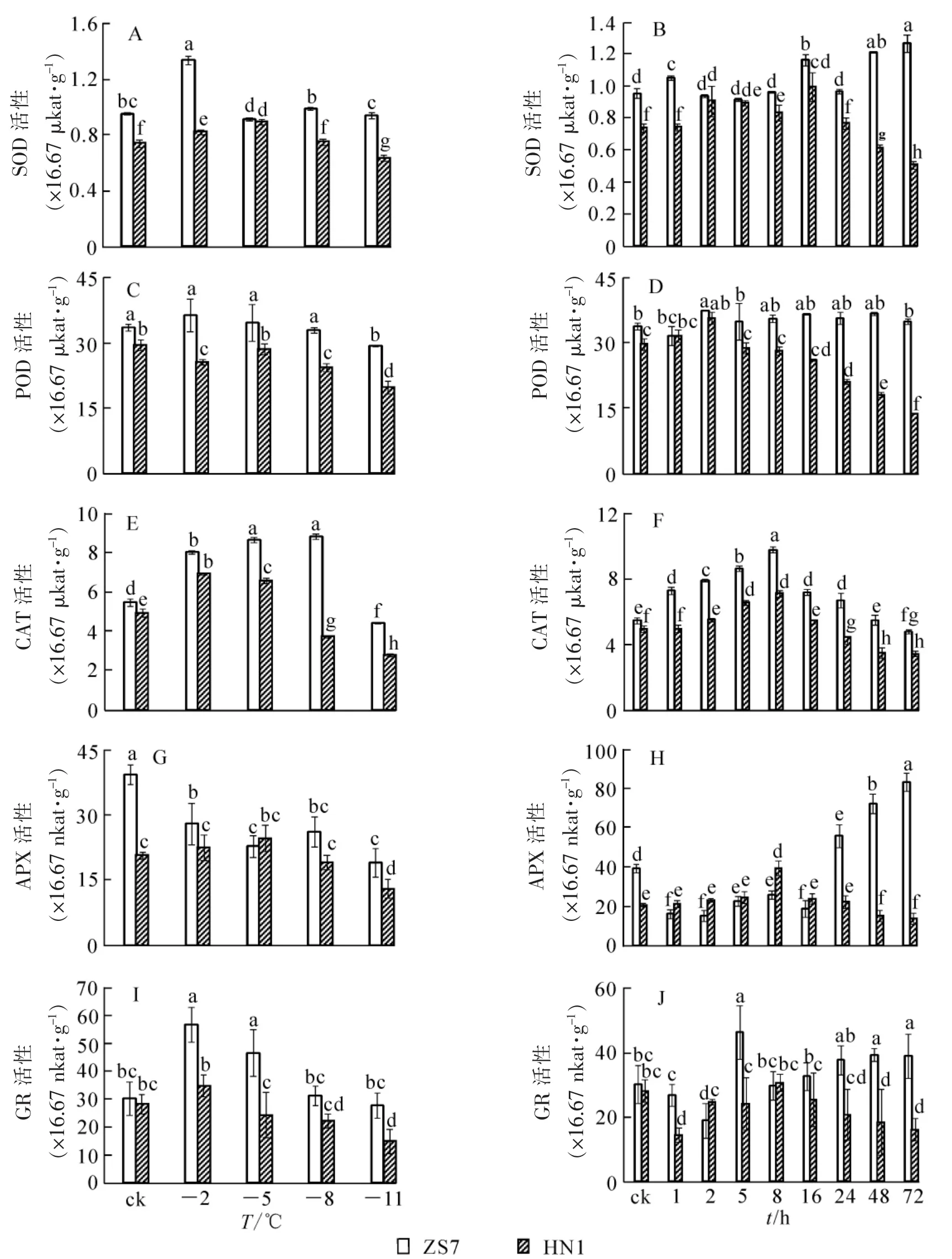
圖4 低溫梯度脅迫和連續低溫脅迫對短枝木麻黃幼苗抗氧化酶活性的影響Figure 4 Effect of successive low temperatures and low temperature (-5℃)conditions for 1-72 h on antioxidant enzyme activity in Casuarina equisetifolia
由圖4可以看出:耐寒無性系ZS7和不耐寒無性系HN1幼苗在低溫梯度處理下SOD,POD,CAT,GR活性變化規律基本一致,呈先上升后下降的變化趨勢;ZS7的APX活性呈下降趨勢,HN1的APX活性則先上升后下降。在常溫條件下,除ZS7和HN1的GR活性沒有顯著差異外,ZS7的SOD,POD,CAT,APX活性均顯著高于HN1,且在整個低溫脅迫過程中ZS7的保護酶活性始終高于HN1,說明耐寒無性系ZS7對過氧化傷害有較高忍耐能力。低溫梯度處理結束后,ZS7和HN1的SOD,POD,CAT,APX,GR活性的降幅分別為1.3%,12.6%,18.9%,51.5%,8.6%和14.3%,33.2%,43.5%,36.9%,47.4%。
在-5℃持續低溫脅迫下,與HN1相比,ZS7葉片中的保護酶活性在整個低溫脅迫過程中始終較高,且兩者達到了顯著性差異(圖4)。ZS7中SOD活性的最高值集中在低溫脅迫48~72 h,且達到常溫對照的32.7%;HN1的最高值集中在2~16 h,并在第72小時降至對照的33.9%。ZS7中POD活性在整個脅迫期間始終維持較高水平;HN1的POD活性在第72小時達到最低,并降至對照的53.7%。ZS7和HN1的CAT活性的最高值都出現在第8小時,至脅迫結束,與常溫對照相比,降幅分別為12.1%和30.9%。ZS7的APX活性在低溫脅迫24~72 h時顯著上升,并升至常溫對照的110%;HN1的APX活性一直較低,在脅迫結束時降至對照的32.6%。此時,ZS7的APX活性是HN1的6倍。至脅迫結束,ZS7的GR活性與對照相比漲幅為28.9%,HN1的降幅為42.7%。說明耐寒無性系ZS7從保護酶活性水平增強了其低溫下的適應能力。
2.5 低溫脅迫對不同耐寒性短枝木麻黃幼苗谷胱甘肽循環的影響
低溫梯度脅迫下,2種無性系幼苗的GSH質量摩爾濃度均呈先升后降的趨勢(圖5A),且都在-5℃時達到最高值,漲幅分別為23.6%和12.3%,說明GSH在2種無性系中均大量累積。脅迫結束時,ZS7和HN1的GSH質量摩爾濃度均達到各自最低值,降幅分別為19.8%和20.7%。在-5℃持續低溫脅迫下,2種無性系的GSH質量摩爾濃度也都呈現先升后降的趨勢(圖5B),但達到峰值的時間不同。ZS7的GSH質量摩爾濃度在第24小時達到最大值,HN1的GSH質量摩爾濃度在第8小時達到最高,漲幅分別為52.7%和32.5%。ZS7中的GSH質量摩爾濃度從第8小時開始一直顯著高于對照,而HN1僅在第5和第8小時時顯著高于對照。72 h后,ZS7和HN1的GSH質量摩爾濃度分別上升9.4%和下降4.5%。低溫處理下,耐寒無性系ZS7的GSH質量摩爾濃度始終顯著高于不耐寒無性系HN1。由圖5C可知:ZS7和HN1幼苗中的GSSG質量摩爾濃度都在-2℃時達到最高值,分別為對照水平的1.4倍和1.9倍;至低溫脅迫結束,GSSG質量摩爾濃度比對照分別降低64.2%和57.1%。在-5℃低溫持續脅迫下,2種無性系中的GSSG質量摩爾濃度總體呈現先上升后下降的趨勢,且HN1中的GSSG質量摩爾濃度在整個脅迫過程中始終顯著高于ZS7。ZS7和HN1中的GSSG質量摩爾濃度,分別于第2小時和第16小時達到最高值,漲幅分別為79.0%和139.0%;至脅迫結束時,分別比對照下降20.1%和19.2%(圖5D)。低溫梯度脅迫下,ZS7的GSH/GSSG比值呈上升的趨勢,HN1則先下降后上升,至脅迫結束時,都顯著高于常溫對照(圖5E)。在 -5℃持續低溫脅迫下,ZS7的GSH/GSSG比值總體呈上升趨勢,HN1則呈下降趨勢(圖 5F)。
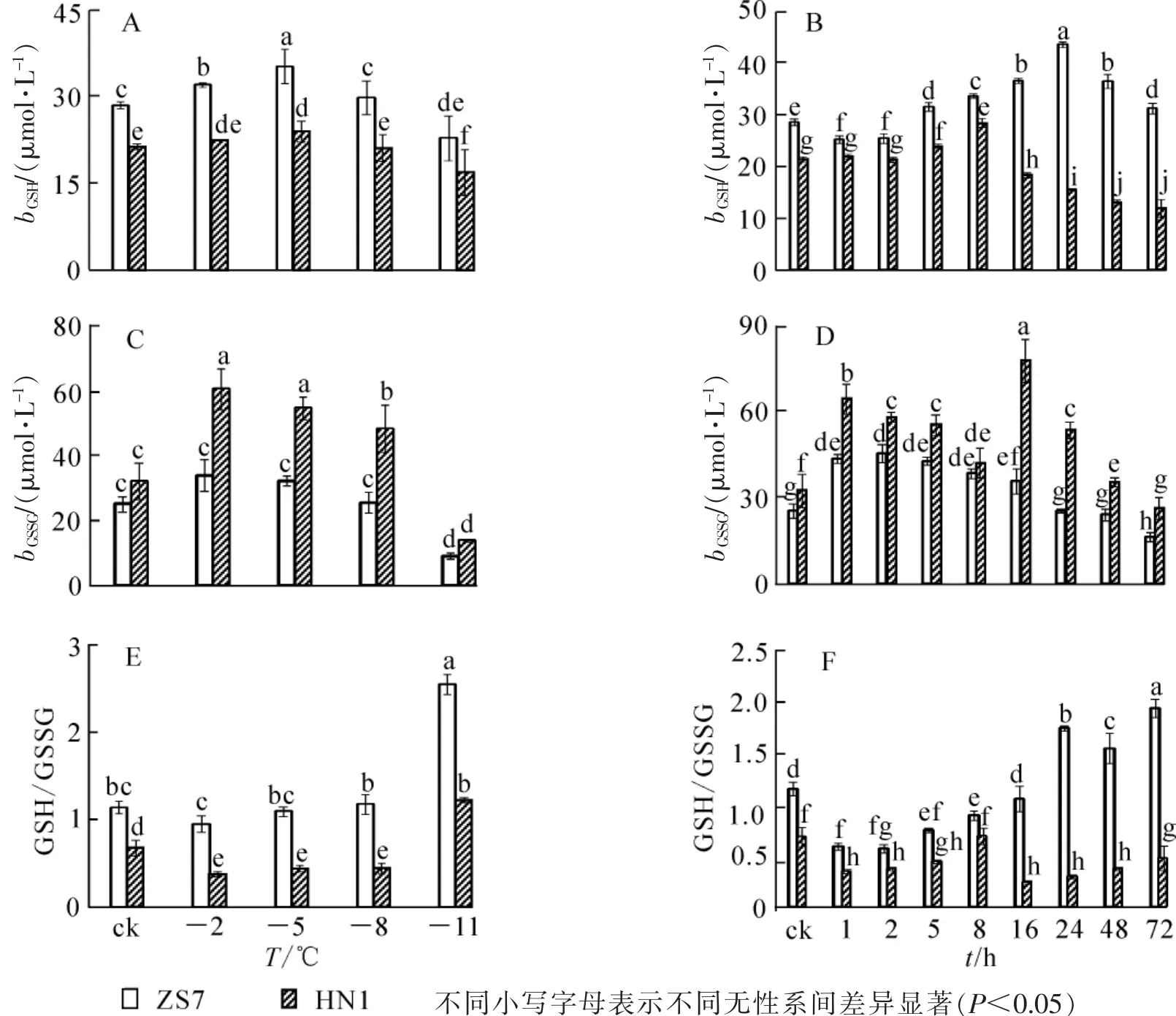
圖5 低溫梯度脅迫和連續低溫脅迫對短枝木麻黃幼苗谷胱甘肽循環的影響Figure 5 Effect of successive low temperatures and low temperature (-5℃)conditions for 1-72 h on glutathione cycle in Casuarina equisetifolia
3 結論與討論
隨著低溫脅迫程度的增強,葉綠素質量分數下降的幅度和速度也加大,可作為植物耐受低溫的重要生理指標[18]。本研究中,低溫梯度脅迫下,不耐寒短枝木麻黃無性系幼苗的葉綠素質量分數呈下降趨勢。在低溫梯度脅迫和-5℃持續低溫脅迫下,耐寒無性系的葉綠素質量分數分別表現出先升高后降低和逐漸上升的趨勢,這說明一定程度的低溫對耐寒植物葉綠素的合成有一定程度的刺激作用,而耐寒植物較強的耐寒性減緩了葉綠素質量分數的下降,增強了適應低溫環境的能力,使葉綠素始終維持較高水平。這在楊樹Populusspp.,金葉女貞Ligustrum vicaryi,紫薇Lagerstroemia indica,福祿考屬Phlox等植物對干旱、低溫等脅迫的生理響應研究中得到相似的結果[19-21]。
H2O2水平升高是低溫下氧化脅迫的標志。它一方面可引起脂質過氧化,對膜造成傷害,另一方面可引起光合系統各種酶的失活[22]。MDA作為膜脂過氧化產物,其質量摩爾濃度的高低與細胞膜受傷害的程度直接相關,與植物的抗寒性呈負相關,是鑒別低溫脅迫對植物膜危害程度的重要指標之一[23]。本研究中,耐寒無性系和不耐寒無性系幼苗的H2O2和MDA質量摩爾濃度都顯著升高,說明低溫脅迫下,植物體內產生大量的活性氧(ROS),細胞膜的結構受到損傷,膜脂過氧化作用加強。但耐寒無性系的MDA和H2O2質量摩爾濃度都顯著低于不耐寒無性系,說明耐寒性較強的無性系能保持細胞內的氧化還原平衡,降低了脂質過氧化程度。
逆境脅迫下,植物細胞能通過主動積累脯氨酸等滲透調節物質來調節滲透勢,阻止細胞脫水,從而維持植物組織內各種酶活性和正常的細胞膜結構[24]。研究發現,脯氨酸在杠柳Peroploca sepium生境適應性上有重要作用,且脯氨酸代謝對植物維持組織細胞內ROS水平密切相關[25]。抗寒性強的油橄欖Olea europaea品種在低溫脅迫下,累積的滲透調節物質含量,如脯氨酸和可溶性蛋白質,逐漸增加且顯著高于其他不耐寒品種[26]。本研究中,耐寒無性系ZS7幼苗在常溫下的脯氨酸和可溶性蛋白質質量分數就顯著高于HN1,且隨溫度的降低或低溫脅迫時間的延長,脯氨酸和可溶性蛋白質質量分數也始終顯著高于HN1。說明在低溫處理下,耐寒無性系通過增強滲透調節能力和降低膜脂過氧化作用,提高對外界環境變化的適應力。
SOD,POD,CAT,APX和GR是植物體內活性氧自由基清除系統的保護酶[27-29],通過協同作用抑制膜脂過氧化,防御活性氧自由基對細胞膜的損傷,緩解低溫脅迫對膜系統的傷害[30]。GR活性和GSH質量分數大小被認為是有機體抗氧化狀態的重要標志。在低溫梯度脅迫下,耐寒無性系和不耐寒無性系的SOD,POD,CAT,GR活性以及GSH質量摩爾濃度均呈先上升后下降的趨勢,表明植物通過調節酶和非酶類活性的高低來適應低溫。GSH/GSSG的比值與蛋白質的生物合成有關,維持較高的比值可以增強植物對低溫的抵抗力。在低溫脅迫下,耐寒無性系中的GSH/GSSG比值呈上升趨勢,不耐寒無性系中則呈下降趨勢,說明GSH和GSSG對于短枝木麻黃耐寒性的提高具有重要作用。耐寒無性系中的APX活性在低溫梯度脅迫下逐漸下降,但在-5℃持續低溫脅迫下,表現出了強烈的誘導作用。這可能與適應低溫的過程中,植物在代謝補償性調節上發揮作用有關,某些代謝過程的增強抑制或降低了另外的代謝途徑,暗示植物在適應低溫環境時發生了積極的代謝調節[31]。低溫脅迫處理下,耐寒無性系的酶類和非酶類抗氧化劑含量高,且始終顯著高于不耐寒無性系,更有利于維持其細胞結構的穩定性。
本研究結果反映了耐寒性強的短枝木麻黃無性系通過在低溫脅迫下保持較高的脯氨酸和可溶性蛋白質質量分數,增強SOD,CAT,POD,APX和GR等抗氧化酶的活性以及提高非酶抗氧劑質量摩爾濃度,抑制植物細胞中葉綠素質量分數的下降及膜脂過氧化程度的加劇,進而適應和抵御低溫。
猜你喜歡
中學生數理化·中考版(2022年10期)2022-11-10 09:37:42
中學生數理化·八年級物理人教版(2022年12期)2022-02-14 07:08:42
中學生數理化·八年級物理人教版(2021年12期)2021-12-31 03:23:08
中學生數理化·中考版(2020年10期)2020-11-27 01:59:48
中國生殖健康(2019年2期)2019-08-23 08:12:08
石油化工建設(2018年6期)2018-04-22 03:16:54
產品可靠性報告(2017年7期)2017-09-05 09:49:12
中學生數理化·八年級物理人教版(2017年12期)2017-04-18 12:59:38
汽車觀察(2016年3期)2016-02-28 13:16:26
民生周刊(2014年7期)2014-03-28 01:30:54
