陸地棉低世代群體纖維品質QTL定位及候選基因功能注釋
2019-07-24 07:16:34喬文青嚴根土石建斌王寧張亞林許慶華周紅黃群
棉花學報 2019年4期
喬文青,嚴根土,石建斌,王寧,張亞林,許慶華,周紅,黃群
(中國農業科學院棉花研究所/棉花生物學國家重點實驗室,河南安陽455000)
全球陸地棉產量占棉花總產的95%以上[1]。陸地棉高產的優勢難以掩蓋其纖維品質較差的缺陷,因此以高產為基礎、提高陸地棉纖維品質的研究是當前重要的研究方向之一。隨著分子標記技術的發展,利用分子標記輔助育種來提高陸地棉纖維品質具有重要意義。
棉花纖維品質性狀屬于數量性狀,具有復雜的遺傳背景且易受環境影響[2-4]。傳統的分子標記如簡單重復序列標記(Simple sequence repeats,SSR)雖然具有成本低、可獲得性及穩定性高等特點而被廣泛應用于數量性狀位點(Quantitative trait loci,QTLs)分析[5-9],但是利用 SSR 篩選獲得的QTLs較少、置信區間較大,難以直接應用于分子輔助育種。單核苷酸多態性(Single nucleotide polymorphism,SNP)作為1種重要的分子標記,能夠將同一位點的不同等位基因之間有差異的核苷酸標記出來[10],具有位點豐富、數量多、密度大、可覆蓋全基因組、代表性高、穩定性高等特點。研究人員利用SNP標記獲得大量高密度遺傳圖譜[11-15],這些遺傳圖譜為目標性狀QTLs定位奠定了基礎,而陸地棉基因組的公布[16-17],則為QTLs基因功能注釋及挖掘提供了條件,為尋找與纖維品質有關的基因提供了可能性。盡管遺傳圖譜構建和QTLs作圖方法等不斷改進和發展,受QTLs的置信區間相對較大、能夠解釋的表型變異相對較低、相關序列信息缺少等[18-19]的限制,定位精度和準確度依然有待提高,QTLs定位仍處于基礎研究階段,與生產實踐結合不緊密。目前尚未見利用分子標記輔助育種方法育成棉花新品系或新品種的報道。因此,構建有指導意義的可重復利用的作圖群體、結合陸地棉基因組信息開展QTLs定位研究、利用高密度遺傳圖譜做精細定位、深度挖掘基因信息等進行標記輔助育種[20]具有重要意義。
本研究主要利用2個陸地棉品種(系)中棉所49和396289構建F2群體,以高密度遺傳圖譜[21]為基礎對纖維品質性狀進行QTLs定位,找出在多環境下的低世代群體中穩定存在的QTLs進行基因功能注釋,挖掘與纖維品質有關的基因,以提高育種進度及效率。該研究同時為分子標記輔助育種提供理論支撐,為QTLs精細定位和有關功能基因的發掘提供依據。
1 材料與方法
1.1 試驗材料與種植方法
本試驗以陸地棉栽培品種(系)中棉所49(簡稱為“中49”)和396289為親本配制雜交組合,于2015年夏天在中國農業科學院棉花研究所東場試驗地(河南安陽)雜交,同年冬天在海南加代獲F2,2016年春天將F2群體種植于河南安陽,隨機篩選160個單株于7月份取親本及單株的幼嫩葉片,于-80℃超低溫冰箱中儲存。對160個單株做自交,構建F2:3家系,將單株自交鈴種子隨機分成3份,于2017年分別種植于河南安陽(17AY)、安徽安慶(17AQ)以及新疆阿拉爾(17ALE),均采用單行區種植。其中:17AY行長3 m,行間距0.8 m,株距0.25 m;17AQ行長5 m,行間距1.2 m,株距0.5 m;17ALE行長5 m,行間距0.6 m,株距0.1 m。
1.2 DNA提取及分子檢測
取中49和 396289及其構建的F2群體的160個單株的幼嫩新葉,采用改良的CTAB法[22]提取DNA。利用親本重測序和160個子代SLAF簡化基因組測序技術(Specific-locus amplified fragment sequencing,SLAF-seq[23])、HighMap[24]軟件,對F2遺傳分離群體開發高密度分子標記并構建遺傳圖譜[21]。
1.3 表型數據的獲得與分析
隨機摘取F2:3家系株行材料的25個正常吐絮棉鈴,按照農業農村部棉花纖維品質監督檢驗測試中心(河南省安陽市)的要求提供材料并委托其檢測纖維品質,檢測儀器為HVI1000和AFIS pro2。其中:HVI1000檢測的指標包括5個,分別為纖維上半部平均長度(纖維長度,FL)、斷裂比強度(比強度,FS)、長度整齊度指數(整齊度,FU)、馬克隆值(馬克隆值,Mic)和斷裂伸長率(伸長率,FE);AFIS pro2檢測的指標包括纖維細度(細度,Fin)和纖維成熟度(成熟度,Mat)。 品質指標后括號中依次為文中簡稱和QTLs命名中性狀對應的字母標記。
表型數據利用MS Excel 2007統計最大值、最小值、極差、平均值、偏度、峰度等,使用SAS9.2進行方差分析。
1.4 QTLs定位方法
QTLs定位使用軟件MapQTL6,利用復合區間作圖法,優勢對數(Logarithm of odds,LOD)值設為2.5。QTLs的命名規則:(q+性狀)+染色體/連鎖群+QTL數目[5]。QTLs作用方式用顯性度(|D/A|,即顯性效應與加性效應的比值的絕對值)表示,|D/A|小于0.20時為加性作用,在0.21~0.80為部分顯性作用,在0.81~1.20為顯性作用,大于1.20為超顯性作用[25]。
1.5 基因功能注釋
確定一致性QTLs后,提取親本QTLs區域的序列信息,進行開放閱讀框(Open reading frame,ORF)預測,獲得基因信息,同時獲取雙親間的非同義突變基因,作為最終的候選基因。應用BLAST軟件對置信區間的編碼基因通過直系同 源 基 因 簇(Clusters of orthologous groups,COG)、基因本體(Gene ontology,GO)和京都基因與基因組百科全書(Kyoto Encyclopedia of Genes and Genomes,KEGG)等數據庫做深度注釋。其中:COG數據庫可對候選基因產物做直系同源分類;GO數據庫從生物過程、分子功能和細胞組分3個方面對候選基因做富集分析;KEGG數據庫用于基因的代謝通路分析。
2 結果與分析
2.1 纖維品質表型數據基本統計
在3個F2:3家系中,纖維品質相關性狀偏度絕對值均小于1,表型值近似符合正態分布(表 1)。
纖維長度:同一環境下中49長于396289,且二者在不同的環境中均具有極顯著差異;整齊度:同一環境下中49高于396289,但不同環境中兩者的顯著性差異程度不同;比強度:17AY和17AQ的中49極顯著高于396289,而17ALE中二者則無顯著差異;馬克隆值、伸長率、細度:17AY和17AQ的中49和396289有極顯著差異,17ALE中則無顯著差異;成熟度:中49與396289在17AQ中有顯著差異,在17AY和17ALE中無顯著差異。
在F2:3家系中,除比強度在17AY和17AQ中為單向超親分離(F2:3家系中比強度的最大值大于/小于親本的最大值,家系的最小值大于/小于親本的最小值)、在17ALE中為雙向超親分離(F2:3家系中的最大值大于親本的最大值,最小值小于親本的最小值)之外,其余性狀在不同環境均為不同程度雙向超親分布。
2.2 纖維品質性狀的相關分析
通過纖維品質相關分析發現,不同環境中纖維品質間的相關性可能不同,但總的來說:纖維長度與比強度、伸長率呈極顯著正相關,與細度呈極顯著負相關;整齊度與伸長率呈顯著正相關;比強度與伸長率呈極顯著正相關,與馬克隆值、細度呈極顯著負相關;馬克隆值與細度、成熟度呈極顯著正相關;伸長率與細度呈顯著負相關;細度與成熟度呈極顯著正相關(表2)。
2.3 纖維品質QTLs定位
不同性狀的QTLs的統計列于表3。
纖維長度:共獲得25個與纖維長度有關的QTLs, 分布于 A03、A04、A07等 8條染色體上,能夠解釋7%~13.6%的表型變異。17AY、17ALE和17AQ檢測到的QTLs分別為2、11和14個,其中qFL-A03-1在3個環境中均能檢測到,屬于穩定QTL,其加性效應為正,有利等位基因來自父本中49,具有正向部分顯性作用。
整齊度:共獲得19個與整齊度有關的QTLs,分布于 A02、A03、A04 等 11 條染色體上,能夠解釋6.9%~16.2%的表型變異。17AY、17ALE和17AQ檢測到的QTLs分別為7、8和5個,其中qFU-A04-1在2個環境(17ALE和17AQ)可檢測到,屬于穩定QTL,其加性效應為正,有利等位基因來自父本中49,具有正向超顯性作用。
比強度:共獲得25個與比強度有關的QTLs, 分布于 A03、A04、A07等 7條染色體上,能夠解釋7%~11.5%的表型變異。17AY、17ALE和17AQ檢測到的QTLs分別為9、1和15個。
馬克隆值:共獲得26個與馬克隆值有關的QTLs, 分布于 A03、A04、A11等 8條染色體上,能夠解釋7.1%~16.4%的表型變異。17AY、17ALE和17AQ檢測到的QTLs分別為11、7和11 個 ,qMic-A11-1、qMic-D03-1 和 qMic-D12-2均能在17AY和17ALE環境中檢測到,屬于穩定QTLs,其中qMic-A11-1的加性效應為正,有利等位基因來自父本中49,具有正向超顯性作用;qMic-D03-1的加性效應為負,有利等位基因來自母本396289,在17ALE和17AY中分別具有負向顯性作用和負向部分顯性作用;qMic-D12-2的加性效應在不同環境中有差異,其有利等位基因來自不同的親本,而QTLs的作用同樣表現為不同的方向,但都具有超顯性作用。

表1 不同環境中纖維品質相關性狀的基本統計Table 1 Basic statistics of traits related to fiber quality traits in different environments
伸長率:共獲得19個與伸長率有關的QTLs, 分布于 A03、A04、A08等 6條染色體上,能夠解釋7.3%~13.3%的表型變異。17AY、17ALE和17AQ檢測到的 QTLs分別為 6、1和14個,其中qFE-A04-1和qFE-A04-2在17AY和17AQ環境中均能檢測到。qFE-A04-1的加性效應為正,有利等位基因來自父本中49,在17AY和17AQ中分別具有正向超顯性和正向顯性作用;qFE-A04-2的加性效應同樣為正,具有正向顯性作用。
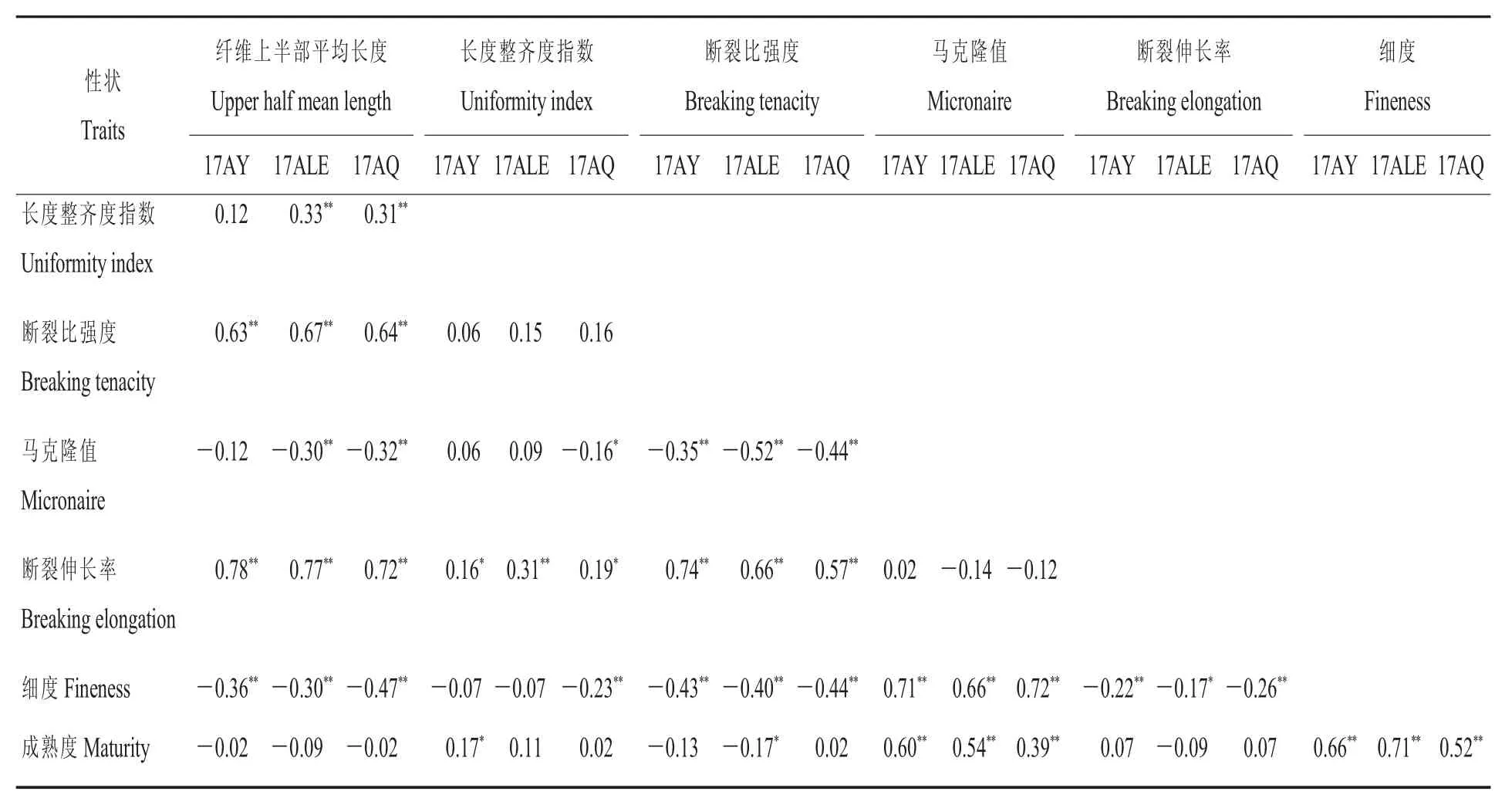
表2 不同環境下纖維品質性狀的相關分析Table 2 Correlation analysis of fiber quality traits in different environments
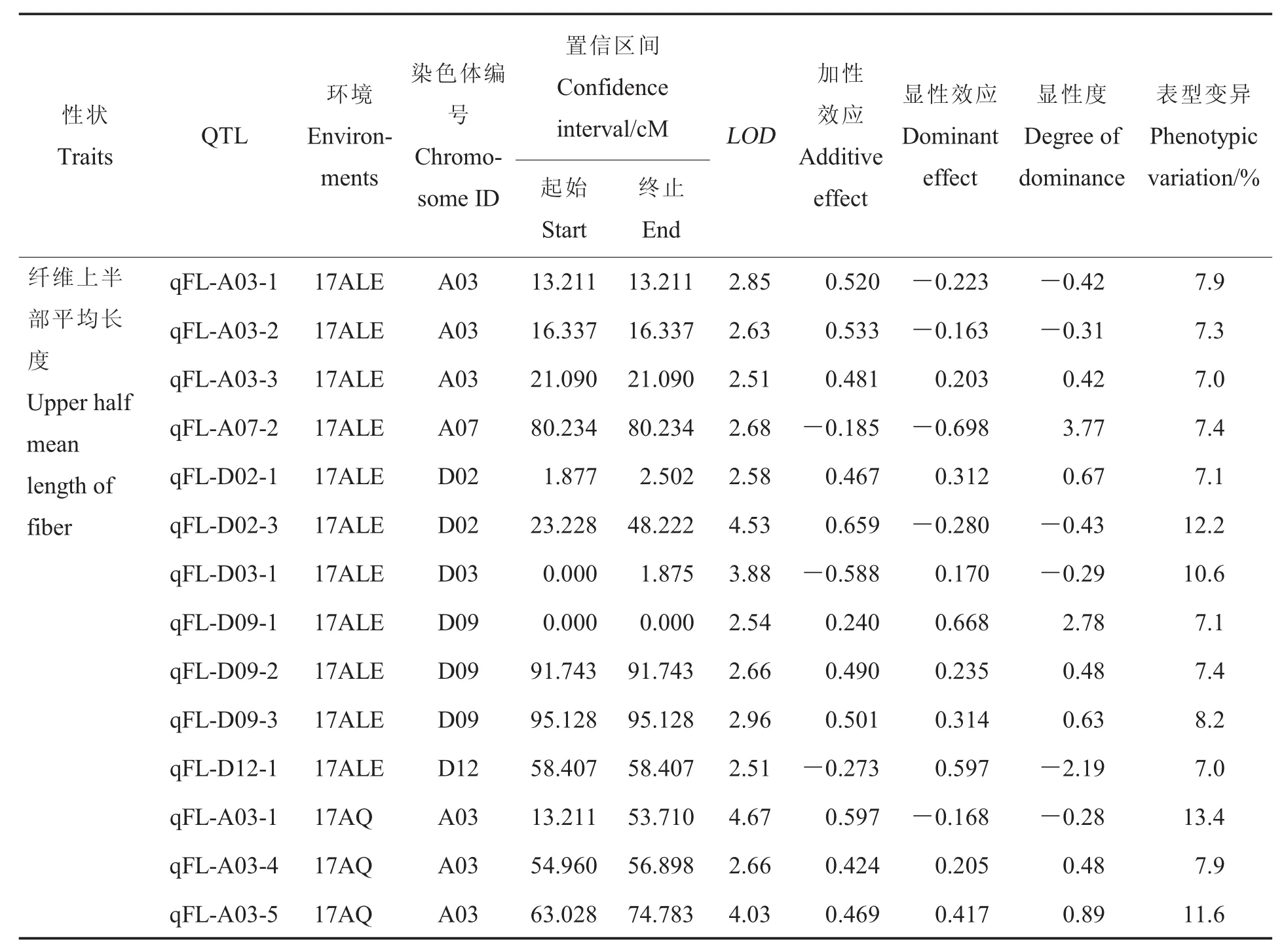
表3 纖維品質相關性狀的QTLTable 3 QTLs for fiber quality traits
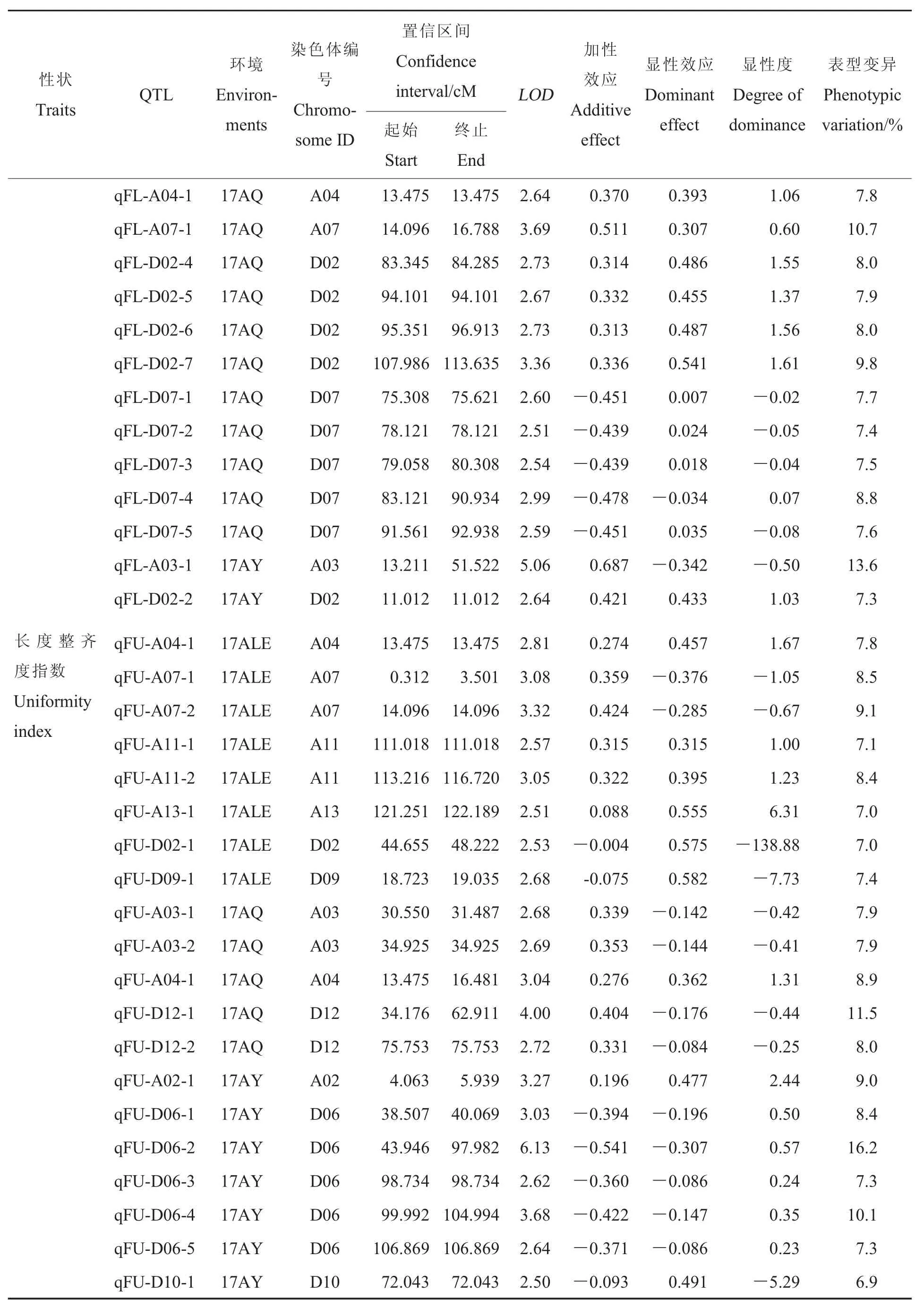
表3(續)Table 3 (Continued)
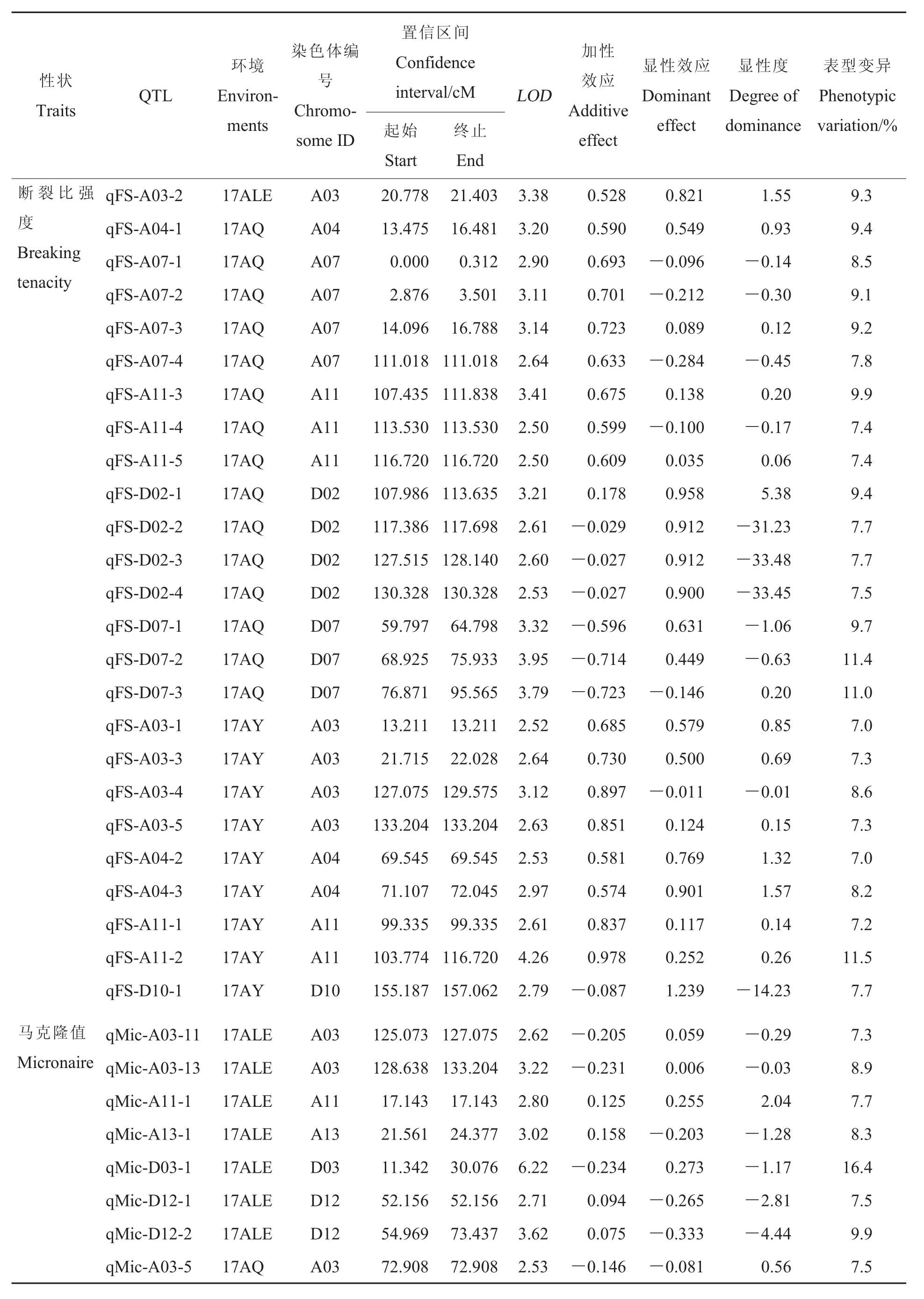
表3(續)Table 3 (Continued)
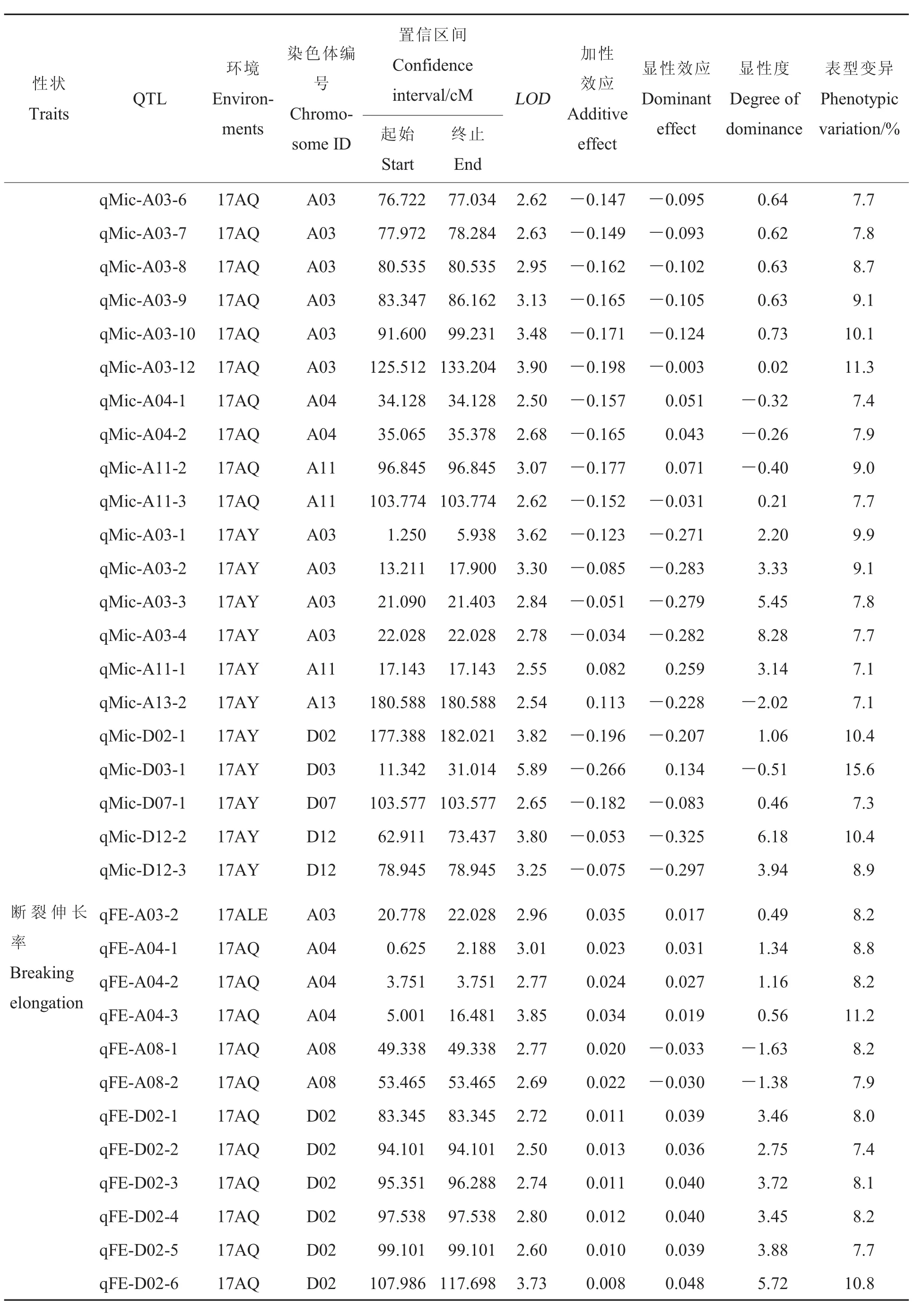
表3(續)Table 3 (Continued)
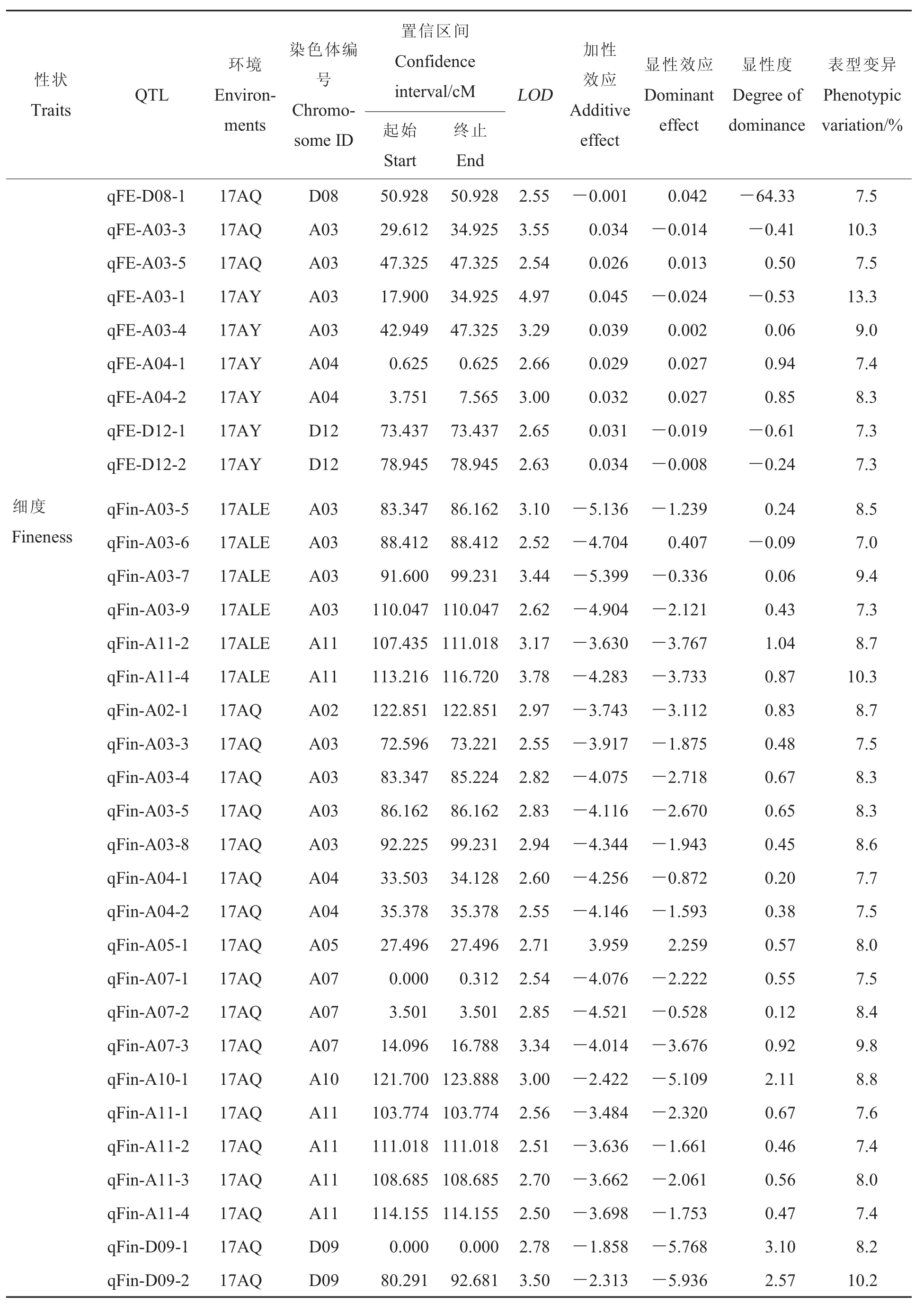
表3(續)Table 3 (Continued)
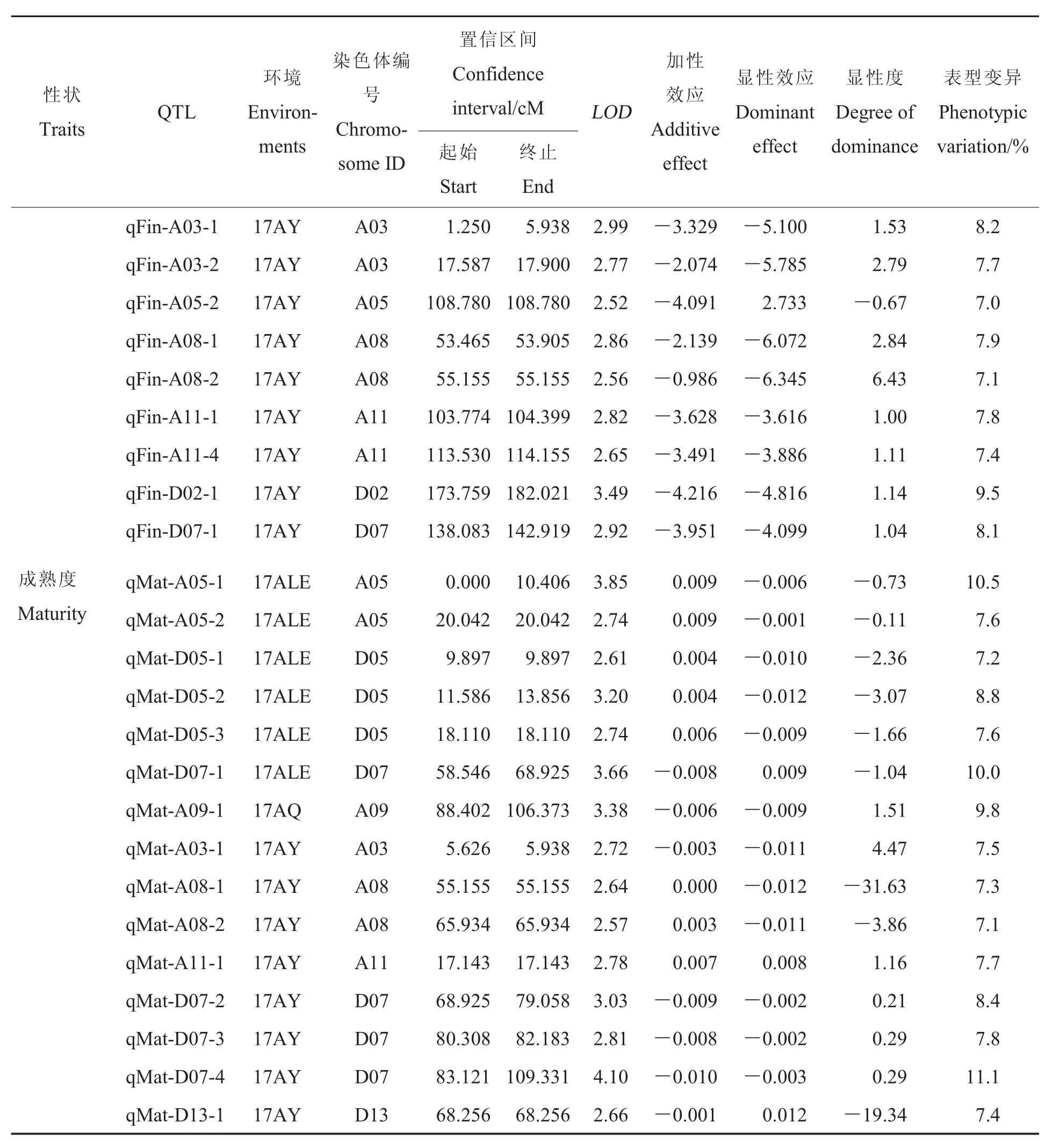
表3(續)Table 3(Continued)
細度:共獲得28個與細度有關的QTLs,分布于A02、A03、A04等11條染色體上,能夠解釋7%~10.3%的表型變異。17AY、17ALE和17AQ檢測到的 QTLs分別為 9、6和 18個,其中qFin-A11-4在3個環境中均能檢測到,qFin-A03-5和qFin-A11-2在17ALE和17AQ環境中均能檢測到,而qFin-A11-1在17AY和17AQ環境中能檢測到。
成熟度:共獲得15個與成熟度有關的QTLs, 分布于 A03、A05、A08等 8條染色體上,能夠解釋7.1%~11.1%的表型變異。17AY、17ALE和17AQ檢測到的QTLs分別為8、6和1個。
2.4 纖維品質QTLs候選區域內的基因功能注釋
纖維長度:與纖維長度有關的QTLs置信區間中共有1 434個候選基因,其中1 002個被注釋,各QTL中候選基因數從0到274個不等(附表1,印刷版略,電子版參見本刊網站)。在COG分類有513個注釋基因,其中有151、75、70、69、62個基因分別與一般功能預測、信號轉導機制、轉錄、翻譯后修飾/蛋白質周轉/伴侶蛋白、復制/重組/修復等有關。在GO富集中有901個注釋基因,其中在細胞組成中,有較多的基因與細胞核、膜的組成部分、質膜等有關;在分子功能中,有較多的基因與ATP結合、金屬離子結合、DNA結合有關;在生物過程中,有較多的基因與氧化還原過程、轉錄調控、DNA模板化、蛋白質磷酸化等有關。穩定QTL qFL-A03-1置信區間中只有1個基因Gh_A03G0097,該基因是膜的組成部分,參與硫代葡萄糖苷生物合成過程、植物細胞壁纖維素代謝過程以及細胞壁果膠代謝過程。在KEGG分析中,524個基因參與96條通路,其中,有 31、16、16、16、16 個基因分別與核糖體、 碳代謝、谷胱甘肽代謝、植物-病原體相互作用、蛋白質在內質網中的加工通路有關等。
整齊度:與整齊度有關的QTLs置信區間中有2 024個候選基因,其中1 330個被注釋,各QTL中候選基因數從0到954個不等。在COG分類中有636個注釋基因,其中有203、121、115、99、56個基因分別與一般功能預測、轉錄、復制/重組/修復、信號轉導機制、翻譯后修飾/蛋白質周轉/伴侶蛋白等有關。在GO富集中有1 171個注釋基因,在細胞組成中,有較多的基因與細胞核、膜的組成部分、質膜等有關;在分子功能中,有較多的基因與ATP結合、金屬離子結合、DNA結合等有關;在生物過程中,有較多的基因與氧化還原過程、蛋白質磷酸化、轉錄調控、DNA模板化有關。在KEGG分析中,693個基因參與107 條通路。 其中,有 28、27、24、23、21 個基因分別與核糖體、碳代謝、植物激素信號轉導、RNA運輸、氧化磷酸化通路有關等。在穩定QTL qFU-A04-1置信區間中未發現候選基因。
比強度:與比強度有關的QTLs置信區間中有957個候選基因,其中618個被注釋,各QTL中候選基因數目從0到316個不等。在COG分類中有 290 個注釋基因,其中有 76、47、39、35、33個基因分別與一般功能預測、轉錄、信號轉導機制、復制/重組/修復、翻譯/核糖體結構/生物發生等有關。在GO富集中有562個注釋基因,在細胞組成中,有較多的基因與細胞核、膜的組成部分、質膜有關;在分子功能中,有較多的基因與ATP結合、DNA結合、金屬離子結合有關;在生物過程中,有較多的基因與氧化還原過程、轉錄調控、DNA模板化、蛋白質磷酸化有關。在KEGG分析中,321個基因參與94條通路。其中,有 22、14、11、9、8 個基因分別與核糖體、 氨基酸的生物合成、碳代謝、植物激素信號轉導、谷胱甘肽代謝通路有關。
馬克隆值:與馬克隆值有關的QTLs置信區間中有1 454個候選基因,其中990個被注釋,各QTL中候選基因數目從0到630個不等。在COG分類中有506個注釋基因,其中有143、62、59、58、47個基因分別與一般功能預測、轉錄、信號轉導機制、復制/重組/修復、翻譯后修飾/蛋白質周轉/伴侶蛋白等有關。在GO富集中有872個注釋基因,在細胞組成中,有較多的基因與膜的組成部分、質膜、細胞核有關;在分子功能中,有較多的基因與ATP結合、鋅離子結合、金屬離子結合有關;在生物過程中,有較多的基因與氧化還原過程、轉錄調控、DNA模板化、代謝過程有關。在KEGG分析中,518個基因參與96條通路。 其中,37、25、17、16、15 個基因分別與植物激素信號轉導、核糖體、碳代謝、內質網中的蛋白質加工、淀粉和蔗糖代謝通路有關等。穩定QTL的qMic-A11-1的置信區間中無候選基因;qMic-D03-1共 有 424個 候 選 基 因(Gh_D03G1014—Gh_D03G1437),其中有注釋信息的基因有309個,在COG、GO和KEGG中分別有163、276和173個;qMic-D12-2的置信區間中共有122個候選基因(Gh_D12G1203—Gh_D12G1324),其中有注釋信息的基因有87個,在COG、GO和KEGG中分別有49、82和46個。
伸長率:與伸長率有關的QTLs置信區間中有535個候選基因,其中364個被注釋,各QTL中候選基因數目從0到215個不等。在COG分類中有 177 個注釋基因,其中有 51、28、24、23、17個基因分別與一般功能預測、轉錄、復制/重組/修復、信號轉導機制)、翻譯后修飾/蛋白質周轉/伴侶蛋白等有關。在GO富集中有335個注釋基因,在細胞組成中,有較多的基因與細胞核、膜的組成部分、質膜有關;在分子功能中,有較多的基因與ATP結合、DNA結合、金屬離子結合有關;在生物過程中,有較多的基因與氧化還原過程、轉錄調控,DNA模板化、蛋白質磷酸化有關。在KEGG分析中,208個基因參與72條通路。其中,10、9、9、7、5 個基因分別與碳代謝、植物 -病原體相互作用、核糖體、氨基酸的生物合、戊糖和葡萄糖醛酸相互轉化通路有關等。在穩定QTL qFE-A04-1和qFE-A04-1置信區間中未發現候選基因。
細度:與細度有關的QTLs置信區間中有946個候選基因,其中613個被注釋,各QTLs中候選基因數目從0到225個不等。在COG分類中有 305 個注釋基因, 其中有 86、43、38、29、29個基因分別與一般功能預測、轉錄、信號轉導機制、復制/重組/修復、翻譯后修飾/蛋白質周轉/伴侶蛋白等有關。在GO富集中有512個注釋基因,在細胞組成中,有較多的基因與膜的組成部分、質膜、細胞核有關;在分子功能中,有較多的基因與ATP結合、金屬離子結合、核苷酸結合有關;在生物過程中,有較多的基因與氧化還原過程、轉錄調控,DNA模板化、跨膜運輸有關。在KEGG分析中,333個基因參與92條通路。其中,33、11、10、9、8 個基因分別與植物信號轉導、氨基酸的生物合成、核糖體、吞噬、氧化磷酸化通路等有關。穩定QTLs qFin-A11-1和qFin-A11-2置信區間中無候選基因,qFin-A11-4的置信區間中14個候選基因(Gh_A11G2846—Gh_A11G2859),其中有注釋信息的基因有10個,在COG、GO和KEGG中分別有3、8和6個;qFin-A03-5共有32個候選基因(Gh_A03G0852—Gh_A03G0883),其中有注釋信息的基因有22個,在COG、GO和KEGG中分別有11、20和10個。
成熟度:與成熟度有關的QTLs置信區間中有1 728個候選基因,其中1 240個被注釋,各QTL中候選基因數目從0到703個不等。在COG分類中有643個注釋基因,其中有200、126、112、107、69 個基因分別與一般功能預測、轉錄、信號轉導機制、復制/重組/修復、翻譯/核糖體結構/生物發生等有關。在GO富集中有1 092個注釋基因,在細胞組成中,有較多的基因與膜的組成部分、細胞核、質膜有關;在分子功能中,有較多的基因與ATP結合、金屬離子結合、核糖體的結構成分有關;在生物過程中,有較多的基因與氧化還原過程、轉錄調控,DNA模板化、蛋白質磷酸化有關。在KEGG分析中,667個基因參與 100 條通路。 其中,53、27、24、22、19 個基因分別與核糖體、植物激素信號轉導、碳代謝、氨基酸的生物合成、內質網中的蛋白質加工通路等有關。
3 討論
3.1 纖維品質各指標間的表型分析
通過對親本和F2:3家系在多環境下表現研究發現,同一環境中,親本的成熟度差異不明顯,但其他性狀有顯著差異,這可能說明環境對成熟度影響不大。纖維品質相關性分析表明,性狀間的相關性因環境而異,但不同環境中,馬克隆值與細度和成熟度間的相關性一致,這可能與馬克隆值是細度和成熟度的綜合指標有關。基于紡紗工業對纖維品質的要求,本研究結果可為育成紡織工業要求的棉花新材料提供理論依據,同時為同步改良部分纖維品質性狀提供理論依據。
3.2 纖維品質相關QTL定位
親本材料、群體類型和分子標記類型等方面的不同為發現大量QTLs提供可能,同時也有助于發現控制同一性狀的穩定QTLs[5-9]。隨著高通量測序技術的發展,利用高密度遺傳圖譜[19,26]對纖維品質進行定位越來越普遍,而前人的研究大多集中在纖維長度、整齊度、比強度、馬克隆值和伸長率等5個指標,很少研究細度和成熟度等指標。本研究通過對包括細度和成熟度在內的7個纖維品質性狀定位,共獲得157個與纖維品質有關的QTLs,其中有99個QTLs的遺傳距離在1 cM以下,與前人[3,9,19]的研究相比,QTLs的置信區間明顯縮小。qFL-A03-1和qFin-A11-4在3環境中均能檢測到,有9個QTLs在其中2個環境中能檢測到,這說明了低世代群體中也可能存在穩定QTLs,這些穩定的QTLs對分子標記輔助育種具有重要參考價值。除此之外,有7個性狀的QTLs分布在A03上,6個性狀的QTLs分布于A04和D02上,5個性狀的QTLs分布于A11和D07上,這種多個性狀的QTLs集中在同一條染色體呈簇分布的現象,與He等[27]和秦永生等[28]的研究結果相似,這些QTLs簇可能與纖維品質各性狀有關, 同時也說明 A03、A04、D02、A11 和D07等染色體極有可能是控制纖維品質性狀的關鍵染色體,可作為纖維品質改良的重點研究內容,在分子標記輔助育種中具有重要參考價值。
3.3 纖維品質QTLs區域內候選基因的功能注釋
在7個性狀中,共有4 763個注釋基因,其中在COG中有2 416個注釋基因,分為24類,其中含有基因數較多的類有一般功能預測、轉錄、信號轉導機制、復制/重組/修復、翻譯后修飾/蛋白質周轉/伴侶蛋白、翻譯/核糖體結構/生物發生等,這些基因可能與纖維發育有關,影響纖維品質。GO富集中有4 188個注釋基因,其中,在生物過程中含有基因最多的3個亞類包括氧化還原過程、轉錄調控、DNA模板化;在細胞組成中含有最多基因的3個亞類是細胞核、膜的組成部分、質膜;在分子功能分類中還有基因最多的3個亞類是ATP結合、金屬離子結合、核糖體的結構成分;該結果與各性狀的GO分類結果類似。Qin等[29]和Tang等[30]研究表明,活性氧在纖維發育中起關鍵作用,在本研究中與纖維長度有關的基因Gh_A03G0483參與對活性氧的反應,與整齊度有關的基因Gh_A07G0233參與對活性氧的反應,Gh_A11G2849參與活性氧代謝過程的調控;與比強度有關的基因Gh_A11G2849參與活性氧代謝過程的調控;與馬克隆值有關的基因Gh_D03G1055參與對活性氧的反應;與細度有關的基因Gh_A11G2849參與活性氧代謝過程的調控。上述基因可能是調控纖維品質的重要基因,其作用機理有待進一步研究。在KEGG通路中有2 521個注釋基因參與121個通路,其中核糖體、植物激素信號轉導、碳代謝通路是參與基因數最多的通路。一些研究表明類黃酮途徑可能參與纖維的發育過程[31-33]。同樣有研究表明植物激素信號轉導途徑對纖維發育有影響[34]。上述與纖維發育有關的通路,在本研究中均有注釋到,因此與這些通路相關的基因可能參與纖維發育的過程,有可能影響纖維品質,但其作用機理有待進一步研究。
4 結論
本研究以高密度遺傳圖譜為基礎,對纖維品質性狀QTLs進行定位及候選基因功能注釋,主要結論如下:(1)纖維品質各性狀的表型分析發現,纖維長度與比強度和伸長率呈極顯著正相關、與細度呈極顯著負相關,纖維強度與伸長率呈極顯著正相關、與馬克隆值和細度呈極顯著負相關,馬克隆值與細度和成熟度呈極顯著正相關,該結果不僅為篩選適宜紡織工業的新材料提供理論依據,而且為同步改良部分纖維品質性狀提供理論依據。(2)本研究通過對包括細度和成熟度在內的7個纖維品質性狀進行QTLs定位,獲得157個QTLs,分布于20條染色體上,有的染色體上分布有控制多個性狀的QTLs,如:A03、A04和D02等,這些染色體極有可能是控制纖維品質性狀的關鍵染色體。除此之外,獲得了11個穩定的QTLs,其中qFL-A03-1和qFin-A11-4在3個環境中重復出現,另有9個QTLs在2個環境中重復出現。(3)在 11個穩定 QTLs中,qFL-A03-1、qMic-D03-1、qMic-D12-2、qFin-A11-4和qFin-A03-5上共有593個候選基因,其中有429個有注釋信息,可通過進一步的篩選來驗證是否是控制纖維品質的關鍵基因。
附表1:QTL候選基因在COG、GO和KEGG數據庫的基因功能注釋信息(見期刊網站)
