低溫對枇杷苗葉片與根尖的生理狀況及超微結構的影響
2019-08-08 06:58:30杜麗君牛先前林曉紅張玉容鄭國華
熱帶亞熱帶植物學報 2019年4期
杜麗君,牛先前,林曉紅,張玉容,鄭國華*
(1.漳州城市職業學院,福建 漳州 363000;2.福建農林大學園藝學院,福州 350002;3.福建省熱帶作物科學研究所,福建 漳州 363001)
枇杷(Eriobotrya japonica)是著名的名優特水果,具有較高的經濟價值。然而,枇杷開花座果期一般在11月至次年1月,此時正值一年中氣溫最低的季節,花果極易受凍。凍害是限制枇杷生產的重要因子。有關枇杷凍害發生機制的研究有較多報道,涉及葉片[1-6]、幼果[7-12]和枇杷外生菌(冰核細菌)[8-10,13]、抗寒基因[14-15]、轉錄組[16]等方面。枇杷凍害機制的理論研究為解決枇杷凍害提供了思路,也為緩解枇杷凍害起到了積極作用,但低溫仍然是限制枇杷生產的主要因子。前人的研究表明,枇杷葉片和幼果在耐寒生理方面具有密切相關性[17],作為植物生產力和養分利用率的重要驅動者——地下部,直接參與養分活化、轉化、吸收和運輸等過程,通過緊密而復雜的物質和信息交換,實現各要素間的相互作用[18]。地上部與地下部在資源獲取和利用方面又具有相對獨立性[18]。已有研究表明,地下部的低溫條件會造成地上部的低溫傷害并影響其低溫傷害程度[19-20]。植物地下部能顯著影響地上部的抗逆性,這在黃瓜(Cucumis sativus)[21]、茄子(Solanum melongena)[22]嫁接苗的抗冷性以及葡萄(Vitis vinifera)[23]嫁接苗的抗旱性上均獲得了證實。因此,研究枇杷地下部與地上部低溫脅迫下的生理變化,認識枇杷地上與地下在耐寒生理方面的相關性,對解決枇杷凍害問題具有重要意義。本文以較不耐寒品種‘解放鐘’枇杷苗為材料,人工模擬自然低溫條件,在透射電鏡下觀察低溫下根尖和葉片細胞超微結構的變化,并結合低溫脅迫下的相關生理變化,探討枇杷地上部與地下部的抗寒生理相關性,以期為枇杷凍害問題提供解決思路。
1 材料和方法
1.1 材料
試驗于2011年10月至2014年1月在福建農林大學園藝場進行。供試枇杷(Eriobotrya japonica)品種為 ‘解放鐘’實生苗,選擇長勢健壯、無病蟲害且整齊度高的1年齡枇杷苗,移入培養缽(直徑20 cm)進行單株培養,培養基質為果園土∶復合肥=20∶1,統一管理。生長1年半后,選擇株高30~35 cm,生長健壯、無病蟲危害的枇杷苗285株為低溫處理試驗材料。
1.2 低溫處理
參考謝鐘琛等[24]的試驗設計,設置3個處理溫度:0℃、-3℃和-5℃。枇杷苗在10℃低溫鍛煉2 d后,采用梯度降溫法,將植株放入改裝的可控溫冰柜(上海精創 ETC-100通用型溫控儀控溫),以10℃ h-1降溫到4℃維持10 h,再以2℃ h-1降溫到設置溫度。在0℃、-3℃、-5℃分別脅迫0(對照)、12、24、36、48、60和72 h后,將植株在10℃恢復生長2 d后統計枇杷苗受害情況,葉片以褐變面積大于總面積1/2判定為受凍害褐變[13]。所有處理均在室內進行,每處理5株,重復3次。葉片褐變率=植株褐變葉片總數/植株葉片總數×100%。
1.3 電鏡觀察
取從頂葉算起第4片功能葉,取完為止,用蒸餾水洗凈,吸干,然后用鋒利刀片沿主脈中部兩側取0.5 mm×1 mm的組織塊,每片葉在相同部位切取4片,重復5次。
割開營養缽后用自來水浸泡,待土松軟后獲得完整根系,用自來水洗凈根系,再用蒸餾水清洗,吸水紙吸干,然后用鋒利刀片從根尖向內1 cm處取0.5 mm×1 mm的組織塊4片,重復5次。
參照鄭國華等[7]的方法制備超薄切片。將切取好的組織塊迅速投入用含2%焦銻酸鉀的磷酸緩沖液(pH 7.6)配制的3%戊二醛固定液中,脫氣后于4℃固定1 d以上;用2%焦銻酸鉀-0.1 mol L-1磷酸氫二鉀(pH 7.6)洗滌4次,每次30 min,4℃過夜;將材料轉移至用含2%焦銻酸鉀的磷酸緩沖液(pH 7.6)配制的1%鋨酸中,4℃冰箱內固定4 h;然后重蒸水洗滌4次,每次30 min,冷乙醇梯度脫水,丙酮過渡,環氧樹脂618包埋,LKB-5型超薄切片機切片(厚度50 nm),經醋酸雙氧鈾染色后,在JEM1010型透射電鏡(日本電子株式會社,日本)下觀察照相。
1.4 生理指標的測定
根系活力測定參考張志良等[25]的方法;細胞質膜相對透性(PMP)的測定參考李合生[26]的方法。
1.5 數據處理
采用DPS軟件的新復極差法對數據進行統計分析,用SigmaPlot作圖。
2 結果和分析
2.1 葉肉細胞超微結構的變化
透射電鏡下,正常枇杷葉肉細胞的細胞壁結構緊密,細胞膜完整,液泡很大占據中央;葉綠體呈橢圓形或近圓形,雙層膜完整,且沿細胞膜排列,切面上可見葉綠體內嵌3~5個明亮的淀粉粒(圖1:A);淀粉粒周圍分布垛疊規則的類囊體片層,并和基質片層連成整體(圖1:B);線粒體分布在葉綠體周圍,呈圓形或橢圓形,雙層膜結構完整,嵴清晰且數量較多(圖 1:C)。
0℃脅迫12 h葉肉細胞的細胞壁未有明顯變化(圖1:D),葉綠體淀粉粒清晰,基粒片層排列有序,但片層之間出現明顯間隙(圖1:D,E);線粒體數量增加2~3倍且嵴更清晰(圖1:E)。脅迫24 h的葉綠體雙層膜結構破壞,淀粉粒清晰,基粒片層垛疊程度降低;線粒體結構保持完整且嵴數量明顯增加(圖1:F)。脅迫36 h的葉綠體雙層膜結構消失,淀粉粒變小,基粒片層結構解體明顯(圖1:G);線粒體膜及嵴模糊不清,細胞內含物明顯增多,細胞壁變化不明顯(圖1:G)。脅迫48 h的葉綠體邊緣解體,淀粉粒開始消失(圖1:H),線粒體變形,雙層膜破裂,嵴消失(圖1:I)。脅迫60 h細胞解體嚴重,細胞內散布小泡、片層、絮狀物,細胞器只有線粒體依稀可辨(圖 1:J)。
-3℃脅迫12 h的葉綠體發生膨脹、扭曲,雙層膜斷裂,基粒片層松散但排列依然整齊(圖1:K);線粒體數量增多但嵴數量減少,并少數出現空胞化(圖1:K)。脅迫24 h細胞壁完好,液泡完全破裂,葉綠體脹裂,雙層膜消失,基粒片層松散并出現解體,淀粉粒清晰(圖1:L);細胞內出現大量絮狀物,線粒體明顯空胞化但尚可辨認(圖1:M)。脅迫36 h后各細胞器解體嚴重,形態紊亂,細胞中零星分布殘存的小泡、顆粒和葉綠體片層(圖1:N)。脅迫60 h細胞器完全解體難以辨別(圖1:O)。
-5℃脅迫12 h的葉綠體膨漲變形,淀粉粒清晰,葉綠體被膜消失(圖1:P);片層結構松散斷裂(圖1:Q),線粒體數量增加2~3倍,嵴消失,內部出現解體和空胞化,細胞壁結構破壞明顯(圖1:Q)。24 h后原生質體縮成一團,細胞器解體嚴重難以辨別(圖 1:R)。
2.2 根細胞超微結構的變化
透射電鏡下觀察,常溫下枇杷根細胞有明顯的質壁分離,中央液泡很大并將其他細胞器擠到邊緣,細胞中無葉綠體(圖2:A),液泡膜結構清晰可見,線粒體呈圓形或橢圓形,雙層膜結構明顯,嵴清晰(圖 2:B)。
0℃脅迫12 h,根細胞液泡膜破裂,泡內基質外流;線粒體數量增多2~3倍(圖2:C),雙層膜結構完整、嵴清晰,細胞壁有破壞現象(圖2:C)。脅迫24 h,細胞壁結構變得松散,內側出現絮狀物(圖2:D);線粒體變形,雙層膜模糊,但嵴依然清晰(圖2:E)。脅迫36 h,細胞壁破壞加劇,線粒體膜破裂,嵴消失,空胞化嚴重(圖2:F)。脅迫48 h時,細胞壁結構出現明顯間隙(圖2:G),線粒體少量依稀可辨,大部分被膜破裂或消失,嵴嚴重解體(圖2:H)。脅迫60 h時,細胞壁破壞嚴重,線粒體解體為絮狀物難以辨別(圖 2:I)。
-3℃脅迫12 h,細胞膜結構破壞明顯,液泡消失,原生質體縮成一團聚在中間(圖2:J),細胞壁邊緣解體出現絮狀物(圖2:K),線粒體數量增多2~3倍,形態完整但嵴的數目明顯減少且部分出現空胞化(圖2:L)。脅迫24 h時細胞壁胞內一側解體加劇(圖2:M),線粒體依稀可辨,嵴嚴重解體或消失(圖2:N)。脅迫36 h后,細胞嚴重破壞,細胞器完全解體,無法辨認(圖 2:O)。
-5℃脅迫12 h,細胞器聚成一團,線粒體外膜破裂、內嵴解體,較難識別(圖2:P)。脅迫24 h時,細胞壁破壞嚴重,細胞器解體,殘體散落在細胞各處(圖2:Q)。脅迫36 h后,細胞器完全解體,細胞內充滿絮狀物(圖 2:R)。
2.3 葉片受害情況
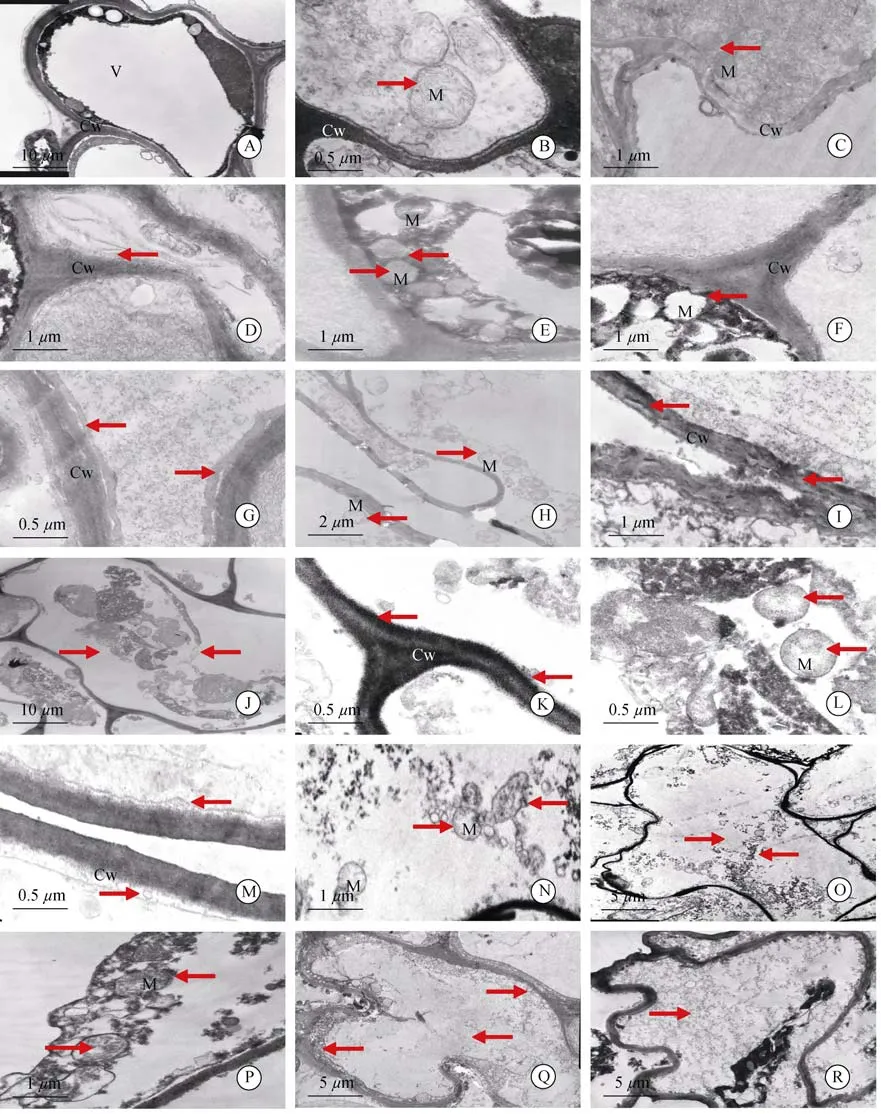
圖 2 低溫脅迫過程中根細胞結構的變化(箭頭)。A,B:正細胞;C:0℃脅迫 12 h;D,E:0℃脅迫 24 h;F:0℃脅迫 36 h;G,H:0℃脅迫 48 h;I:0℃脅迫60 h;J~L:-3℃脅迫 12 h;M,N:-3℃脅迫 24 h;O:-3℃脅迫 36 h;P:-5℃脅迫 12 h;Q:-5℃脅迫 24 h;R:-5℃脅迫 36 h。Fig.2 Changes in structure of root cells under low temperature stress(arrows).A,B:Normal cel;C:Under 0℃ for 12 h;D,E:Under 0℃ for 24 h;F:Under 0℃ for 36 h;G,H:Under 0℃ for 48 h;I:Under 0℃ for 60 h;J-L:Under-3℃ for 12 h;M,N:Under-3℃ for 24 h;O:Under-3℃ for 36 h;P:Under-5℃ for 12 h;Q:Under-5℃ for 24 h;R:Under-5℃ for 36 h.
低溫脅迫下,最先出現低溫傷害癥狀的是幼葉 (約占20%)、其次為成熟功能葉(約占60%)、最不敏感的是老葉(約占20%)。由圖3可見,0℃脅迫12~36 h,葉片褐變率僅增加5.02%,受凍葉片基本為幼葉,而在低溫持續48 h后,成熟葉片出現傷害癥癥狀,脅迫60 h后51.37%的葉片呈現低溫受害癥狀。而-3℃脅迫12 h的傷害癥狀與0℃脅迫60~72 h的相當。枇杷苗在-5℃脅迫12 h后表現出完全凍害癥狀。這說明葉片褐變率與脅迫溫度呈反比,與脅迫時間呈正比,且-3℃脅迫36 h和-5℃脅迫12 h即表現完全凍害癥狀。
2.4 低溫對根系活力的影響
由圖3可見,在0℃、-3℃、-5℃下,枇杷苗的根系活力隨脅迫時間的延長整體呈下降趨勢,且溫度愈低,根系活力下降幅度愈大。0℃脅迫12 h的根系活力顯著下降26.09%(P<0.05),之后根系活力緩慢下降但未達到顯著水平,說明隨0℃脅迫時間的延長,根系受害程度增加。-3℃脅迫0~12、12~24 h的根系活力分別下降32.74%和26.01%,均達極顯著差異,脅迫24 h后變化平緩,說明-3℃脅迫根系即持續受害。-5℃脅迫12 h的根系活力極顯著下降42.13%(P<0.01),之后則下降平緩,說明-5℃脅迫使根系受害嚴重。
2.5 細胞質膜相對透性(PMP)的變化
由圖4:A可見,在0℃、-3℃和-5℃脅迫下,葉片的PMP隨脅迫時間的延長呈持續上升趨勢,且溫度越低上升幅度越高。0℃脅迫12 h,PMP顯著上升37.47%(P<0.05),脅迫48h的PMP僅上升23.6%,與脅迫12h的差異不顯著。-3℃脅迫下,PMP隨脅迫時間的延長持續上升,不同時間間均達到差異顯著水平。-5℃脅迫12 h的PMP極顯著上升51.19%(P<0.01),之后仍緩慢上升,說明低溫脅迫對葉片造成受害。
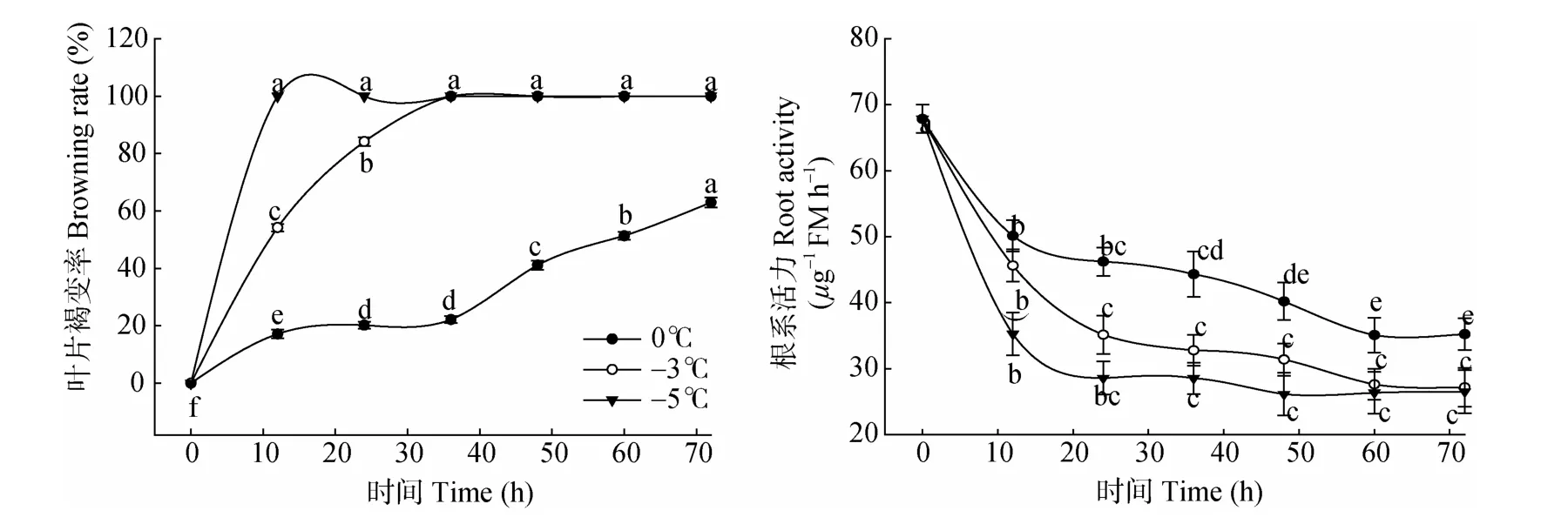
圖3 低溫脅迫下枇杷苗葉片褐變率和根系活力的變化。不同英文字母表示差異顯著(P<0.05)。下同。Fig.3 Changes in leaf browning rate and root vigor of loquat seedlings under low temperature stress.Different letters indicate significant difference at 0.05 level.The same is following figure.
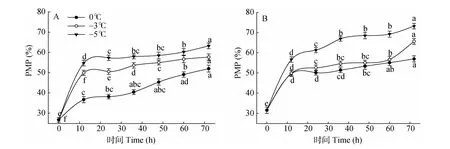
圖4 低溫脅迫對枇杷苗葉(A)、根(B)的PMP變化Fig.4 Changes in leaf(A)and root(B)PMP of Eriobotrya japonica under low temperature stress
由圖4:B可見,在0℃、-3℃、-5℃脅迫下,根系的PMP隨脅迫時間的延長均呈上升趨勢,且脅迫溫度愈低,PMP上升愈高。0℃脅迫12 h的PMP極顯著上升36.49%(P<0.01),之后緩慢上升。-3℃脅迫12、72 h PMP分別比脅迫0、60 h顯著上升了36.74%和13.72%(P<0.05)。-5℃脅迫12、24和36 h的PMP分別比脅迫0、12、24 h顯著上升44.32%、7.53%和8.46%(P<0.05)。
3 討論
許多研究表明,低溫脅迫下植物細胞會出現質壁分離現象[6],因質壁分離可以引起液泡脫水,從而使液泡縮小以增加原生質濃度來減輕低溫傷害[27]。質壁分離程度越高,對細胞結構的破壞程度越大,表明細胞抗低溫能力越弱[28]。本研究中枇杷植株在10℃低溫鍛煉2 d后,根細胞出現質壁分離和液泡縮小現象,而葉肉細胞結構略有變化但未出現質壁分離現象,說明根細胞的抗低溫能力弱于葉肉細胞。
低溫脅迫下細胞可以通過消耗呼吸底物,增強呼吸作用,提高能量代謝來抵抗低溫傷害。電鏡下觀察表明,低溫脅迫后線粒體和嵴數量增多[8]。本研究中,輕度低溫(0℃)脅迫下根細胞和葉肉細胞的線粒體和嵴數量均明顯增多,這與前人的研究結果一致。隨脅迫時間的延長,根細胞和葉肉細胞對低溫表現出明顯差異,根細胞線粒體雙層膜破裂、嵴消失的現象比葉肉細胞早發生12 h,這可能是葉肉細胞中的淀粉粒為逆境呼吸提供了更多的呼吸底物。淀粉是葡萄糖分子聚合而成的長鏈化合物,逆境下淀粉粒可以重新分解為葡萄糖,作為呼吸底物,參加三羧酸循環。而根細胞比葉肉細胞缺少了淀粉粒,這可能是導致地下部抗寒性低于地上部的一個重要因素。
膜體系的損傷是造成植物受害的根本機制[29-30]。細胞發生冰凍傷害會產生冰晶,造成細胞脫水[8],脫水時的收縮直接造成了質膜的破壞,表現為液泡破裂、原生質體濃縮、葉綠體膨大、線粒體腫脹,類囊體排列不規則,線粒體膜和嵴消失、細胞內出現空泡化等現象[31]。本研究在-3℃和-5℃脅迫下,根細胞和葉肉細胞內均結冰,導致一系列受害現象,葉綠體解體和線粒體的內嵴破壞,都是不可逆的,并最終導致植株死亡[32]。
本研究在透射電鏡下觀察表明,低溫不僅會損傷膜系統,還會對細胞壁造成破壞。細胞壁主要由纖維素、半纖維素和果膠質構成,結構高度復雜[33],細胞壁表面糖蛋白脫落到細胞間隙中或凝集成塊進入細胞內部都是導致個體間抗寒性差異的原因[31]。因細胞壁本身沒有生命跡象,所以不會因低溫而產生直接破壞。而細胞結冰就會對細胞壁造成機械傷害[8]。輕度低溫脅迫36 h,葉肉細胞壁無明顯受害癥狀,而根細胞壁明顯破壞,且先發生在細胞壁內側。從細胞壁的破壞程度和發生位置可以推測,輕度低溫脅迫引起了根細胞胞間結冰。細胞間因吸水導致體積膨脹,細胞內因缺水導致收縮,是細胞壁外側光滑而內側首先損傷的根本原因[8]。
葉片褐變率、根系活力和PMP指標可以直接反映葉片和根的凍害程度。輕度低溫脅迫12 h后,根系活力下降26.09%,而葉片褐變率僅升高17.15%,脅迫12~48 h根系PMP值持續升高而葉片PMP值升高不顯著,表明0℃下枇杷苗根部表現持續受害,而葉片則表現出較強的適應性,受害癥狀不明顯。這說明地下部較地上部對低溫更敏感。
綜上所述,枇杷苗葉肉細胞比根細胞對低溫具有更強的適應性,一方面是因為不同器官在不同環境下長期的適應性,可能導致地下部根細胞的適應性弱于地上部葉肉細胞;另一方面是與底物貯藏量有關,低溫脅迫下細胞通過增強呼吸作用提高抗逆性,而無論是三羧酸循環還是抗氰呼吸在脅迫環境下都需要消耗大量底物,枇杷苗根細胞底物的儲備量相對葉肉細胞的少,因此更容易受低溫危害。
枇杷苗根尖比葉對低溫更敏感,因此從防御策略上,對枇杷園進行生草栽培,或在極端低溫到來之前實施地表覆草、覆蓋等保溫措施,提高地溫,并結合噴灑防霜劑和植物生長調節劑類物質[10],來提高枇杷耐寒能力,以期達到更好的防寒效果。因本研究僅是對較不耐寒品種,對耐寒品種地下部與地上部低溫脅迫下的反應有待進一步研究。
