rno-miR-16-5p 靶基因預測、生物信息學分析及鑒定
2019-08-13 03:34:14王芳潘茂興蔡三金梅志剛馮知濤
實用醫學雜志 2019年14期
王芳 潘茂興,2 蔡三金 梅志剛 馮知濤,2
1三峽大學醫學院國家中醫藥管理局中藥藥理科研三級實驗室(湖北宜昌443002);2暨南大學醫學院(廣州510632)
微小RNA(microRNA,miRNA)是一類單鏈非編碼小分子RNA,通過堿基互補配對原則與特定靶基因的信使RNA(message RNA,mRNA)的3′-UTR區結合,起到抑制mRNA 翻譯或直接降解其特定mRNA 的作用[1]。miRNA 在細胞生長發育、分化、凋亡等方面扮演重要角色[2],超過50%的miRNA 位于與癌癥相關脆弱區和DNA 斷裂點上[3],且miRNA 的表達失調可參與控制脊索瘤[4]、乳腺癌[5]、結腸癌[6]等多種疾病的進程。目前許多miRNA 的靶基因尚未明確,探索其具體作用機制仍較困難,因此利用生物信息學準確預測miRNA 的靶基因,結合雙熒光素酶報告實驗進行靶基因鑒定對研究miRNA的后續作用機制顯得尤為重要。
miR-16 在研究慢性淋巴細胞白血病時被發現[7],可以通過靶向調控細胞周期素D1等基因誘導細胞周期阻滯從而參與細胞周期進程的調控[8]。miR-16 被證實為腫瘤抑制因子,它的下行調節可以影響腫瘤細胞的凋亡[9],且與類風濕關節炎的發生發展密切相關[10]。研究[11]表明,miR-16 參與重度抑郁癥的形成,提示miR-16 可能是調控神經細胞的一個重要miRNA。腦源性神經營養因子(brain-derived neurotrophic factor,BDNF)可能是miR-16 的下游靶基因之一[12],但對其確切性及具體機制的研究甚少。本實驗通過miRNA 靶基因數據庫預測rno-miR-16-5p 的靶基因,利用基因本體論(gene ontolog,GO)注釋對靶基因進行分子功能和生物學分類,京都基因與基因組百科全書(Kyoto Encyclopedia of Genes and Genomes,KEGG)軟件對靶基因富集的信號通路進行富集性分析,并構建雙熒光素酶報告實驗載體,對預測出的靶基因進行鑒定,為挖掘rno-miR-16-5p 的功能奠定實驗基礎。
1 材料與方法
1.1 實驗材料蛋白胨、酵母提取物購自OXOID(英)公司;限制性內切酶(XhoI、NotI)、Phusion DNA 聚合酶、連接反應液、普通DNA 聚合酶購自Thermo 公司;DH5α 感受態細胞、Universal DNA Purification Kit DNA 純化回收及TIANprep Mini Plasmid Kit 質粒小提試劑盒均購自天根生化科技(北京)有限公司;瓊脂糖購自BioWest(西班牙)公司;氨芐青霉素購自sigma 公司;Dual-Glo?Luciferase Assay System 購于Promega(美國)公司;LipofectamineTM2000 購于Invitrogen(美國)公司;DEME 培養基與胰酶購于GIBCO(美國)公司。
1.2 實驗方法
1.2.1 rno-miR-16-5p 靶基因預測利用miRanda(http://www.microrna.org)、miRDB(http://www.mirdb.org)、TargetScan(http://www.targetscan.org)3 個數據庫預測rno-miR-16-5p 的靶基因,取三者結果的交集以減少假陽性。
1.2.2 GO 分析與KEGG 通路分析將預測到的靶基因集合集進行GO 和KEGG 分析。GO 分為生物過程(biological process,BP),分子功能(molecular function,MF)和細胞組件(cellular component,CC)3 個層面,并利用超幾何分析檢驗法進行統計學分析,以P <0.01 為顯著性閾值分別得到GO 注釋顯著性分析。KEGG 通路分析利用Fisher Exact Test 計算P 值,以P <0.05 得到差異具有統計學意義的疾病通路及信號轉導。
1.2.3 雙熒光素酶報告載體的構建通過美國國家生物技術信息中心(National Center of Biotechnology Information,NICB)基因庫查找大鼠BDNF 基因3′-UTR序列(NM_012513),應用軟件設計擴增引物,引物序列為:r-BDNF_3′-UTR_F:GGCGGCTCGAGTGTTGCCGTTGCCAAGAAT;r-BDNF_3′-UTR_R:AATGCGGCCGCCCTGACCCATGCCAGAAGA。下劃線為XhoI、NotI 的酶切位點序列,預計引物擴增長度為573 bp。引物由蘇州金唯智生物科技有限公司合成。以C6 基因組DNA 為模板,采用30 μL 反應體系進行PCR 擴增,1.5%瓊脂糖電泳分析,限制性內切酶XhoI、NotI 雙酶切上述純化產物和pmiR-RBREPORTTM 載體,酶切后回收產物進行純化、連接、轉化,挑取轉化后的5 個菌落進行PCR 驗證,條帶理論大小為850 bp。
1.2.4 細胞的瞬時轉染取對數生長期的293T 細胞接種于96 孔板中(每孔1.5 × 104細胞,總體積100 μL),37°C培養箱中培養24 h。取10 μL OPTIMEM 培養基稀釋miRNA mimic(miRNA 模擬物,試驗組)或Non-target Control(NC,陰性對照組),15 μL OPTI-MEM 培養基稀釋BDNF 基因3′-UTR雙熒光素酶報告基因載體,25 μL OPTI-MEM 培養基稀釋0.25 μL LipofectamineTM2000 試劑,混勻后靜置20 min。96 孔板中每孔吸走50 μL 培養基后加入上述50 μL 混合液,每組設3 個復孔。6 h 后再加100 μL 新鮮培養基。
1.2.5 rno-miR-16-5p 靶基因的驗證細胞轉染后48 h,加入1 ×PBS 與luciferase 底物(35 μL 每孔),震蕩10 min 后測定螢火蟲熒光值;再加入Stop reagent(30 μL),震蕩10 min 后測定海腎熒光值。
1.3 統計學方法實驗數據應用SPSS19.0 軟件進行統計分析,數據以均數±標準差表示,采用t 檢驗進行兩組間比較,P <0.05 為差異具有統計學意義。
2 結果
2.1 靶基因的預測結果采用TargetScan、miRDB、miRanda 數據庫預測出的靶基因分別有1 633、737、640 個,為減少假陽性,取三者交集得到的靶基因有48 個(圖1)。
2.2 靶基因GO 分析結果對以上48 個預測靶基因進行GO 分析,在BP 層面,靶基因主要富集于細胞生長發育、促進突觸合成及神經系統發育等與神經細胞有關的GO 條目中(P <0.01)。在CC 層面,靶基因主要富集于血小板α 顆粒、力蛋白復合體等GO 條目中(P <0.01)。在MF 層面,靶基因主要富集于粘多糖聚集GO 條目中(P <0.01)。其中在符合條件的GO 條目中幾乎所有BP 層面的基因功能注釋均有BDNF 出現(圖2)。
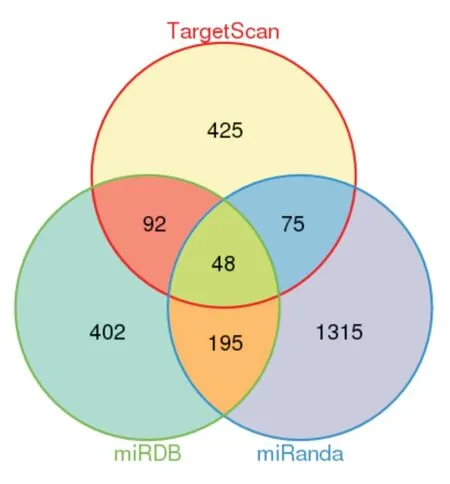
圖1 TargetScan、miRDB、miRanda 對rno-miR-16-5p 靶基因的預測結果Fig.1 The prediction results of rno-miR-16-5p target genes of TargetScan,miRDB and miRanda
2.3 靶基因集KEGG 通路分析結果在GO 注釋分類的基礎上,通過對靶基因集合的KEGG 分析發現靶基因主要富集于4個生物學通路中(P <0.05),分別為唾液分泌通路(Salivary secretion)、mTOR信號通路(mTOR Signaling pathway)、cAMP 信號通路(cAMP Signaling pathway)與胰腺分泌通路(Pancreatic secretion),其中BDNF 參與cAMP 信號通路(圖3)。
2.4 BDNF 基因3′-UTR 區雙熒光素酶基因報告載體的構建通過PCR 擴增BDNF 基因3′-UTR 片段,得到長度為573 bp 的片段,與預期相符(圖4)。隨后經雙酶切試驗,選取5 個菌落進行菌落PCR 鑒定,結果發現,挑取的5 個單克隆有4 個長度為850 bp 為陽性克隆,與預期一致(圖5)。表明雙熒光素酶報告實驗載體構建成功。
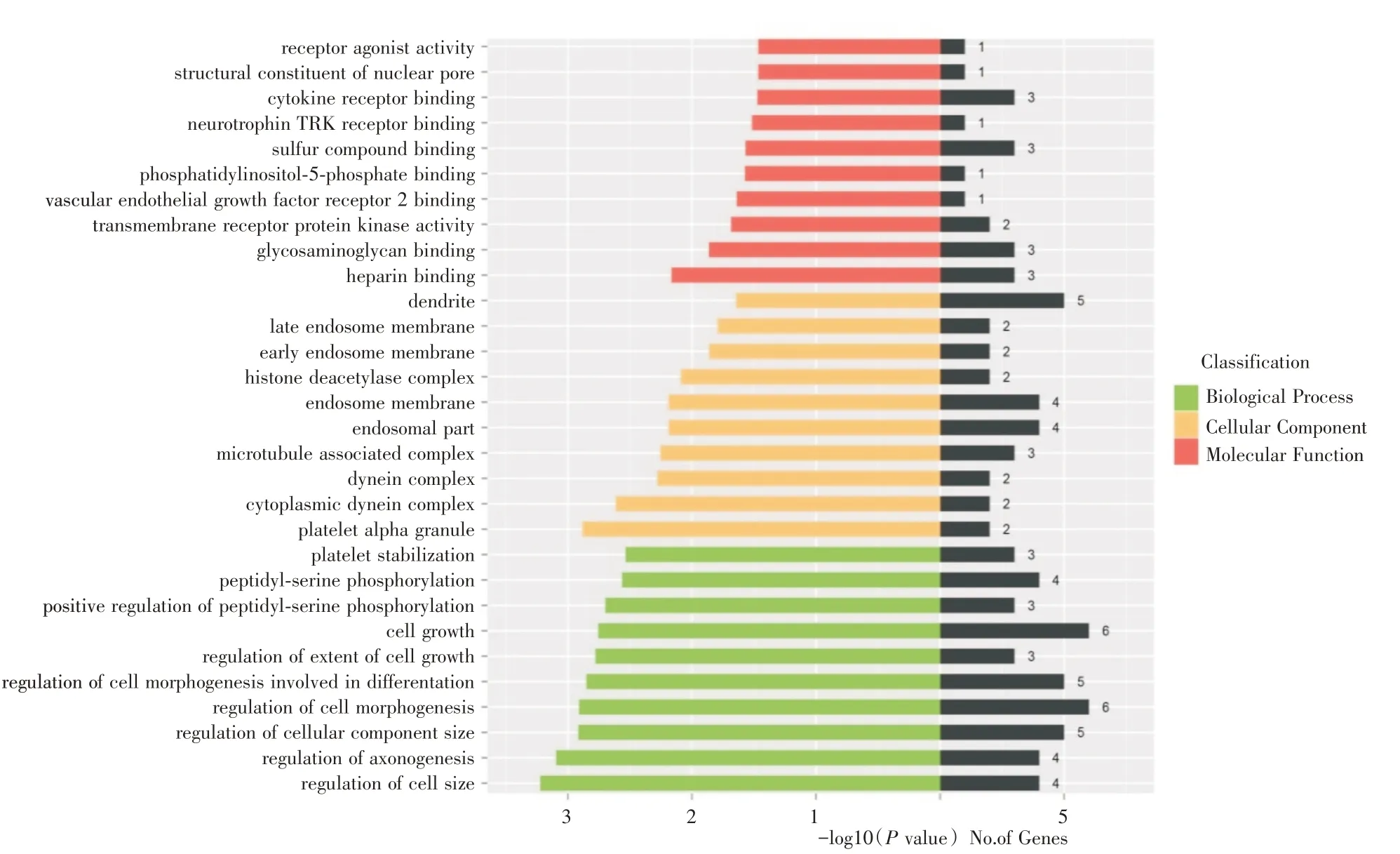
圖2 rno-miR-16-5p 預測靶基因集GO 分類圖Fig.2 GO terms of predicted target genes of rno-miR-16-5p
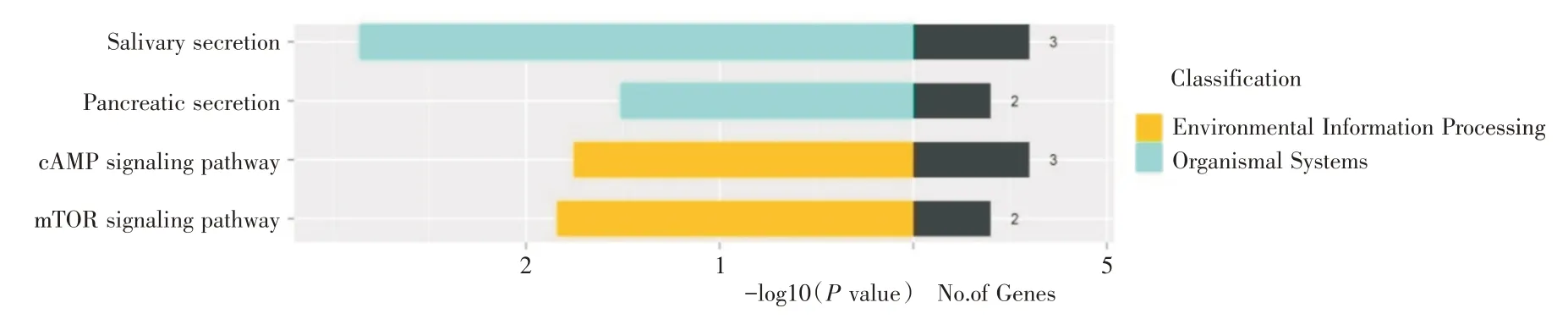
圖3 rno-miR-16-5p 預測靶基因KEGG 分類圖Fig.3 KEGG terms of predicted target genes of rno-miR-16-5p
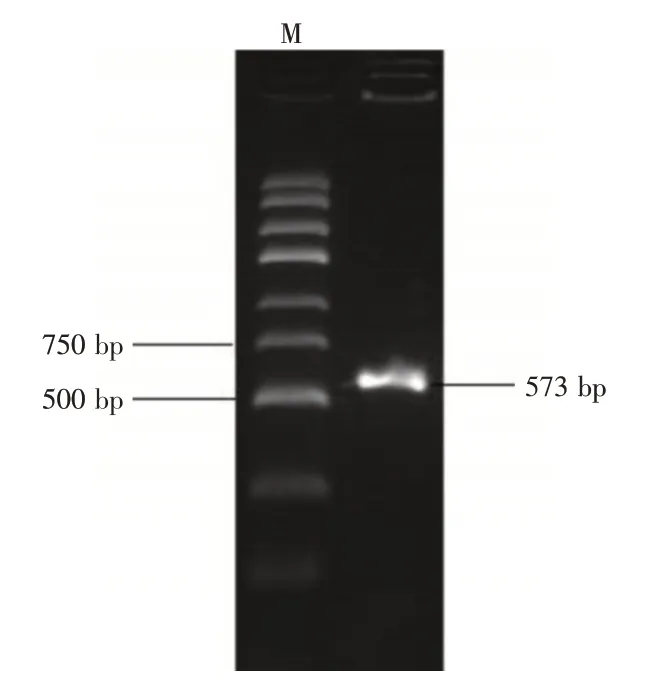
圖4 BDNF 基因3′-UTR 序列PCR 電泳結果Fig.4 PCR amplification product of BDNF gene 3′-UTR
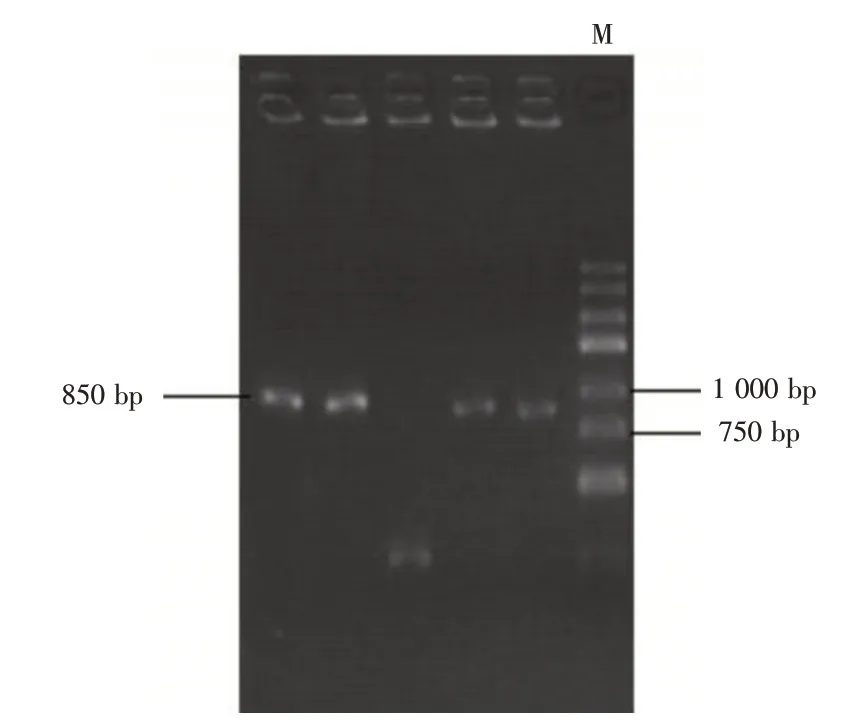
圖5 轉化菌落PCR 電泳分析結果Fig.5 PCR amplification product of transformed colony
2.5 rno-miR-16-5p 的靶基因鑒定為了檢測rnomiR-16-5p 對BDNF 基因3′-UTR 區的調控作用,對熒光素酶的活性進行了檢測。結果顯示,rno-miR-16-5p mimic 對BDNF 野生型的報告熒光有顯著的下調作用(t = 48.99,P = 0.000),提示BDNF 是rnomiR-16-5p 的靶基因(圖6)。
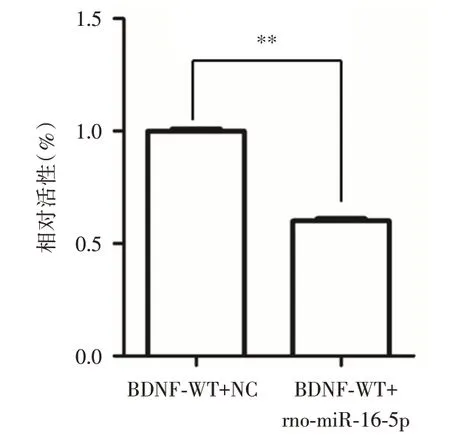
圖6 雙熒光素酶報告結果Fig.6 The results of dual luciferase reporter gene assays
3 討論
miR-16 在腫瘤、神經系統疾病等疾病中發揮重要作用。miR-16 的下調可通過孕激素介導的致癌基因信號促進乳腺癌的發展[13];miR-16 通過靶向肝癌衍生生長因子從而抑制非小細胞肺癌中腫瘤細胞的增殖與侵襲[14]。研究[11]發現,miR-16對人類和嚙齒動物神經系統功能相關的基因表達進行多效調節,導致“抗應激”的行為表現;miR-16的表達水平與膠質母細胞干細胞的分化呈正相關,但對膠質母細胞瘤細胞的運動性有負面影響[15],并且miR-16 可作為抑郁癥等疾病的早期診斷指標[16],但是miR-16 在神經功能調節中的具體機制鮮有文獻報道。本實驗利用TargetScan、miRDB、miRanda 數據庫預測rno-miR-16-5p 的靶基因分別有1 633、737、640 個,取三者的交集靶基因共48 個,包括BDNF、VEGFA 等,具有高特異性及低假陽性率,其中miR-16 可通過抑制多發性骨髓瘤中VEGFA 的表達來調節抗腫瘤免疫[17],而有關rno-miR-16-5p 靶基因之一BDNF 的文獻資料較少,因此對靶基因BDNF 的生物信息學分析及鑒定具有一定的意義。
為了進一步揭示rno-miR-16-5p 的生物學功能,本實驗對預測的靶基因集進行了GO 與KEGG分析,結果顯示,rno-miR-16-5p 靶基因功能富集于細胞生長發育、促進突觸合成及神經系統發育等生物學過程;血小板α顆粒、力蛋白復合體等細胞組件;粘多糖聚集等分子功能。rno-miR-16-5p 靶基因集通路富集于唾液分泌通路、mTOR 信號通路、cAMP 信號通路與胰腺分泌通路。其中BDNF參與了上述幾乎所有的生物學過程和cAMP 信號通路,提示BDNF 可能通過cAMP 信號通路調控多個生物學過程。cAMP 信號通路又稱為蛋白激酶A 系統(protein kinase A system,PKA),是環核苷酸系統的一種,可通過活化cAMP 依賴的PKA 使下游靶蛋白磷酸化,從而影響細胞代謝和細胞行為[18]。研究[19]表明,PKA 磷酸化的α-氨基羥甲基惡唑丙酸(α-amino-3-hydroxy-5-methyl-4-isox-azolepropionate,AMPA)受體亞基對突觸可塑性有重要作用,而AMPA 受體可介導中樞神經系統興奮性突觸傳遞,且PKA 可促進短期記憶轉化為長期記憶來儲存[20],提示cAMP 信號通路可能是一條重要的與神經系統功能相關的通路。
BDNF 是哺乳動物中樞神經系統中最豐富的神經營養因子,對神經元的存活、分化、生長發育起重要作用[21]。研究[22]發現BDNF 水平下降與癲癇患者認知障礙有關,且是抗抑郁藥效的基本決定因素[23]。為了驗證BDNF 是否為rno-miR-16-5p的靶基因之一,本研究成功構建了野生型雙熒光素酶報告實驗載體,并檢測熒光素酶的活性來觀察rno-miR-16-5p 對BDNF 基因3′-UTR 區的調控作用,結果顯示,rno-miR-16-5p mimic 組報告熒光顯著減少,表明BDNF 是rno-miR-16-5p 的靶基因。本研究存在不足之處,在后續研究工作中會進一步完善關于突變型BDNF 基因3′-UTR 區雙熒光素酶基因報告,進一步證實rno-miR-16-5p 與靶基因BDNF 的調控關系。
綜上所述,BDNF 是rno-miR-16-5p 的靶基因之一,結合GO 與KEGG 分析結果,提示rno-miR-16-5p 可能靶向作用于BDNF,再可能通過參與cAMP信號通路調控細胞生長發育、監管軸突生成、促進神經系統發育等生物學過程,但其具體的分子機制尚未明確,需進一步探討。
猜你喜歡
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
鴨綠江(2021年35期)2021-04-19 12:24:18
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
電子制作(2018年18期)2018-11-14 01:48:24
電子制作(2018年11期)2018-08-04 03:25:42
山東工業技術(2016年15期)2016-12-01 05:31:22
發明與創新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
