江蘇鹽城原生濱海濕地土壤細菌群落多樣性分析
2019-08-13 08:54:10趙婧祝遵凌
江蘇農業科學 2019年2期
關鍵詞:群落結構
趙婧 祝遵凌
摘要:以江蘇鹽城原生濱海濕地蘆葦灘、蘆葦堿蓬灘、堿蓬灘、堿蓬米草灘、互花米草灘、青蛤灘這6種典型灘面為研究對象,采用高通量測序方法測定土壤細菌群落的功能多樣性,利用冗余分析細菌群落與環境因子的關系,以探討濱海濕地土壤細菌群落結構空間分布及環境影響因子。結果表明,濱海濕地土壤理化指標有明顯差異,由蘆葦灘到青蛤灘,土壤pH值呈逐漸上升趨勢,土壤有機碳(SOC)、全氮(TN)、銨態氮(NH+4-N)、硝態氮(NO3--N)含量呈逐漸降低趨勢,土壤全磷(TP)含量變化趨勢不明顯;濱海濕地30個土壤樣品高通量測序共得到50 172條高質量序列,平均長度為8 362 bp,有97%相似,土壤樣品文庫的覆蓋率范圍為98.04%~99.56%;不同土壤細菌群落豐度指數與Shannon指數排序一致,為蘆葦灘>蘆葦堿蓬灘>堿蓬灘>堿蓬米草灘>互花米草灘>青蛤灘,豐度相對較高的細菌門為變形菌門、放線菌門、酸桿菌門,豐度值分別為27.26%~33.06%、23.47%~29.03%、15.03%~19.02%,主要的優勢菌綱為α-變形菌綱、放線菌綱、酸桿菌綱;由蘆葦灘到青蛤灘,土壤細菌Chao、Ace、Shannon指數、OTU數量呈逐漸降低趨勢,土壤細菌覆蓋率、Simpson指數沒有明顯規律性變化;土壤pH值、SOC含量、NH4+-N含量是濱海濕地土壤細菌群落結構和多樣性的主要影響因子。
關鍵詞:濱海濕地;土壤細菌;群落結構;江蘇鹽城;蘆葦灘;青蛤灘
中圖分類號: S154.38+1 ?文獻標志碼: A ?文章編號:1002-1302(2019)02-0258-04
整個生態圈中,陸地和江河湖泊占據了大部分生態區域,而濕地作為二者的過渡地帶,其獨特的生態環境和生物群落分布使之備受關注,處于水、土交界的地帶存在著較強的生物生產力,對碳、氮等元素的轉化、循環起著重要的促進作用,成為地表元素運動的重要途徑之一[1-3]。微生物作為重要的生物群落之一,是濕地物質能量交換、濕地生態的維系者,在促進濕地生態系統正常運轉方面起著不可忽視的作用[4]。微生物生長及作用的發揮受環境的影響較大,從群落結構、生長狀況等研究微生物和環境之間的關系,可為濕地元素的轉化研究奠定基礎,也有利于對濕地的管理和合理開發利用[5-6]。近年來,分子生物學發展迅速,高通量測序技術因其低成本、無需克隆、通量高等獨特優勢,在短時間內可深入研究微生物的群落特征,甚至稀有物種的生長發育狀況[7-10]。
保持土壤肥力和活性是土壤保持生命活力的前提,而土壤微生物在肥力保持方面起著不可忽視的作用,不僅能夠使腐殖質等迅速分解,轉化為土壤養分來源,而且能夠疏松土壤、增強土壤孔隙度等,對有機化合物的分解、碳氮循環等起著無可替代的作用,同時,能夠促進能量的轉化循環[11]。有研究表明,微生物作為土壤質量的重要指標之一,其多樣性和活性對土壤肥力起著重要的作用[12-13];細菌作為微生物群落的重要組成部分,占微生物類群的70%,且遺傳多樣性較為豐富,能促進土壤有機質分解及大量營養物質釋放,在碳、氮循環過程中起著重要的促進作用,參與整個物質循環和能量交換[14-16]。
我國面積最大、生物多樣性明顯的淤泥質濱海濕地為鹽城濱海濕地。20世紀80年代,江蘇人民在沿海進行大規模圍墾,全省近70%的圍墾集中在鹽城地區。隨著人為干預的增強,濱海濕地發生巨大變化,其土壤類型和物種多樣性也發生巨大變化[17]。李龍山等研究發現,濱海地區植物交替明顯,景觀呈現典型的動態變化,土壤覆被類型也在不斷變化[18]。但是,濕地微生物群落經歷什么變化,這些變化與土壤之間關系怎樣,如何合理利用濱海濕地資源等等,有關這些研究相對較少。本研究從鹽城濱海濕地出發,探究微生物特點與土壤之間的關系,分析細菌多樣性對土壤帶來的影響、作用機制及其微生物群落的生長發育特點,為鹽城濱海濕地土壤資源的合理開發利用提供有益參考。
1 材料與方法
1.1 研究區概況
江蘇省鹽城濱海濕地屬典型的季風氣候特征,受到大陸氣候和海洋季風的影響,具有較為獨特的氣候特點,年均降水量近1 000 mm,最冷月份在1月,平均氣溫約為3 ℃,最熱月集中在6—8月,平均氣溫達到30 ℃左右,9月至翌年2月并不寒冷。鹽城濱海濕地核心區地處新洋河和斗龍河過渡區,該區域受到人為的干擾并不明顯,依然保持較為完好的生態環境,呈現典型的濱海生態特點,具有差異明顯的分帶性,能從不同層次反映濕地的生態環境,從陸地向濱海延伸分布著蘆葦灘、堿蓬灘、互花米草灘,同時還因青蛤廣布而形成青蛤灘,對濕地研究具有很好的代表性。
1.2 土樣采集
于2016年11月2日枯水期,采用“S”形5點取樣法,分別從濱海濕地分層地帶采集0~10 cm土層的土壤,一部分用于測定土壤理化性狀,另一部分提取DNA用于測序。
1.3 測定內容和方法
1.3.1 土壤理化性狀 分別采用氯化鉀溶液提取-分光光度法(HJ 634—2012)、堿熔-鉬銻抗分光光度法(HJ 632—2011)測定土壤總氮(TN)、總磷(TP)含量;采用氯化鉀溶液提取-分光光度法(HJ 634—2012)測定銨態氮(NH4+-N)、硝態氮(NO3--N)含量。
1.3.2 細菌16S rRNA基因測序 采用Omega試劑盒、引物F27提取16S rDNA宏基因組,采用ABI Gene Amp 9700型PCR儀進行擴增,試驗采用Trans Gen AP 221-02,20 μL反應體系為:5×fast pfu buffer 4 μL,2.5 mmol/L dNTPs 2 μL,5 μmol/L 正反向引物各0.8 μL,Fast Pfu聚合酶0.4 μL,TemplateDNA 10 ng,加H2O至20 μL。反應參數為:95 ℃預處理210 s; 55 ℃ 30 s,72 ℃ 45 s,27次循環;72 ℃延伸 10 min。采用AxyPrep DNA凝膠回收,進行Tris_HCl洗脫處理,電泳檢測。采用Illumina MiseqPE 300平臺測序。
1.4 生物信息處理
對原始DNA通過Mothur軟件過濾處理,去除其中的嵌合體;以97%相似性為依據劃分可操作分類單元;通過稀釋性曲線分析獲取豐度、覆蓋度、多樣性指數,通過主成分分析獲取運算分類單元(OTU)相似性;采用貝葉斯算法,獲取不同分類水平上的樣品群落特點,整理OTU列表獲得分類信息與豐度,在門和綱分類水平下進行冗余分析(RDA),得到細菌群落組成結構及其與環境因子的關系。
2 結果與分析
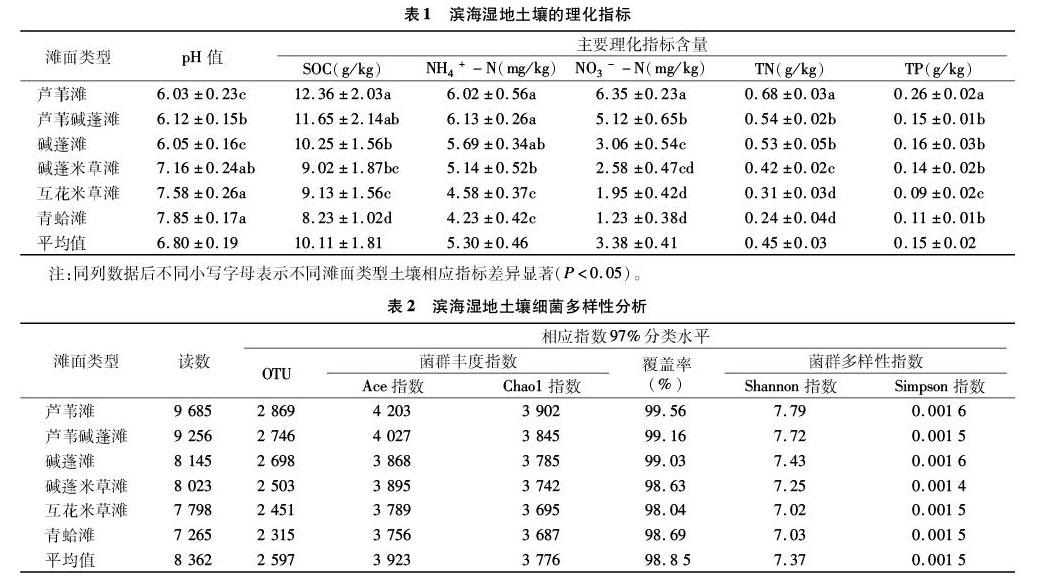
2.1 濱海濕地土壤理化指標
由表1可知,濱海濕地土壤pH值在6.03~7.85之間,平均值為6.80,由蘆葦灘到青蛤灘,土壤pH值呈逐漸增加趨勢;土壤有機碳(SOC)含量在8.23~12.36 g/kg之間,平均值為10.11 g/kg,由蘆葦灘到青蛤灘,SOC呈逐漸降低趨勢;從蘆葦灘到青蛤灘,NH4+-N、NO3--N含量呈逐漸降低趨勢,在蘆葦灘出現最大值,分別為6.02、6.35 mg/kg,NH4+-N、NO3--N平均含量分別為5.30、3.38 mg/kg;TN含量變化趨勢與SOC相一致,從蘆葦灘到青蛤灘呈逐漸降低趨勢,其平均值為0.45 g/kg;TP含量變化趨勢不明顯,總體上呈遞減趨勢,其平均值為0.15 g/kg。
2.2 濱海濕地土壤細菌的多樣性
由表2可知,濱海濕地30個土壤樣品的高通量測序共得到50 172條高質量序列,平均長度為8 362 bp,97%相似;各樣品文庫的覆蓋率為98.04%~99.56%,說明土樣中基因序列被檢出的概率相對較高,測序結果能夠代表濕地土壤細菌群落的真實情況。Chao1指數用于估算樣品中所含OTU總數,反映菌群豐度,Chao1指數越大,說明細菌群落豐度越高;Shannon指數反映細菌群落alpha多樣性指數,Shannon指數值越大,說明細菌群落多樣性越高。由表2可知,不同土壤細菌群落豐度指數與Shannon指數排序一致,為蘆葦灘>蘆葦堿蓬灘>堿蓬灘>堿蓬米草灘>互花米草灘>青蛤灘,從蘆葦灘到青蛤灘,土壤細菌Chao1、Ace、Shannon指數、OTU數量呈逐漸降低趨勢,土壤細菌覆蓋率、Simpson指數變化沒有明顯規律性變化。
2.3 濱海濕地土壤細菌門水平分類和綱水平分類
2.3.1 門分類水平 由圖1可知,沿湖面至坡地,土壤的門分類細菌相對豐度存在一定的變化趨勢,濱海濕地土壤相對豐度較高的細菌門為變形菌門、 放線菌門、酸桿菌門、綠彎菌門、芽單胞菌門、厚壁菌門、綠菌門、硝化螺旋菌門,相對豐度值分別為27.26%~33.06%、23.47%~29.03%、15.03%~19.02%、7.13%~13.58%、5.13%~9.87%、2.16%~5.12%、1.54%~4.23%、0.66%~4.87%;由蘆葦灘到青蛤灘,變形菌門相對豐度大致呈先增大后減小趨勢,酸桿菌門大致呈增大趨勢,綠彎菌門大致呈先減小后增大趨勢,綠菌門沒有明顯變化規律。
2.3.2 綱分類水平 由圖2可知,濱海濕地土壤檢測到的主要綱有α-變形菌綱、放線菌綱、酸桿菌綱、β-變形菌綱、嗜熱油菌綱、芽單胞菌綱、桿菌綱、δ-變形菌綱,相對豐度值分別為26.06%~29.78%、18.30%~29.30%、15.02%~21.05%、8.26%~15.06%、5.32%~8.32%、3.12%~8.30%、1.45%~8.14%、1.21%~7.27%,其中,前6個綱在土壤中占主導地位,約占所有綱的80%。
2.4 濱海濕地土壤理化性質與土壤細菌群落結構的相關性
土壤可以提供細菌群落生長繁殖的微環境,而不同的植被類型可通過改變土壤的微環境間接影響土壤細菌群落結構的組成。由表3可知,變形菌門、酸桿菌門、厚壁菌門相對豐度與土壤pH值呈顯著負相關(P<0.05),綠彎菌門相對豐度與土壤pH值呈極顯著負相關(P<0.01);變形菌門、酸桿菌門、芽單胞菌門、厚壁菌門、硝化螺旋菌門相對豐度與SOC含量呈顯著正相關,綠彎菌門相對豐度與土壤SOC含量呈極顯著正相關;酸桿菌門、芽單胞菌門、厚壁菌門、硝化螺旋菌門相對豐度與NH4+-N含量呈顯著正相關,變形菌門、放線菌門相對豐度與NH4+-N含量呈極顯著正相關;變形菌門、酸桿菌門、綠彎菌門、綠菌門、綠菌門相對豐度與NO3--N含量呈顯著正相關,芽單胞菌門相對豐度與NO3--N含量呈極顯著正相關;變形菌門、綠彎菌門、厚壁菌門、綠菌門相對豐度與TN含量呈顯著正相關,芽單胞菌門相對豐度與TN含量呈極顯著正相關;不同細菌門相對豐度與TP含量相關性不顯著(P>0.05)。
2.5 濱海濕地環境因素對土壤細菌群落結構的影響
由圖3可知,對土壤理化環境因子與細菌主要菌落(門)所占比重進行中心標準化,并進行校正,提取第一、第二坐標軸信息,對方差總解釋率分別為61.25%、23.58%;第一主軸上,SOC、NH4+-N含量是主要的影響因子,第二主軸上,SOC含量、pH值是主要的影響因子。因此,與濱海濕地土壤細菌群落相關性較大的土壤理化指標為pH值、SOC含量、NH4+-N 含量。
3 結論與討論
生物的多樣性對維系整個生態平衡具有舉足輕重的作用,微生物群落的多樣性對促進整個生態循環同樣具有重要的意義,不僅有利于營養鹽的循環,且在有機物降解、養分轉化方面發揮著重要作用[19]。本研究發現,蘆葦灘菌群的豐度、多樣性相對最高,其次是蘆葦堿蓬灘、堿蓬灘,而堿蓬米草灘、互花米草灘的豐度及多樣性并不明顯,而青蛤灘微菌群分布相對最少;土壤菌群的豐度、多樣性、OTU數量等從陸地向濱海地帶多呈遞降趨勢,這可能是距離河岸較遠的地方土壤肥力相對較強,對微生物的生長發育有利。對位置相對接近的地帶,其土壤菌群分布具有較明顯的相似性[20]。有研究發現,微生物的生長發育及活動明顯受到水文狀況的制約,干濕交替對部分細菌生長具有明顯的促進作用,與淹沒區有顯著不同[21];水位狀況及其變化直接影響微生物群落的分布,且水文條件影響下的土壤沉積差異對微生物群落的影響更為顯著[22-23]。本研究結論與之較為吻合。
微生物菌群中的變形菌門在黃河三角洲濕地具有較高的豐度[24]。對鹽城濱海濕地而言,相對豐度較高的變形菌門α、β、δ-變形菌綱中,前2類菌綱具有較多的固氮細菌,能夠明顯提升土壤的固氮水平,對土壤肥力的保持具有重要作用,而δ-變形菌綱為食菌性細菌[25],能夠明顯促進濕地養分循環;酸桿菌門相對豐度值也較高,相對豐度值15.03%~19.02%,與張瓊瓊等的研究結果[26]基本一致,但其多為未培養細菌,雖然在土壤中廣泛存在,但相關研究并不多;試驗中菌群相對豐度位于第4位的綠彎菌門,以CO2為能量來源,在光合作用下獲取能量,進而使其能夠在SOC含量較低的土壤中擁有較強的生存能力和適應性。通過冗余分析發現,SOC含量、NH4+-N含量、pH值對濕地菌群分布具有明顯的制約效應。
參考文獻:
[1]龐丙亮,崔麗娟,馬牧源,等. 若爾蓋高寒濕地生態系統服務價值評價[J]. 濕地科學,2014,12(3):273-278.
[2]張 彪,史蕓婷,李慶旭,等. 北京濕地生態系統重要服務功能及其價值評估[J]. 自然資源學報,2017,32(8):1311-1324.
[3]魏 強,佟連軍,楊麗花,等. 三江平原濕地生態系統生物多樣性保護價值[J]. 生態學報,2015,35(4):935-943.
[4]丁冬靜,李 玫,廖寶文,等. 海南省濱海自然濕地生態系統服務功能價值評估[J]. 生態環境學報,2015(9):1472-1477.
[5]李 偉,崔麗娟,龐丙亮,等. 濕地生態系統服務價值評價去重復性研究的思考[J]. 生態環境學報,2014,23(10):1716-1724.
[6]張 晶,陳書明,王小國. 天鵝湖濕地微生物群落PCR-DGGE分析[J]. 基因組學與應用生物學,2016(9):2423-2428.
[7]陳 偉. 納帕海高原濕地微生物多樣性研究[D]. 昆明:昆明理工大學,2016.
[8]魯青原. 遼河三角洲濱海濕地微生物群落組成及其環境意義[D]. 北京:中國地質大學,2016.
[9]常軍軍,吳蘇青,梁 康,等. 復合垂直流人工濕地微生物特征對典型污水的響應差異[J]. 環境科學研究,2016,29(8):1200-1206.
[10]黃興如,張瓊瓊,張瑞杰,等. 再生水補水對河流濕地香蒲根際細菌群落結構影響研究[J]. 中國環境科學,2016,36(2):569-580.
[11]王 林,李 冰,朱 健. 高通量測序技術在人工濕地微生物多樣性研究中的研究進展[J]. 中國農學通報,2016,32(5):10-15.
[12]吳文衛,劉 昂,谷照虎,等. 采用PCR-DGGE技術研究處理農田退水組合人工濕地微生物群落特征[J]. 應用與環境生物學報,2016,22(6):978-985.
[13]魏佳明,崔麗娟,李 偉,等. 表流濕地細菌群落結構特征[J]. 環境科學,2016,37(11):4357-4365.
[14]李龍山,倪細爐,李昌曉,等. 生活污水對土壤及濕地植物根際細菌群落的影響[J]. 農業環境科學學報,2016,35(11):2163-2170.
[15]王 鵬,陳 波,張 華. 基于高通量測序的鄱陽湖典型濕地土壤細菌群落特征分析[J]. 生態學報,2017,37(5):1650-1658.
[16]隋 心,張榮濤,楊立賓,等. 模擬氮沉降對三江平原小葉章濕地土壤細菌多樣性的影響[J]. 草業科學,2016,33(4):589-598.
[17]靳 亮,高學梅,杜建華,等. 尕海湖濕地泥炭細菌多樣性分析[J]. 微生物學通報,2016,43(11):2396-2404.
[18]李龍山,倪細爐,李昌曉,等. 生活污水對土壤及濕地植物根際細菌群落的影響[J]. 農業環境科學學報,2016,35(11):2163-2170.
[19]武海鵬. 生態環境修復及江湖關系改變對洞庭湖濕地土壤微生物的影響[D]. 長沙:湖南大學,2016.
[20]徐 飛,蔡體久,楊 雪,等. 三江平原沼澤濕地墾殖及自然恢復對土壤細菌群落多樣性的影響[J]. 生態學報,2016,36(22):7412-7421.
[21]靳振江,曾鴻鵠,李 強,等. 起源喀斯特溶洞濕地稻田與旱地土壤的微生物數量、生物量及土壤酶活性比較[J]. 環境科學,2016,37(1):335-341.
[22]周雪芳,王立立,劉 鈴,等. 珠江河口濕地沉積物中耐鹽硝化細菌的篩選及其河口硝化特性研究[J]. 生態環境學報,2016,25(12):1991-1998.
[23]黃 媛,褚文珂,謝蔚鵬,等. 杭州西溪濕地沉積物中不同培養基分離的細菌種群多樣性[J]. 杭州師范大學學報(自然科學版),2016,15(6):600-605.
[24]曾 靜. 錫林河濕地厭氧氨氧化菌群多樣性、豐度及空間特征研究[D]. 呼和浩特:內蒙古大學,2016.
[25]張瑞杰,張瓊瓊,黃興如,等. 再生水濕地香蒲根內生細菌群落多樣性及其水質特征分析[J]. 中國環境科學,2016,36(3):875-886.
[26]張瓊瓊,黃興如,郭逍宇. 基于T-RFLP技術的不同水位梯度植物根際細菌群落多樣性特征分析[J]. 生態學報,2016,36(14):4518-4530.
猜你喜歡
熱帶農業科學(2016年11期)2017-01-21 14:59:21
熱帶農業科學(2016年11期)2017-01-21 14:57:08
河北漁業(2015年12期)2015-12-21 14:37:31
河北漁業(2015年11期)2015-11-18 16:39:54
河北漁業(2015年11期)2015-11-18 16:37:49
河北漁業(2015年10期)2015-10-15 11:14:57
湖北農業科學(2015年18期)2015-10-13 21:34:01
河北漁業(2015年7期)2015-07-21 11:06:52
河北漁業(2015年6期)2015-06-24 11:04:37
安徽農學通報(2015年5期)2015-03-23 23:51:20
