HCV Core基因真核表達載體構(gòu)建及其對HeLa細胞自噬的影響
2019-08-16 07:43:40王敏敏魏建宏任來峰
天津醫(yī)藥 2019年7期
關(guān)鍵詞:研究
王敏敏,魏建宏,任來峰
丙型肝炎病毒(hepatitis C virus,HCV)感染是肝細胞癌(hepatocellular carcinoma,HCC)的重要風險因素[1],而HCC是最為難治的惡性腫瘤之一,也是我國位居第二的癌癥“殺手”,HCV感染約與20%的HCC相關(guān)[2]。HCV核心蛋白(Core蛋白)除了參與病毒組裝外,還能與正常細胞蛋白相互作用,擾亂細胞正常的信號轉(zhuǎn)導、轉(zhuǎn)錄調(diào)控等信號傳遞[3]。HCV Core蛋白被認為是HCV相關(guān)HCC的潛在癌基因或輔因子[4-5],有研究表明,過表達HCV Core后,在其轉(zhuǎn)基因(TG)小鼠中可以誘導肝癌的形成[6]。
自噬是一種利用溶酶體將自身細胞內(nèi)受損、變性或衰老的蛋白以及細胞器消化降解的過程,并參與細胞多種生理及病理過程調(diào)控[7],細胞自噬與腫瘤的發(fā)生發(fā)展密切相關(guān)。有研究表明,HCV Core蛋白可誘導肝細胞自噬[8-9]],而 Hara等[10]研究顯示,HCV Core蛋白可抑制HCV感染時的有絲分裂;有研究認為,這種抑制可導致線粒體泛素化、有絲分裂體形成和自噬降解的失敗[11]。有關(guān)HCV Core蛋白與自噬的關(guān)系研究尚不多,其具體機制仍有待闡明。因此,本研究擬通過在HeLa細胞中表達外源HCV Core蛋白,研究其對HeLa細胞自噬的影響,為進一步闡明HCV相關(guān)的HCC的發(fā)病機制提供參考。
1 資料與方法
1.1 主要材料 PLVX-IRES-ZsGreen1過表達質(zhì)粒和HCV復制子質(zhì)粒(pJFH1)為本實驗室保存,大腸埃希菌感受態(tài)細胞DH5α購自索萊寶生物技術(shù)公司,人宮頸癌HeLa細胞株購自豐暉生物技術(shù)公司,In-Fusion?HD Cloning Kit購自Clontech公司,凝膠回收試劑盒和質(zhì)粒抽提試劑盒均購自O(shè)mega公司。限制性內(nèi)切酶XhoⅠ購自北京百奧萊博科技有限公司,委托生工生物工程(上海)股份有限公司完成引物合成及DNA測序;DNA轉(zhuǎn)染試劑Lipofectamine 2000購自Invitrogen公司,核染料DAPI購自Vector公司,抗HCV Core抗體購自博奧森公司,實驗所用FITC標記的二抗(抗鼠)和內(nèi)參α-tubμlin抗體以及Cyc3標記的二抗(抗兔)均購自Sigma公司,抗LC3B抗體購自Cell Signaling Technology公司。
1.2 方法
1.2.1 載體的構(gòu)建 (1)目的基因擴增:按照同源重組原理設(shè)計Core基因引物,上游5′-CGGTGAATTCCTCGAATGAGCACAAATCCTAAACC-3′,下游 5′-TAGAACTAGTCTCGATCAAGCAGAGACCGGAACGGT-3′。以 pJFH1為模板進行PCR擴增,反應體系為:上、下游引物各1μL,pJFH1模板0.5μL,Taq DNA聚合酶0.5μL,dNTP混合液0.5μL,PCR Buffer 2.5μL,ddH2O 19μL。反應條件:95℃預變性3 min;95℃ 30 s,55 ℃ 30 s,72 ℃ 150 s,共34個循環(huán);72 ℃ 5 min充分延伸。PCR產(chǎn)物分離并純化,4℃保存?zhèn)溆谩#?)重組載體克隆:XhoⅠ限制性內(nèi)切酶切割空載體PLVX-IRES-ZsGreen1使之線性化,并用凝膠回收試劑盒回收純化;用IN-fusion cloning kit將
HCV Core基因片段與線性化載體進行重組;將重組質(zhì)粒轉(zhuǎn)化到含氨芐青霉素(Amp)LB培養(yǎng)平板,于37℃培養(yǎng)12 h;挑取LB轉(zhuǎn)化板上的陽性菌落,接種于15 mL(含Amp)LB液體培養(yǎng)基中,置于37℃搖床培養(yǎng)18 h(180 r/min)。次日按說明書提取質(zhì)粒,選用EcoRⅠ和BamHⅠ雙酶切鑒定重組質(zhì)粒PLVXIRES-ZsGreen1-Core,并送博奧公司測序證實。
1.2.2 細胞的傳代與培養(yǎng) 用DMEM高糖培養(yǎng)基(含10%胎牛血清和1%雙抗)在37℃、5%CO2孵箱中培養(yǎng)宮頸癌HeLa細胞,每隔1~2 d更換新鮮培養(yǎng)基,待細胞長至匯合度達85%~90%時,用胰酶消化進行傳代培養(yǎng)。
1.2.3 質(zhì)粒轉(zhuǎn)染 選取對數(shù)生長期的HeLa細胞接種于無菌24孔細胞培養(yǎng)板中,常規(guī)培養(yǎng)待細胞貼壁生長至約50%~60%匯合度時進行細胞轉(zhuǎn)染,設(shè)PLVX-IRES-ZsGreen1-Core重組質(zhì)粒轉(zhuǎn)染組、PLVX-IRES-ZsGreen1空載體轉(zhuǎn)染對照組以及未轉(zhuǎn)染質(zhì)粒的空白對照組,采用Lipofectamine 2000按試劑說明書進行操作。轉(zhuǎn)染完成6~8 h后換常規(guī)DMEM高糖培養(yǎng)基并于48~72 h后進行后續(xù)實驗。
1.2.4 免疫印跡(Western Blot)檢測Core蛋白以及LC3蛋白的表達 參照文獻[12]:轉(zhuǎn)染48 h后,棄去24孔板中的培養(yǎng)基并用預冷的PBS清洗1次,經(jīng)RIPA裂解液裂解后收集各組細胞并抽提細胞總蛋白,經(jīng)BCA蛋白定量試劑盒定量并調(diào)整蛋白濃度,按說明書加入5×loading buffer煮沸后得到蛋白樣品。總蛋白經(jīng)12%SDS-PAGE電泳分離后轉(zhuǎn)移到PVDF膜上,5%脫脂奶粉封閉1 h(搖床),分別用抗HCV Core(1∶100)、LC3B(1∶1 000)、α-tubμlin(1∶5 000)抗體4 ℃孵育過夜,TBST漂洗;再用HRP標記的抗兔和抗鼠二抗(1∶2 000)37℃孵育1 h,TBST漂洗,將膜用ECL顯色,F(xiàn)luor Chem FC2成像儀曝光并分析結(jié)果,實驗重復3次。
1.2.5 免疫熒光(IF)檢測LC3水平 參照文獻[13]:HeLa細胞接種到24孔板中的細胞爬片上并轉(zhuǎn)染相應質(zhì)粒48 h后直接收樣;PBS洗滌1次,4%多聚甲醛室溫固定10 min,PBS漂洗后;用0.3%Triton X-100室溫通透10 min,PBS漂洗;用免疫熒光封閉液(0.1%Triton X-100+1%BSA)室溫封閉30 min以上,加入抗LC3B或抗Core抗體37℃濕盒中孵育30 min,用PBS洗3次;37℃濕盒中孵育熒光素標記的抗兔或抗鼠二抗30 min,PBS洗3次;DAPI封片,熒光顯微鏡拍片記錄結(jié)果,實驗重復3次。
1.3 統(tǒng)計學方法 采用SPSS 17.0軟件進行統(tǒng)計學分析,符合正態(tài)分布的計量數(shù)據(jù)以均數(shù)±標準差(±s)表示,2組間均數(shù)比較采用t檢驗,以P<0.05為差異有統(tǒng)計學意義。
2 結(jié)果
2.1 PLVX-IRES-ZsGreen1-Core重組質(zhì)粒的構(gòu)建及鑒定 重組載體用BamHⅠ和EcoRⅠ雙酶切結(jié)果顯示,酶切后的重組質(zhì)粒為2個條帶,大小分別與空載體PLVX-IRES-ZsGreen1和目的基因Core近似,見圖1A。測序結(jié)果示,與標準序列比對,重組質(zhì)粒PLVX-IRES-ZsGreen1-Core序列完全正確,見圖1B。
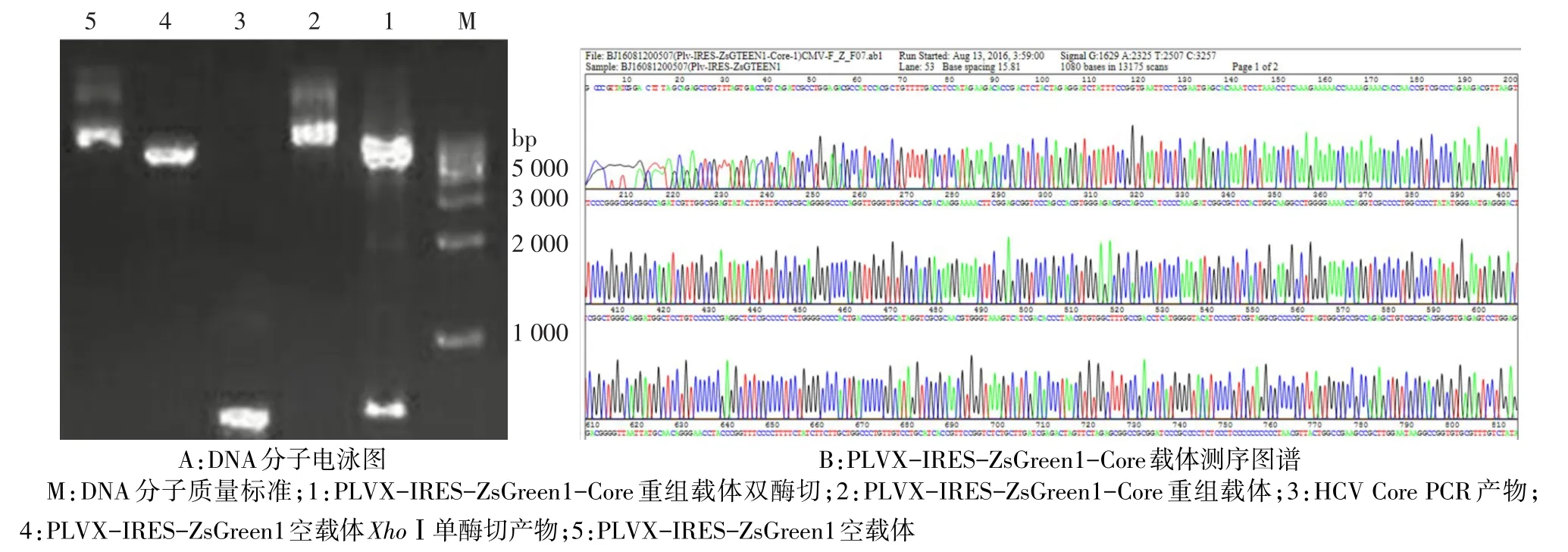
Fig.1 Core-Green digested with EcoRⅠand BamHⅠ圖1 PLVX-IRES-ZsGreen1-Core重組質(zhì)粒的鑒定
2.2 PLVX-IRES-ZsGreen1-Core重組質(zhì)粒在HeLa細胞中的表達 只有PLVX-IRES-ZsGreen1-Core重組質(zhì)粒轉(zhuǎn)染組的細胞可表達HCV Core蛋白,而PLVX-IRES-ZsGreen1空載體轉(zhuǎn)染組和未轉(zhuǎn)染質(zhì)粒空白對照組的無蛋白表達,且與Core蛋白的相對分子質(zhì)量符合,見圖2。
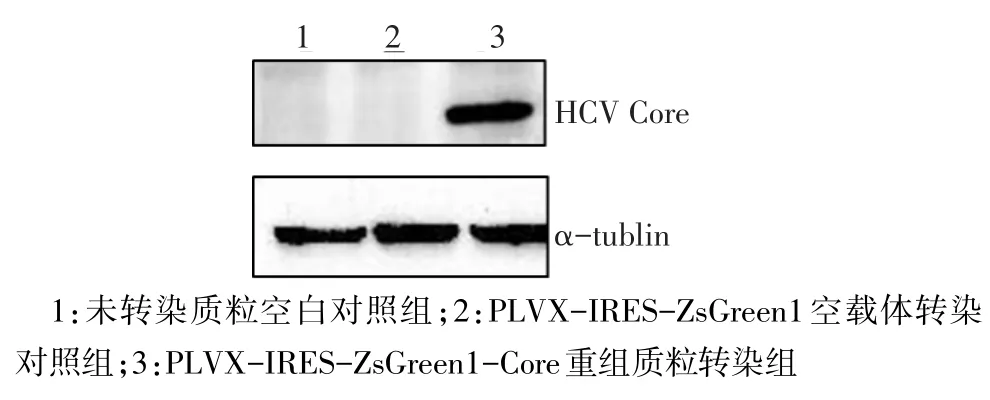
Fig.2 Expression of HCV Core protein in HeLa cells圖2 HCV Core蛋白在Hela細胞中的表達
2.3 PLVX-IRES-ZsGreen1-Core重組質(zhì)粒誘導HeLa細胞自噬 LVX-IRES-ZsGreen1-Core重組質(zhì)粒轉(zhuǎn)染組的細胞LC3Ⅱ水平(1.069±0.049)高于PLVX-IRES-ZsGreen1空載體轉(zhuǎn)染對照組(0.776±0.047),差異有統(tǒng)計學意義(t=7.433,P<0.05),見圖3。免疫熒光染色結(jié)果顯示,與Western Blot結(jié)果一致,HCV Core蛋白高表達的細胞(綠色熒光)LC3焦點明顯增多,且主要分布于細胞質(zhì)中,見圖4。
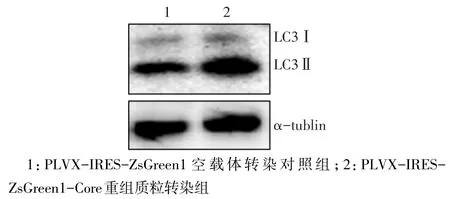
Fig.3 Expression of LC3 detected by Western Blot assay圖3 Western Blot法檢測LC3蛋白表達
3 討論
基因克隆是分子生物學的基本技術(shù),與傳統(tǒng)的基于雙酶切的分子克隆技術(shù)比較,近年來發(fā)展起來的基于同源重組的分子克隆技術(shù)(又稱為無縫克隆),具有簡便、快速、高效和適用范圍廣等優(yōu)點[14]。本實驗運用同源重組克隆方法構(gòu)建了PLVX-IRESZsGreen1-Core真核表達載體,并經(jīng)雙酶切和基因測序證實,重組DNA序列正確,重組質(zhì)粒轉(zhuǎn)染人宮頸癌HeLa細胞后可表達HCV Core蛋白,這些結(jié)果證實已成功構(gòu)建了HCV Core真核表達載體。
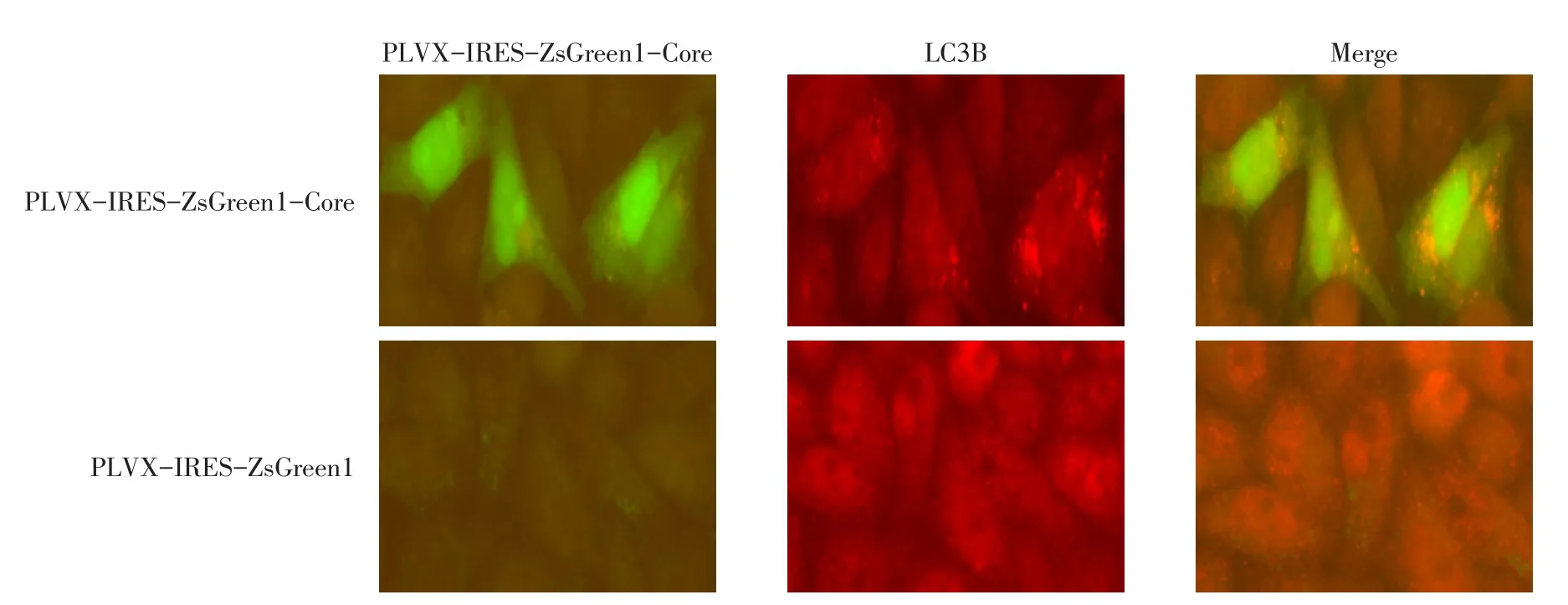
Fig.4 The changes of LC3 punctuates induced by HCV Core(×1 000)圖4 HCV Core蛋白誘導的LC3焦點變化(×1 000)
研究表明,HCV Core蛋白是誘導肝癌發(fā)生的最重要蛋白之一[15],可通過干擾宿主細胞多條信號通路誘導細胞轉(zhuǎn)化[16],但其具體致癌細節(jié)仍需進一步研究。自噬是細胞適應環(huán)境變化的一種生理病理過程,以細胞質(zhì)內(nèi)出現(xiàn)自噬體為特征,也是吞噬降解細胞內(nèi)部分受損細胞器或變性蛋白的過程[17]。在自噬發(fā)生時,微管相關(guān)蛋白1輕鏈3(microtubuleassociated protein1 light chain3,LC3)定位于前自噬體和自噬體表面,參與自噬體的整個形成過程,LC3有Ⅰ型和Ⅱ型,自噬發(fā)生時伴隨著LC3Ⅰ向LC3Ⅱ的轉(zhuǎn)化,因此,該轉(zhuǎn)化過程成為評價細胞自噬的重要參考[18-19]。自噬與腫瘤的發(fā)生發(fā)展密切相關(guān),Liu等[20]研究表明,HCV Core蛋白可誘導肝QSG-7701細胞自噬,并伴隨著LC3Ⅱ升高;Wang等[21]在肝癌細胞Huh7中的研究表明,HCV Core蛋白通過EIF2AK3和ATF6促進細胞自噬;而Hara等[10]研究則顯示,HCV Core蛋白可抑制細胞線粒體自噬。本研究中,Western Blot結(jié)果顯示,PLVX-IRES-ZsGreen1-Core重組質(zhì)粒轉(zhuǎn)染組自噬標記分子LC3Ⅱ表達增加,免疫熒光結(jié)果顯示LC3Ⅱ熒光焦點增多,證實HCV Core蛋白可提高HeLa細胞自噬水平。另有研究表明,HCV誘導細胞自噬對病毒自身的復制至關(guān)重要[22],同時細胞自噬信號的擾亂勢必干擾細胞正常生理過程,可能是其引起細胞惡性轉(zhuǎn)化的促進因素,但具體細節(jié)仍待進一步研究。
綜上所述,本研究用無縫克隆方法構(gòu)建PLVXIRES-ZsGreen1-Core重組質(zhì)粒,并可在轉(zhuǎn)染的HeLa細胞中誘導細胞自噬,為進一步探索HCV Core蛋白干擾宿主細胞功能奠定了一定的基礎(chǔ)。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設(shè)計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務(wù)財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
