低氧或亮氨酸干預對運動心肌Sestrin2/AMPKα2信號通路激活的影響
2019-08-21 00:56:06楊赟王蘊紅王智強
中國運動醫學雜志 2019年7期
關鍵詞:模型
楊赟 王蘊紅 王智強
1 首都體育學院研究生部(北京100191)
2 首都體育學院運動科學與健康學院
Sestrin2 是應激誘導蛋白Sestrins 家族中的重要成員,在機體受到應激刺激,如DNA 損傷、氧化應激、缺氧等情況下誘導產生[1]。AMPK(Adenosine Monophosphate Activated Protein Kinase,磷酸腺苷活化蛋白激酶)作為一種重要的蛋白激酶參與多種代謝過程,其活性主要受AMP/ATP 比值調控。當機體處于應激狀態如運動、組織缺血、缺氧等,都可以使AMP/ATP 比值升高,啟動AMPK激酶系統,刺激葡萄糖轉運和脂肪酸氧化等增加ATP 產生的代謝過程,同時阻斷脂類、蛋白質合成以及糖異生等需消耗ATP 的代謝通路[2]。有研究發現細胞Sestrin2 的表達可顯著提高能量狀態感受因子AMPK Thr172位點磷酸化水平[3],且4周和6周跑臺運動能引起小鼠骨骼肌Sestrin2蛋白表達和AMPKα 2 磷酸化水平顯著增加,提示運動可通過Sestrin2 激活AMPKα2信號分子[4]。也有研究表明,低氧應激能夠顯著提高人神經母細胞瘤細胞Sestrin2 蛋白表達[5]。Sestrin2 是一種高度特異的亮氨酸傳感器,機體內的亮氨酸能夠通過Sestrin2 調控mTORC1 信號通路[6,7]。但目前有關運動心肌以及低氧/亮氨酸干預能否通過調控運動心肌Sestrin2/AMPKα2 信號通路來調控心肌能量代謝的研究還未見報道。為此,本實驗通過建立單純運動模型、低氧運動模型和運動補充亮氨酸模型,探討Sestrin2/AMPKα2 信號通路在運動心肌中的時相變化以及低氧或亮氨酸干預時運動心肌Sestrin2/AMPKα2信號通路的表達規律。
1 對象與方法
1.1 研究對象
8 周雄性SPF 級SD 大鼠,由北京維通利華實驗動物技術中心提供(動物合格證號SCXK(京)2015-0001)。以國家標準嚙齒類動物飼料喂養,自由進食和飲水。動物飼養環境為溫度23℃~25℃,相對濕度在41%~53%。自然晝夜節律變化,光照和黑暗交替各12 h(光照時間在8:00-20:00)。
1.2 建立單純運動模型
1.2.1 動物分組
本實驗分為急性耐力運動模型和4 周運動模型,80只大鼠適應性飼養后隨機分組。急性耐力運動模型分為安靜組(Con)、運動后0小時取材組(E0)、運動后3小時取材組(E3)、運動后24小時取材組(E24)。4周耐力運動模型分為安靜組(4Con)、運動后0 小時取材組(4E0)、運動后3小時取材組(4E3)、運動后24小時取材組(4E24)。每組10只。
1.2.2 運動方案
急性耐力運動模型運動方案參照Bedford[8]漸增負荷跑臺運動建立模型,只進行一次耐力運動。運動方案為:初始坡度5°,速度10 m/min,每運動5 min 速度增加5 m/min。15 min 后速度增加至19 m/min,繼續運動45min。4周運動模型訓練方案:適應性訓練周坡度由5°遞增至10°,速度為14 m/min,運動時間由15 min 遞增至25 min。訓練周從第1 周至第4 周坡度保持10°,速度由14 m/min 逐漸遞增到24 m/min。運動時間從第1 周到第3 周由30 min 緩慢遞增到60 min,并保持到訓練結束。動物每周訓練5天,休息2天。
急性耐力運動模型和4周運動模型在運動后按時間點取材,安靜組同時取材。大鼠取材前12小時開始禁食,腹腔注射麻醉,打開胸腔取心臟,用預冷生理鹽水清洗,濾紙吸凈血液和生理鹽水,心臟迅速稱重,分離左心室,液氮凍存。所有操作均在冰上完成,樣本于-80℃冰箱保存。
1.3 建立低氧運動模型
1.3.1 動物分組
60 只大鼠適應性飼養后,隨機分為:常氧安靜組(Con)、常氧耐力運動組(En)、常氧力竭運動組(Ex)、低氧安靜組(Hypo)、低氧耐力運動組(Hypo+En)和低氧力竭運動組(Hypo+Ex)。每組10只。
1.3.2 運動方案
低氧運動模型中的常氧組在正常氧含量下安靜放置或運動,低氧組在北京體育大學動物房低氧房模擬4500米海拔氧環境(氧含量約11.8%)下放置或運動,低氧安靜組在低氧環境下安靜放置,低氧運動組在低氧環境下運動。運動方案參照Leandro[9]漸增負荷耐力運動方案:坡度10°,初始速度5 m/min,運動4 min后速度增加5 m/min。之后每運動3 min速度增加5 m/min。耐力運動組終速度為40 m/min,總運動時間為25 min。力竭運動組速度遞增到50 m/min后運動至力竭。
低氧運動模型所有運動組在運動后即刻取心臟,安靜組同時取材。取材方法同實驗一。
1.4 建立運動補充亮氨酸模型
1.4.1 動物分組
本實驗分為補充亮氨酸急性耐力運動模型和4周運動補充亮氨酸模型,80 只大鼠適應性飼養后隨機分組。急性耐力運動補充亮氨酸模型分為安靜組(Con)、單純運動組(E)、單純補充亮氨酸組(Leu)和補充亮氨酸運動組(Leu+E)。4 周運動補充亮氨酸模型分為安靜組(4Con)、單純運動組(4E)、單純補充亮氨酸組(4Leu)和運動補充亮氨酸組(4Leu+E)。每組10只。
1.4.2 運動方案
急性耐力運動補充亮氨酸模型運動方案同實驗一急性耐力運動模型運動方案,4周運動補充亮氨酸模型運動方案同實驗一的4周運動模型訓練方案。
1.4.3 亮氨酸灌胃方案
補充亮氨酸模型的灌胃量參照Campos[10]補充亮氨酸大鼠模型,依據《實驗動物科學》中動物與人的每公斤體重劑量折算系數表以及人類每天亮氨酸需要量計算出大鼠每天亮氨酸的需求補給量為88 mg/g,據此計算出大鼠每日灌胃量灌胃。
急性耐力運動補充亮氨酸模型大鼠在運動前10 min進行灌胃,安靜對照組和單純運動組灌胃等體積蒸餾水。補充亮氨酸組灌胃166 mg/kg,濃度83 mg/ml的亮氨酸溶液。4 周運動補充亮氨酸模型大鼠在每次運動后10 min進行灌胃,安靜對照組和單純運動組灌胃等體積蒸餾水。補充亮氨酸組灌胃166 mg/kg,濃度83 mg/ml 亮氨酸溶液。每周五下午結束末次運動后,稱量記錄體重并計算亮氨酸補劑量。
急性耐力運動補充亮氨酸模型的運動組在運動后即刻取材,不運動組同時取材。4周運動補充亮氨酸模型的運動組在運動后24 小時取材,不運動組同時取材。取材方法同實驗一。
1.5 Western Blot
稱取一定量心肌組織,按重量:體積比1∶9加入預冷的細胞裂解液,冰上勻漿,靜置20 min,4℃,13000 rpm 離心20 min。取上清BCA 法蛋白定量。以裂解液調整蛋白濃度,加入5X 上樣緩沖液,使樣品終濃度為4 μg/μl。95℃,煮沸10 min 后,制成電泳上樣液。配置12%的SDS-PAG分離膠和5%濃縮膠,加樣電泳,電泳后將蛋白轉至NC 膜。將膜在一抗中孵育,4℃過夜,一抗溶于封閉液,Sestrin2(1∶1000,兔抗,abcam 公司)、AMPKα2(1∶1000,兔抗,天德悅公司)、p-AMPKα(Thr172)(1∶2000,兔抗,abcam 公司)、β-actin(1∶20000,鼠抗,天德悅公司)。二抗使用山羊抗兔IgG(H+L),HRP(1∶5000,天德悅公司)和山羊抗鼠IgG(H+L),HRP(1∶10000,天德悅公司),室溫孵育40 min。洗膜后采用ECL發光試劑與膜在暗室中反應曝光、顯影、定影。膠片掃描后,運用ImageJ軟件處理后,使用Tota-Lab Quant軟件系統進行圖像分析,計算目的蛋白和內參蛋白條帶灰度值,并用目的蛋白的灰度值與相應的內參蛋白灰度值對比,得出每個樣品目的蛋白的相對含量。
1.6 統計學方法
實驗所得數據使用SPSS 進行雙因素方差分析(Two-way ANOVA),數據以平均數±標準差表示,對運動和各干預因素(時間、低氧和亮氨酸)進行主效應及兩者的交互作用分析。如果主效應顯著(P<0.05)則采用Tukey’s test 進行事后多重比較分析(post-hoc test)。P<0.05 表示有顯著性差異。
2 實驗結果
2.1 耐力運動對心肌Sestrin2 表達和AMPKα2 激活的影響
2.1.1 急性耐力運動對心肌Sestrin2蛋白表達的影響
與安靜組相比,運動后0 h 組心肌Sestrin2 蛋白含量顯著升高(P<0.05),在運動后3 h逐漸恢復到安靜水平。表明急性耐力運動能夠提高心肌Sestrin2 蛋白表達(見圖1)。
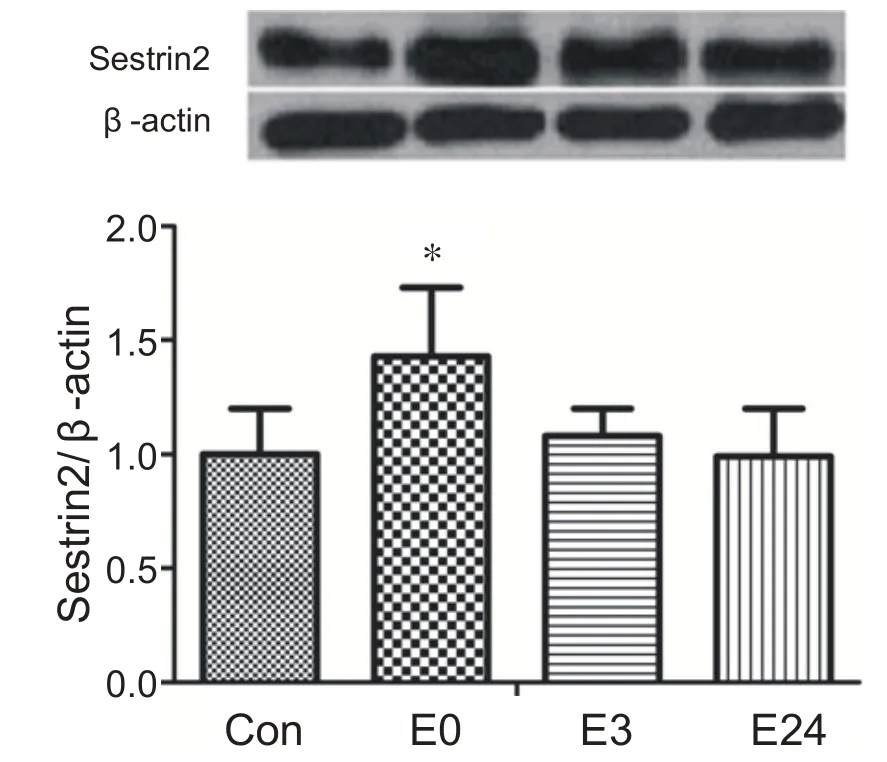
圖1 急性耐力運動后不同時間心肌Sestrin2蛋白表達
2.1.2 急性耐力運動對心肌AMPKα2激活的影響
與安靜組相比,運動后0 h組心肌AMPKα2蛋白活性顯著升高(P<0.05),在運動后3 h逐漸恢復到安靜水平。表明急性耐力運動能夠顯著激活AMPKα2(見圖2)。
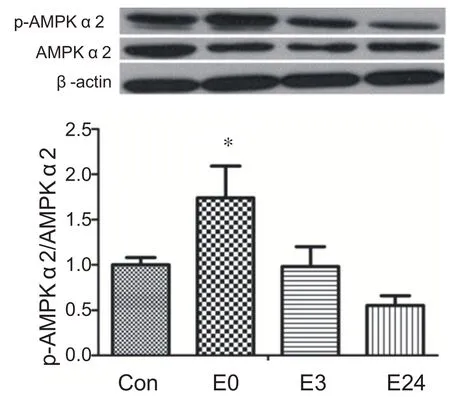
圖2 急性耐力運動后不同時間心肌AMPKα2蛋白活性
2.1.3 4周運動對心肌Sestrin2蛋白表達的影響
經過4 周運動訓練,運動后各時間段心肌Sestrin2蛋白含量與安靜組相比沒有顯著性差異(P>0.05)。說明4周運動訓練后心肌Sestrin2蛋白表達在運動后沒有顯著性變化(見圖3)。
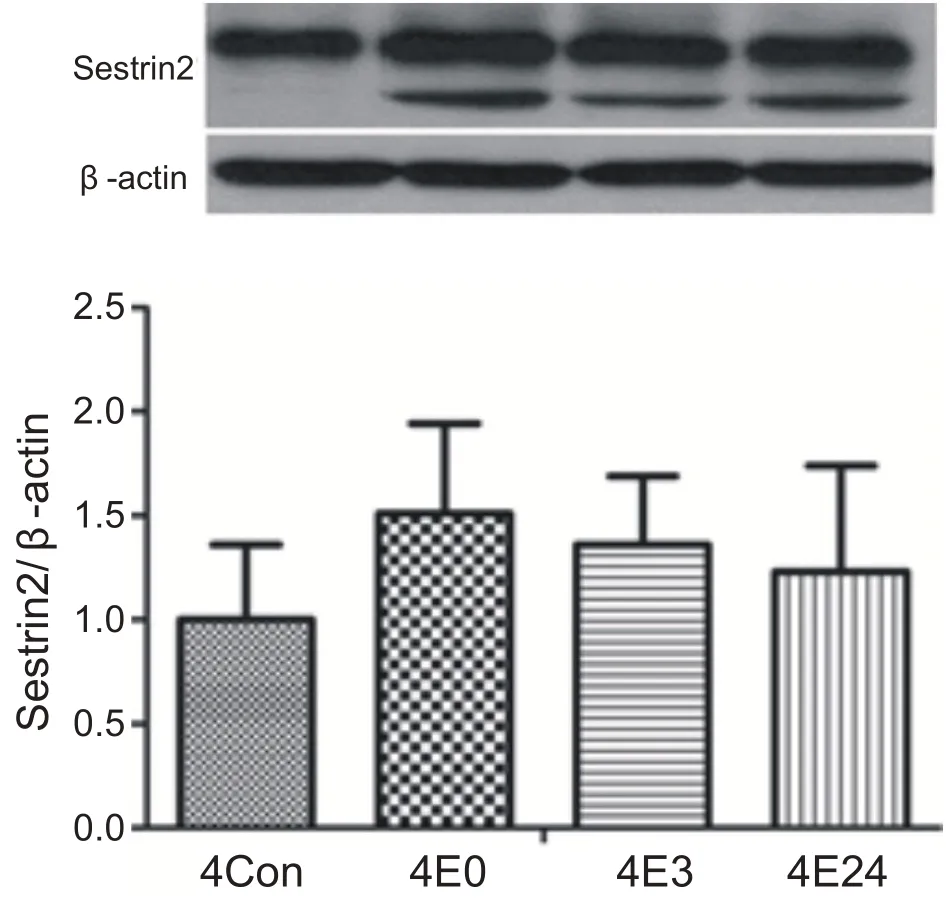
圖3 4周運動后不同時間心肌Sestrin2蛋白表達
2.1.4 4周運動對心肌AMPKα2激活的影響
經過4周運動訓練,運動后各時間段心肌AMPKα2蛋白活性與安靜組相比沒有顯著性差異(P>0.05)。說明4周耐力運動沒有激活心肌AMPKα2(見圖4)。
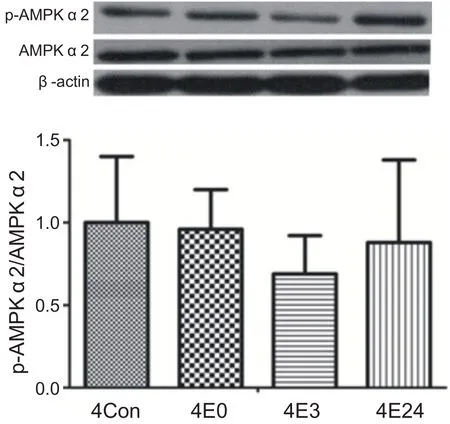
圖4 4周運動后不同時間心肌AMPKα2蛋白活性
2.2 低氧運動對心肌Sestrin2 表達和AMPKα2 激活的影響
2.2.1 低氧力竭運動對心肌Sestrin2蛋白表達的影響
與安靜組相比,常氧力竭運動組和低氧力竭運動組在運動后即刻心肌Sestrin2蛋白含量顯著性增高(P<0.05)。低氧安靜組和安靜組相比心肌Sestrin2 蛋白含量沒有顯著性差異。表明力竭運動和低氧力竭運動能夠提高心肌Sestrin2蛋白表達(見圖5)。
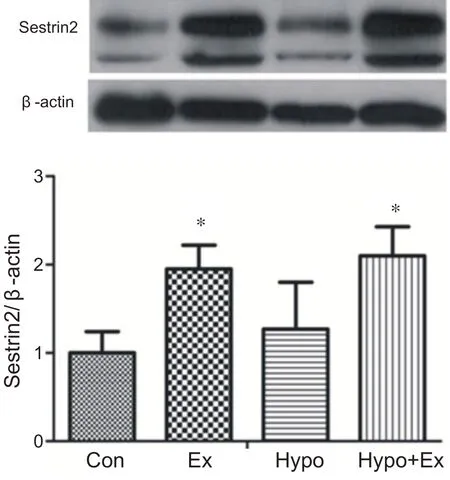
圖5 低氧力竭運動心肌Sestrin2蛋白的表達
2.2.2 低氧力竭運動對心肌AMPKα2激活的影響
與安靜組相比,常氧力竭運動組和低氧力竭運動組在運動后即刻心肌AMPKα2 蛋白活性顯著性增高(P<0.05)。低氧安靜組與安靜組相比心肌AMPKα2蛋白活性沒有顯著性變化。說明力竭運動和低氧力竭運動能夠顯著激活心肌AMPKα2(見圖6)。
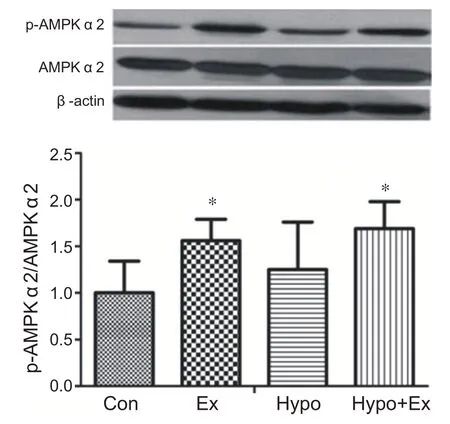
圖6 低氧力竭運動對心肌AMPKα2的影響
2.2.3 低氧耐力運動對心肌Sestrin2蛋白表達的影響
與安靜組相比,常氧耐力運動組、低氧安靜組和低氧耐力運動組心肌Sestrin2 蛋白含量沒有顯著性變化(P>0.05)。說明耐力運動和低氧耐力運動對心肌Sestrin2表達沒有影響(見圖7)。
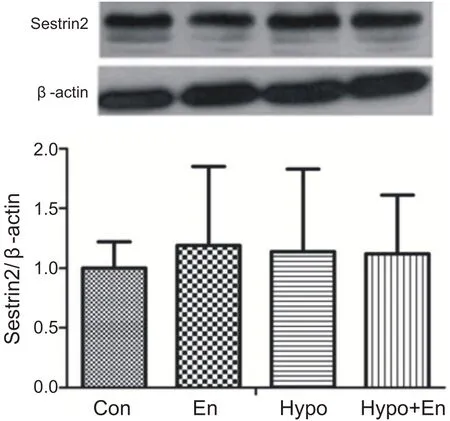
圖7 低氧耐力運動心肌Sestrin2蛋白的表達
2.2.4 低氧耐力運動對心肌AMPKα2激活的影響
與安靜組相比,常氧耐力運動組、低氧安靜組和低氧耐力運動組心肌AMPKα2蛋白激活水平沒有顯著性變化(P>0.05)。表明耐力運動和低氧耐力運動沒有激活心肌AMPKα2(見圖8)。
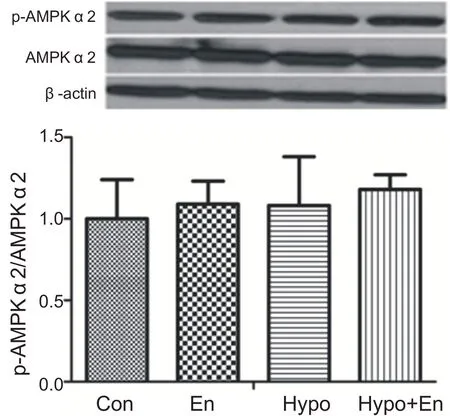
圖8 低氧耐力運動運動對心肌AMPKα2激活的影響
2.3 運動補充亮氨酸對心肌Sestrin2 表達和AMPKα2激活的影響
2.3.1 補充亮氨酸急性運動對心肌Sestrin2蛋白表達的影響
與安靜組相比,單純運動組心肌Sestrin2蛋白含量顯著升高(P<0.05)。補充亮氨酸急性耐力運動組與安靜組和補充亮氨酸安靜組相比Sestrin2 蛋白含量沒有出現顯著性變化(P>0.05)。說明補充亮氨酸急性耐力運動對心肌Sestrin2表達沒有影響(見圖9)。
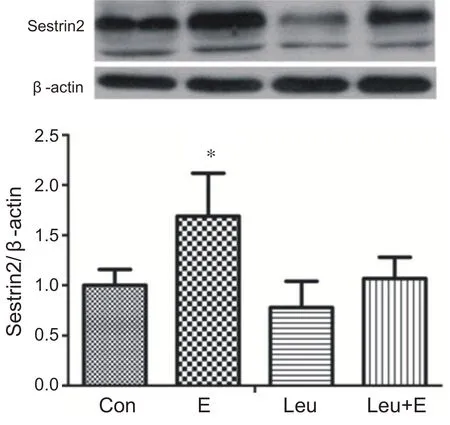
圖9 補充亮氨酸急性耐力運動對心肌Sestrin2蛋白表達的影響
2.3.2 補充亮氨酸急性運動對心肌AMPKα2激活的影響
與安靜組相比,單純運動組心肌AMPKα2 活性顯著升高(P<0.05)。單純補亮氨酸組和補亮氨酸運動組與安靜組相比心肌AMPKα2活性沒有出現顯著性變化(P>0.05)。表明補充亮氨酸急性耐力運動沒有激活心肌AMPKα2(見圖10)。
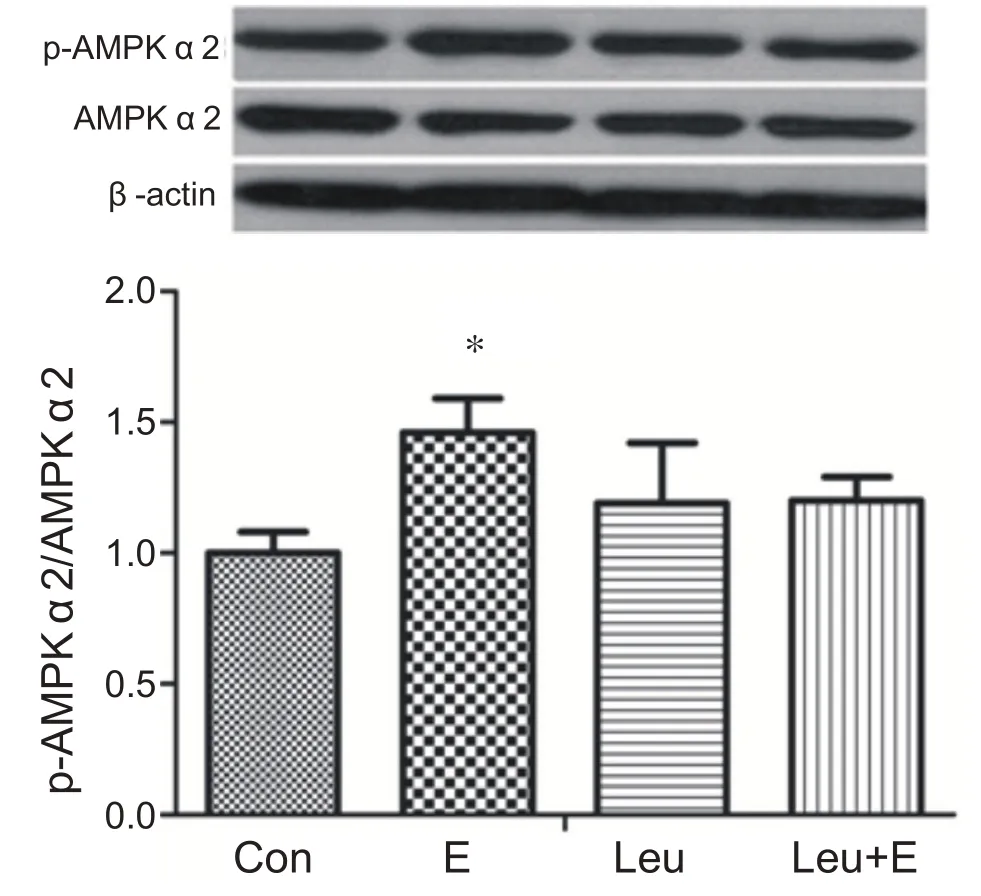
圖10 急性耐力運動補充亮氨酸對心肌AMPKα2激活的影響
2.3.3 4周運動補充亮氨酸對心肌Sestrin2蛋白表達的影響
經過4周運動訓練和補亮氨酸,單純運動組、補亮氨酸安靜組和補亮氨酸運動組與安靜組相比心肌Sestrin2蛋白含量沒有顯著性變化(P>0.05)。表明4周運動補充亮氨酸對心肌Sestrin2蛋白表達沒有影響(見圖11)。
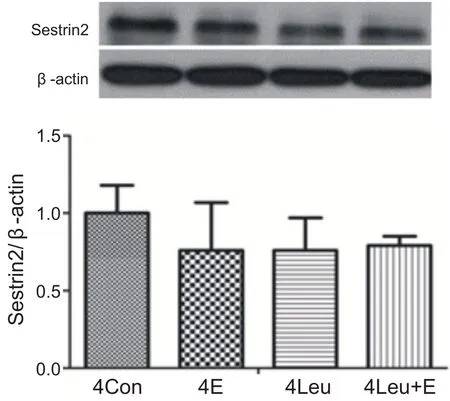
圖11 4周運動補充亮氨酸對心肌Sestrin2蛋白表達的影響
2.3.4 4周運動補充亮氨酸對心肌AMPKα2激活的影響
經過4周運動訓練和補亮氨酸,單純運動組、補亮氨酸安靜組和補亮氨酸運動組與安靜組相比心肌AMPKα2 蛋白活性沒有顯著性變化(P>0.05)。表明4周運動補充亮氨酸沒有激活心肌AMPKα2(見圖12)。
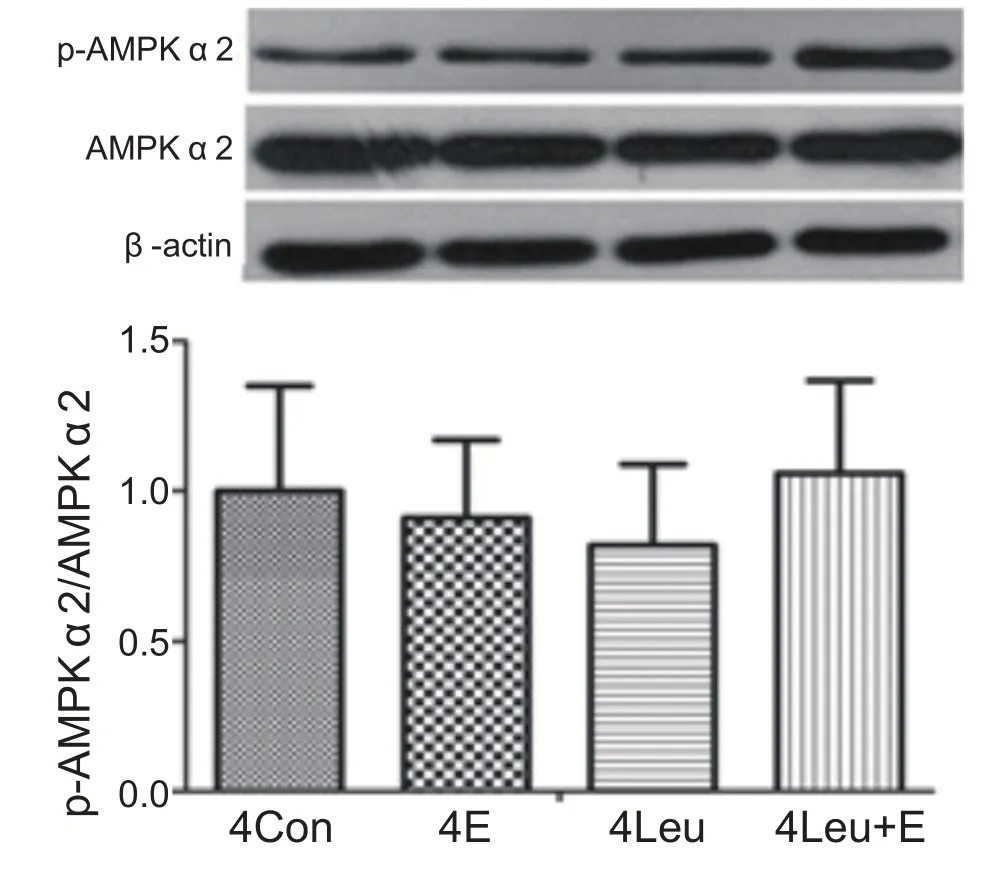
圖12 4周運動補充亮氨酸對心肌AMPKα2激活影響
3 討論
3.1 耐力運動對心肌Sestrin2/AMPKα2 信號通路激活的影響
研究發現,細胞Sestrin2表達的升高可顯著提高細胞能量狀態感受因子AMPK Thr172 位點磷酸化水平[3]。也有研究發現,急性耐力運動能夠顯著激活心肌AMPKα2[11]。但有關急性耐力運動對心肌Sestrin2/AMPKα2信號通路的影響研究還未見報道。本研究發現,運動后即刻心肌Sestrin2 蛋白表達顯著升高,說明急性耐力運動作為一種應激能夠提高心肌Sestrin2 蛋白表達。同時在急性耐力運動模型中發現運動后即刻心肌AMPKα2 活性也顯著升高,提示急性耐力運動中心肌能通過Sestrin2 激活AMPKα2 調節心肌能量代謝。而運動后3 小時心肌Sestrin2 的表達和AMPKα2的激活水平恢復到安靜水平,說明運動對心肌Sestrin2/AMPKα2信號通路的激活是一個短期的激活。
在本實驗中發現經過四周的運動訓練,心肌Sestrin2的表達和AMPKα2的激活水平在運動后各時間點沒有顯著性差異,提示經過四周耐力運動訓練心肌Sestrin2/AMPKα2信號通路表現出對運動的適應性改變。
3.2 低氧運動對心肌Sestrin2/AMPKα2 信號通路激活的影響
心肌供能以有氧代謝為主,對低氧刺激比較敏感。急、慢性低氧能使心肌糖原減少,葡萄糖代謝增強,乳酸增加,脂肪酸代謝增強等[12]。有研究表明,低氧暴露能使心肌AMPKα2磷酸化水平顯著增加[13]。還有研究顯示,低氧應激能夠顯著提高人神經母細胞瘤細胞Sestrin2蛋白表達[5]。但有關低氧應激以及低氧環境運動能否引起心肌Sestrin2 表達和AMPKα2 活性改變還未見報道。本實驗結果顯示,低氧力竭運動和常氧力竭運動均能使心肌Sestrin2 蛋白表達和AMPKα2活性顯著升高,但兩組之間沒有顯著性差異,并且單純低氧對心肌Sestrin2/AMPKα2信號通路沒有明顯影響,表明低氧干預對心肌Sestrin2/AMPKα2 信號通路的激活作用不明顯。而低氧耐力運動組心肌該信號通路激活也沒有發現變化,進一步說明低氧環境下運動誘發心肌該信號通路的變化只與運動強度和時間有關,而與缺氧環境無明顯的聯系。盡管發生的機制尚不清楚,但有研究顯示,大鼠頸總動脈分別結扎150 min 或100 min,造成兩個不同程度(嚴重或中等)的缺氧模型,結果發現,只有嚴重的缺氧模型Sestrin2的表達顯著升高,而中等程度缺氧模型Sestrin2表達沒有變化[14],表明缺氧不是刺激Sestrin2 表達所必需的因素。另有研究顯示,一些降低細胞ATP濃度的物質,如2-脫氧葡萄糖(糖酵解抑制劑)和二甲雙胍(線粒體呼吸抑制劑),也能夠誘導Sestrin2表達[15],提示Sestrin2表達與能量供應下降有關。此外,Sestrin2的表達雖然在一些細胞是受缺氧誘導因子(HIF-1)調控,而在另一些細胞類型,缺氧所誘導的Sestrin2 表達不依賴HIF-1,并且顯示出與其他HIF-1靶基因不同的表達動力學[5,16]。從上述研究結果可以得出,Sestrin2的轉錄不是由缺氧本身所誘導,而可能是由缺氧引起的能量缺乏所致。因此可以推測,在本實驗中,單純低氧暴露和低氧非力竭耐力運動未引起心肌Sestrin2/AMPKα2激活,可能是由于未出現能量供應缺乏,而低氧力竭運動心肌是否出現了能量供應缺乏有待進一步研究。
3.3 運動補充亮氨酸對心肌Sestrin2/AMPKα2 信號通路激活的影響
支鏈氨基酸是用于運動員恢復的常用補劑[17],也是心臟攝取的主要氨基酸,雖然氨基酸在心臟ATP 的產生中比重較小,但它們對蛋白質合成和細胞信號傳導等過程至關重要。其中亮氨酸是重要的供能氨基酸,能調節糖代謝和蛋白質代謝[18]。近來的研究顯示,Sestrin2是一種高度特異的亮氨酸傳感器[7]。當亮氨酸缺乏時,Sestrin2 與蛋白復合體GATOR2 交互作用,抑制mTORC1信號通路激活和細胞生長。而亮氨酸存在時,能夠直接結合Sestrin2,破壞這種交互作用,使mTORC1 信號分子激活[6]。但有關補充亮氨酸對心肌Sestrin2/AMPKα2 信號通路作用的研究還未見報道。為此,本研究通過給予運動大鼠口服亮氨酸,觀察補充亮氨酸對運動心肌Sestrin2/AMPKα2 信號通路的急性作用和長期的效應。由于實驗一結果已經顯示,急性耐力運動后即刻心肌Sestrin2 的表達和AMPKα2 活性提高,因此我們觀察了運動前補充亮氨酸的大鼠急性運動后即刻該信號通路的變化,結果顯示,心肌Sestrin2的表達和AMPKα2活性與安靜組相比沒有顯著性變化,而未補充亮氨酸的急性耐力運動組心肌Sestrin2/AMPKα2通路活性顯著性升高。提示補充亮氨酸能夠抑制運動對心肌Sestrin2/AMPKα2信號通路的刺激作用。
以往的研究表明,運動使AMPKα2 激活后可能通過磷酸化TSC2 和抑制mTOR 的活性降低p70S6K 的磷酸化,進而抑制蛋白合成,促進能量底物分解供能[19,20]。AMPKα2 活性的增加還可以抑制mTOR 上游信號分子Akt 的活性[21]。而Sestrins 能通過增強AMPKα2 活化,抑制mTORC1活性,從而使細胞合成代謝的抑制和分解代謝過程如β-氧化和自噬的增加[1]。我們以往的研究發現,4 周的遞增負荷耐力訓練能引起Akt/mTOR信號通路的持續性激活,為此,本實驗通過建立4周耐力訓練模型,觀察長期耐力運動后給與亮氨酸對Sestrin2/AMPKα2通路的作用,結果顯示,經過4周運動結合補充亮氨酸的大鼠心肌Sestrin2 表達和AMPKα2 活性均與安靜對照組相比沒有顯著差異。在本實驗中,4周運動訓練的大鼠Sestrin2/AMPKα2 通路的活性也與安靜對照組沒有顯著差異,并且我們在相同的模型上還觀察到mTOR 的活性和心重/體重也沒有明顯變化。這提示,所采用的4 周運動方案訓練大鼠心肌與蛋白質合成相關的信號通路沒有激活,而補充氨基酸也沒有對Sestrin2/AMPKα2/mTOR 信號通路產生影響。是由于補充亮氨酸只對運動應激刺激引起的Sestrin2 表達升高起抑制作用,還是運動心肌對一段時間補充亮氨酸產生了適應性改變,還不十分清楚。由于目前還沒有Sestrin2 的磷酸化抗體,因此有關補充亮氨酸對Sestrin2的活性的影響,以及補充亮氨酸對運動心肌蛋白質合成的調控機制還有待進一步研究。
4 結論
4.1 急性耐力運動可以一過性激活心肌Sestrin2/AMPKα2信號通路,而對長期耐力訓練后心肌Sestrin2/AMPKα2信號通路表現出對運動應激的適應。
4.2 運動的負荷量與心肌Sestrin2/AMPKα2 信號通路的激活關系密切,而低氧干預對心肌Sestrin2/AMPKα2信號通路的激活作用不明顯。
4.3 補充亮氨酸可抑制急性耐力運動對心肌Sestrin2/AMPKα2信號通路的刺激作用,而經過4周運動和補充亮氨酸后的心肌Sestrin2/AMPKα2 信號通路沒有顯著性變化。
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
網絡安全與數據管理(2022年1期)2022-08-29 03:15:20
導航定位學報(2022年4期)2022-08-15 08:27:00
中學生數理化·中考版(2022年8期)2022-06-14 06:55:24
新世紀智能(數學備考)(2021年9期)2021-11-24 01:14:36
成都醫學院學報(2021年2期)2021-07-19 08:35:14
新世紀智能(數學備考)(2020年9期)2021-01-04 00:25:14
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
數學物理學報(2020年2期)2020-06-02 11:29:24
光學精密工程(2016年6期)2016-11-07 09:07:19
