連作和輪作模式下花生土壤微生物群落不同微域分布特征*
2019-08-24 08:17:08姚小東李孝剛丁昌峰韓正敏王興祥
土壤學報 2019年4期
姚小東 李孝剛,? 丁昌峰 韓正敏 王興祥
(1 南方現代林業協同創新中心,南京林業大學生物與環境學院,南京 210037)(2 中國科學院土壤環境與污染修復重點實驗室(南京土壤研究所),南京 210008)
花生是我國重要的油料作物和經濟作物。2 0 1 6 年全國種植面積僅次于油菜,高達4 6 6 萬hm2,總產量1571萬t,在我國農業生產和農產品出口貿易中占有重要地位[1]。近年來,由于花生產區規模相對集中,常常多年成片、大規模種植,而隨著種植年限增加,連作障礙現象普遍發生,導致花生產量降低、品質變劣、生育狀況變差以及病蟲害嚴重等諸多生產問題,成為制約我國花生產業可持續發展的重要瓶頸[2]。
花生連作障礙的主要原因可能是,長期單一種植花生導致特定的根系分泌物刺激土壤中病原微生物大量生長并在根部定殖、進而破壞了根組織的正常分化和生理活動,使水分和養分運輸不足。同時,長期連作后,根系分泌物進入土壤也會對微生物種群進行特異性篩選,改變群落結構[3]。很多研究認為連作后土壤微生物多樣性下降,細菌總量下降而真菌顯著上升,連作土壤由“細菌性”轉為“真菌性”土壤[4,5]。諸如,尖孢鐮刀菌、黑曲霉等真菌、青枯勞爾氏菌均是引起花生土傳病害發生的重要病原菌[6,7],然而這些微生物在作物根際微域的分布、豐度與侵染力密切相關,對作物土傳病害的發生具有重要影響。但是,已有研究大多未考慮到根際土與根表土中微生物因受根系生理生化影響不同而產生的差異,多數研究中僅聚焦于非根際土壤或根區土,缺乏比較研究,尤其是連作作物根表微生物群落結構特征。此外,除典型病原菌外,連作栽培下根系分泌物對某些真菌類型也有化感誘導作用,如大豆根系分泌物能顯著促進粉紅粘帚菌的生長,連作太子參根際踝節菌大量繁殖[8, 9]。
近年來,高通量測序被廣泛應用于連作障礙研究。研究發現,花生長期連作后引起潛在病原菌如Fusarium sp.、Phoma sp.和Bionectria sp.相對豐度的增加[10]。然而,這些病原菌相對豐度的增加并不能準確表明其在土壤中的實際變化。此外,由于擴增片段短,大量菌群難以鑒定到種,且缺乏實體菌株的獲得,限制了進一步實驗研究。據此,本研究采用傳統分離培養技術和長片段基因克隆方法,通過比較輪作與連作花生根表、根際及非根際微生物數量、可培養微生物種群結構,以期從土壤可培養微生物組成的角度探討連作花生土傳病害高發的微生態機制,從而為解決花生連作障礙問題提供新的科學依據。
1 材料與方法
1.1 試驗地概況
試驗地位于江西省鷹潭市余江縣劉家站一分場典型的丘陵區紅壤旱地(116°55′E,28°12′N),花生種植普遍,兼有其他旱地作物類型如西瓜、紅薯等。土壤母質發育類型為第四紀紅色黏土,年均氣溫17.6 ℃,年均降水量1750 mm。
1.2 試驗設計
于2017年花生播種前期(3月)在區域內選擇2種典型的種植模式:1)連作,2012—2017年連續種植花生地塊;2)輪作,近年來采用各種旱作物輪換種植的地塊。連作花生地塊面積約為0.35hm2,花生栽培品種為贛花5號,每年4月上旬播種、7月下旬收獲,秋冬季拋荒。輪作地塊近5年分別采用花生、西瓜和紅薯進行倒茬種植,地塊面積約為0.5hm2,2016年種植季為西瓜,同樣秋冬季拋荒。
播種前采集2個地塊土壤樣品,帶回實驗室分析土壤理化性質,結果如表1所示,輪作和連作土壤有機質、pH、全氮、全磷、全鉀及速效養分等均無顯著性差異。于2017年4月上旬,2個地塊均開始播種花生,品種為贛花5號。花生播種株距10cm,行距50cm,每穴播種2粒。在播種前基施尿素150 kg·hm-2、氯化鉀 225 kg·hm-2、鈣鎂磷肥1125 kg·hm-2;其他管理措施如中耕培土、除草均按常規模式進行。
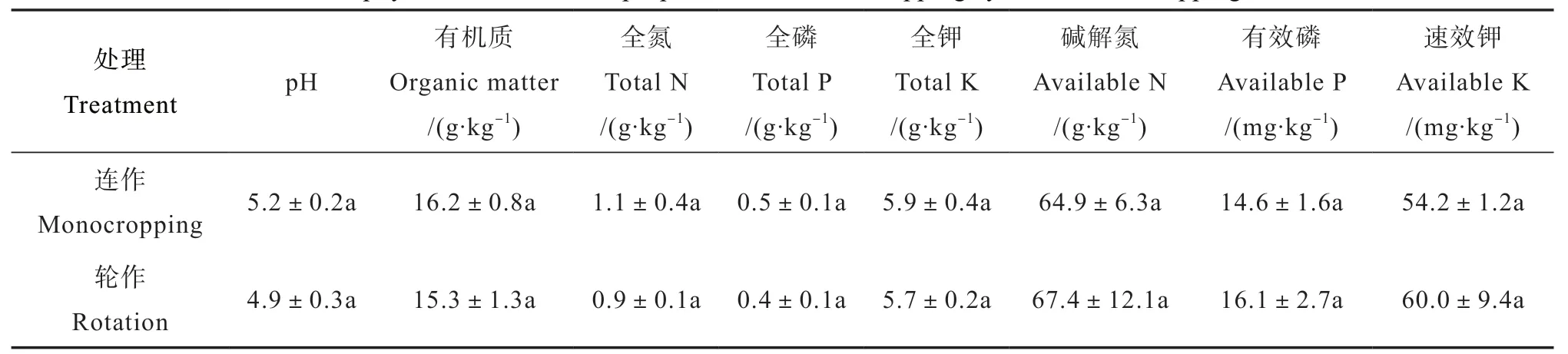
表1 連作、輪作花生土壤主要理化性質比較Table 1 Soil physical and chemical properties relative to cropping systems, monocropping or rotation
1.3 花生生長測定及病害分析
針對花生青枯病的分析策略如下:于2017年6月2日隨機5點取樣,每點選擇20株花生,共調查100株,分0~4級調查花生青枯病發生程度。0為沒有癥狀,1為花生25%以下的葉子萎焉,2為25%~50%葉子萎焉,3為75%以上的葉子萎焉但植株仍然存活,4為整個植株死亡。針對花生根腐病:于2017年7月6日,隨機5點調查,每點拔取20株花生,共100株,分0~4級調查花生根部病害程度。0為沒有癥狀,1為25%花生根部發病,2為25%~50%根部發病,3為50%~75%根部發病,4為整個根部發黑,植株死亡。
考種測產:花生收獲時(8月5日),每個地塊采集5個1 m×1 m的樣方,收獲花生莢果,折算單位面積的實際產量。同時,測定花生植株各生長指標和根瘤數量。
1.4 土壤樣品采樣
于花生花針期(6 月2 日)和結莢期(7 月6日),每塊樣地按5點取樣法采集不同微域土壤,每點隨機選取3株混合作為一個重復,每個地塊共采集5個重復樣品用于分析。花生輕輕拔出后,拍打整個根系使得疏松土塊全部脫落,利用無菌毛刷收集附在根系表面的土壤作為“根際”土壤。同時,在兩行植株中間的無根區域取0~20cm的耕層土壤作為相對的“非根際”土壤。將采過根際土的植株根系剪下稱重,置于裝有50 mL無菌水的三角瓶中,搖床上100 r·min-1振蕩30 min后取出根系,吸水紙吸盡根周圍的水分后再稱重,2次質量之差即為根表土質量[11];溶解于水中的土壤作為“根表土壤”用于后續分析。其他土壤樣品過2 mm篩混勻,去除根系、植物殘體以及其他雜質,4 ℃保存,3 d內完成土壤微生物區系測定。
1.5 土壤微生物數量測定
總真菌采用馬鈴薯葡萄糖瓊脂培養基(PDA),總細菌采用大豆酪蛋白瓊脂培養基(TSA)。分別采用Nash和Snyder培養基(NS)[12]、TSB培養基(TSB)、Masaago培養基(MAS)[13]、和假單胞菌CFC選擇培養基(杭州百思生物技術有限公司),對土壤典型微生物類型進行選擇培養。采用平板涂抹法測定各種微生物數量,換算為每克干土所含菌群數量表示。各種微生物具體采用的土壤稀釋梯度、培養條件見表2。

表2 用于培養土壤各類微生物的培養基、稀釋濃度及培養條件Table 2 Incubation condition, dilutions and media for different cultural microorganisms in soils
1.6 土壤微生物種類鑒定
由于分離、鑒定工作繁重,難以對所有樣品進行微生物種類鑒定,本研究主要分離、鑒定花針期花生根表中各種微生物的類型。計數后,在真菌性培養平板上隨機挑取典型單菌落于PDA平板上培養,經多次純化后轉入PDA斜面,于4℃冰箱保存。在細菌性培養平板上挑取單菌落,經過平板劃線于NA培養基上,37℃培養1~2d,經多次純化后轉入NA斜面,于4℃冰箱保存。純化的真菌菌株在PDA平板上活化,7d后挑取少量菌絲,液氮研磨,使用試劑盒Ezup柱式真菌基因組DNA抽提試劑盒(上海生工生物工程有限公司)提取DNA。純化的細菌菌株接種于NA液體培養基,振蕩培養24h,收集菌液,采用Ezup柱式細菌基因組DNA抽提試劑盒(上海生工生物工程有限公司)提取DNA。
細菌1 6 S r D N A 采用通用引物2 7 F(5′-A G A G T T T G AT C C T G G C T C A G-3′)和1492R(5′-TACCTTGTTACGACTT-3′)[14],真菌I T S 序列采用通用引物I T S 5(5′-GGAAGTAAAAGTCGTAACAAGG-3′)和ITS4(5′-TCCTCCGCTTATTGATATGC-3′)[15]進行擴增測序。4 0 μ L P C R 反應體系包含2×MASTER Mix(20 μL)、模板DNA、正反向引物(10μmol·L-1)各1 μL,ddH2O補足至40 μL。PCR擴增反應程序為,真菌:94℃起始變性5min,94℃變性30s,56℃退火30 s,72℃延伸45 s,32個循環;72℃延伸7 min后4℃保存備用。細菌:94℃起始變性5 min,94℃變性30 s,56℃退火30 s,72℃延伸90 s,32個循環,72℃延伸7 min后4℃保存備用。
將上述的PCR產物送至上海生工生物工程有限公司分析,測序結果提交GenBank數據庫,用Blast軟件在GenBank數據庫進行相似性比對分析。從PDA培養基隨機挑選21個克隆,NS培養基13個,MAS培養基9個,TSB培養基15個,TSA培養基41個,CFC培養基21個等用于序列分析;根據每個克隆的測序頻度計算對應物種分檢的相關豐度。
1.7 數據分析
采用Microsoft Excel和SPSS軟件進行數據整理和分析,數據處理結果用平均值及標準差表示(means±SD),采用Dunnett法分析處理間差異的顯著性(P<0.05)。
2 結 果
2.1 連作對花生生長及病害發生的影響
在成熟期,輪作種植下花生莢果鮮重和生物量分別為8300 kg·hm-2、14100kg·hm-2,是連作條件下的1.89倍和1.32倍;每株平均有效根瘤數為305,高于連作134.6%。相應的,與連作相比,輪作花生根腐病和青枯病病情指數分別降低58.7%和65.8%(圖1)。
2.2 連作和輪作種植模式下花生土壤微生物數量變化規律

圖1 連作、輪作地塊花生生長、產量和病害發生情況Fig. 1 Peanut growth,yield anddisease incidencerelative to cropping system, monocropping or rotation
經典可培養技術方法研究表明,與輪作相比,花針期和成熟期連作花生根際微生物明顯較高,其中根表真菌數量增加83%~134%,根表細菌數量增加108%(花針期);而對于“非根際”樣品,連作與輪作花生之間無顯著差異。尖孢鐮刀菌的分析結果表明,花針期連作和輪作花生根表其數量無顯著差異;而到結莢期,連作花生根表數量高于輪作27.2%(P<0.05)。木霉和疫霉則在連作地塊花生“非根際”的數量均顯著低于輪作花生;而在花生“根際”和“根表”微域內卻呈現出相反的趨勢,連作花生根表木霉和疫霉的數量分別較輪作花生增加390.4%~1344.1%和39.9%~76.1%。假單胞菌在“非根際”微域連作花生的數量顯著低于輪作;但在“根表”和“根際”微域連作花生顯著高于輪作,其中在“根表”區域表現的差異最為顯著,增幅達47.1%~227.7%(圖2)。
從不同微域花生土壤的變異來看,各類微生物均表現從“根表”到“根際”再到“非根際”逐漸下降趨勢,其中“根表”到“根際”下降趨勢劇烈,例如真菌數量下降64.6%~97.8%,細菌數量下降73.8%~99.7%。方差結果顯示各類微生物從“根表”到“根際”的變異程度均高于“根際”到“非根際”(表3)。
2.3 連作和輪作種植模式下花生根表可培養真菌群落變化
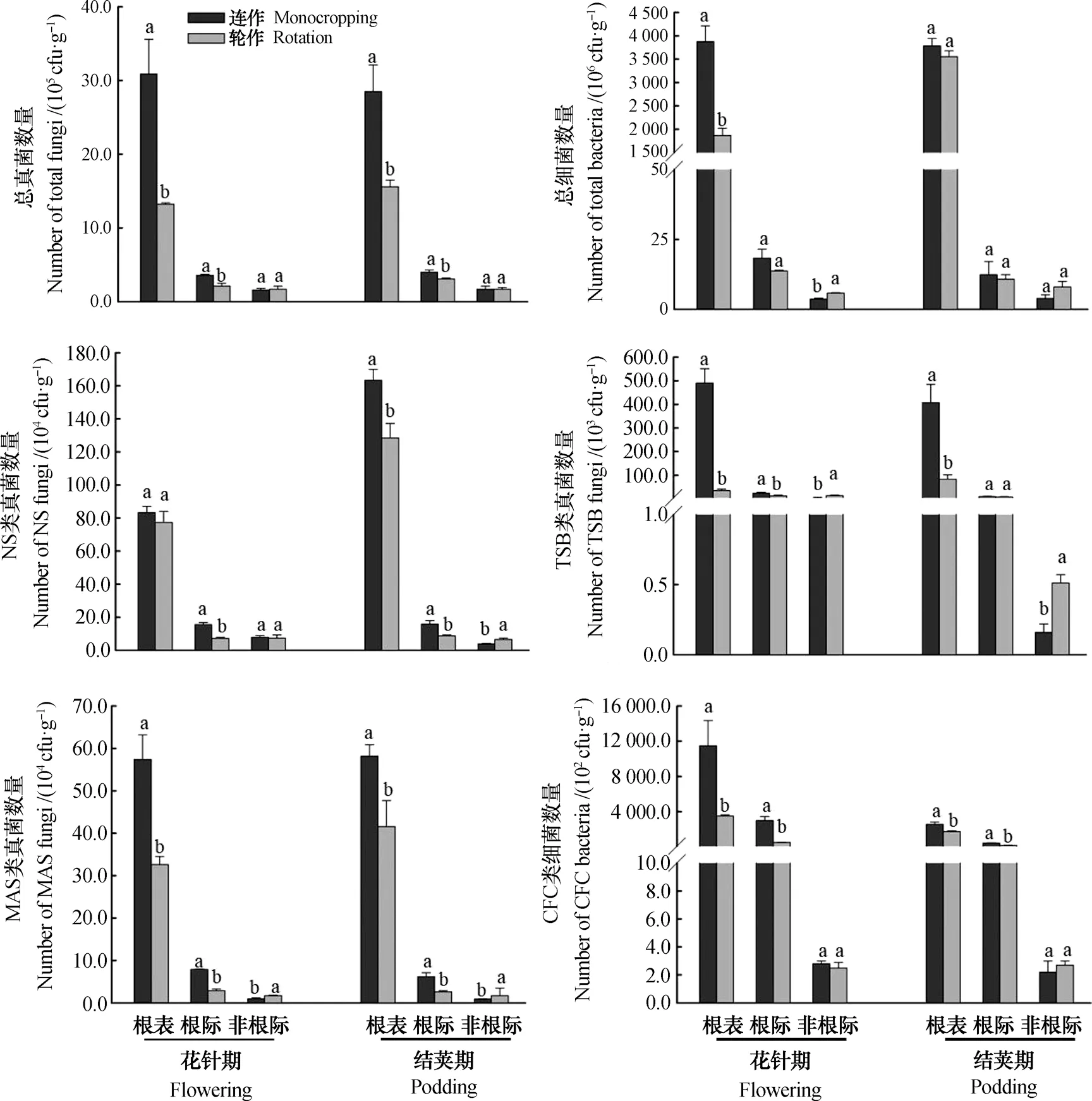
圖2 不同培養基下連作和輪作地塊花生根表、根際及非根際各類可培養微生物數量Fig. 2 Number of culturable microbes in peanut rhizoplane, rhizosphere and bulk relative to culture medium and cropping system
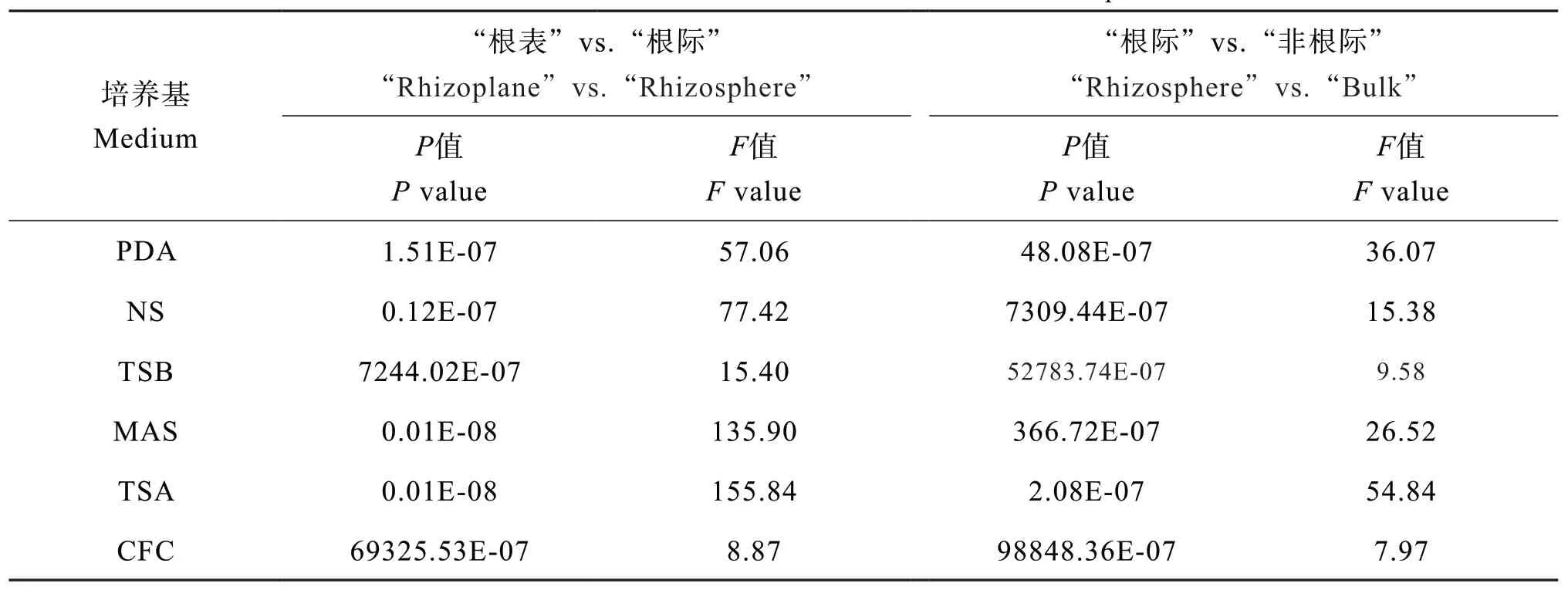
表3 不同微域分布的花生土壤可培養微生物數量的方差比較Table 3 ANOVA of culturable microbes relative to microzonesof peanut soil
由圖3可知,青霉屬(Penicillium sp.)在輪作與連作花生根表均能鑒定到,連作中以微紫青霉菌(P. janthinellum)和Penicillium s p.2 8 B R O-2 0 1 3 為主,輪作中以微紫青霉菌和Penicillium sp. 196F為主。輪作和連作花生根際均分離出曲霉屬(As pergillu s s p.),連作中包括黑曲霉(Aspergillus niger)、煙曲霉(A. fumigatus)和Aspergillus sp. J2;輪作中包括黑曲霉、煙曲霉、Aspergillus sp. J2和Aspergillus sp. BAB-5682,其中連作中黑曲霉豐度較高。粉紅粘帚菌(Clonostachys rosea)主要在連作花生根表鑒定到,且豐度高于輪作花生。在連作花生根表鑒定到踝節屬中Talaromyces pinophilus、T. verruculosus、T. aculeatus,毛殼菌(C h a e t o m i u m s p. AT T 2 1 0),尖孢鐮刀菌(F u s a r i u m o x y s p o r u m)和小球腔菌(Leptosphaeria spegazzinii),但這些真菌類型沒有在輪作花生根表分離到,其中連作花生根表踝節屬菌豐度較高。相反,莖潰瘍病菌(Diaporthe phaseolorum var. caulivora)、毛霉菌(M u c o r s p. F B C C 2 3 1 9)、白地霉(Geotrichum candidum)、木霉菌(Trichoderma gamsii)、卵形孢球托霉(Gongronella butleri)和Didymella molleriana僅在輪作花生根表中鑒定到,其中白地霉在輪作花生根表中豐度較高(圖3)。
用于分離鐮刀菌選擇性培養基(NS)中,除了鑒定到尖孢鐮刀菌外,還分離到青霉和白地霉等菌株。木霉選擇性培養基(TSB)僅分離到少量的木霉,多數屬于踝節菌和曲霉;而在疫霉選擇性培養基(MAS)中并未鑒定到疫霉,多數分離為粉紅粘帚菌和白地霉(圖3)。

圖3 不同培養基中連作和輪作花生根表可培養真菌(a)、細菌(b)鑒定及豐度Fig. 3 Identification of cultivable fungi (a) and bacteria (b) and their abundance in the peanut rhizoplanerelative to medium and cropping system
2.4 連作和輪作種植模式下花生根表可培養細菌群落變化
芽孢桿菌(B a c i l l u s s p.)、節細菌(Arthrobacter sp.)和假單胞菌(Pseudomonas sp.)是花生根表中常見類型,其中假單胞菌在輪作花生根表中豐度更高(圖3)。連作花生根表節細菌以煙草節桿菌(A. nicotianae)、Arthrobacter sp. 95-0188、分枝節桿菌(A. ramosus)和金黃節桿菌(A. aurescens)為主,而輪作中以嗜煙堿節桿菌(A. nicotinovorans)、Arthrobacter sp. 95-0188和分枝節桿菌為主。連作中假單胞菌只有黃褐假單胞菌(Pseudomonas fulva),輪作中包括Pseudomonas sp. BX2、Pseudomonas koreensis和惡臭假單胞菌(Pseudomonas putida)。
在連作和輪作花生根表中均能分離到生癌腸桿菌(Enterobacter cancerogenus),且豐度差異不明顯(圖3)。連作花生根際中分離到沙雷氏菌Serratia sp. MIE2、嗜線蟲沙雷氏菌(Serratia nematodiphila)、皮氏羅爾斯通氏菌(Ralstonia pickettii)、泛菌(Pantoea sp. 57917)和紅串紅球菌(Rhodococcus erythropolis),其中沙雷氏菌屬(Serratia sp.)豐度較高,但這些細菌在輪作花生根表沒有鑒定到。相反,泡囊短波單胞菌(Brevundimonas vesicularis)和溶淀粉類芽胞桿菌(Paenibacillus amylolyticus)均僅在輪作花生根表鑒定到(圖3)。用于分離假單胞菌的CFC培養基中,除了鑒定到假單胞菌外,還分離到一些沙雷氏菌、伯克氏菌(Burkholderia cenocepacia)和無色桿菌(Achromobacter xylosoxidans)等。
3 討 論
連作障礙是很多農作物普遍存在的問題,一般連作后均有可能影響土壤的理化和生物特性,進而影響作物生長及產量。本研究中連作和輪作土壤基本理化性質無顯著性差異。同樣,研究發現土壤高溫滅菌可有效控制連作土壤對作物生長的影響,有的還可明顯促進作物生長[16],說明土壤理化因子一般不是造成連作下作物生長受限的主要因素。本研究發現,與輪作花生相比,連作花生根部根瘤數量降低50%以上,進而導致連作花生自身固氮能力和氮素轉化的降低[17],這可能是引起連作地塊花生生長顯著低于輪作地塊的重要因素之一。然而,連作花生根部土傳病害發生顯著上升,這與王明珠和陳學南[18]針對紅壤區花生病害調查結果一致,說明花生連作后根系對病原菌的微生態抗性能力顯著下降。
根際效應是植物通過根系分泌物的代謝,致使植物根際微生物種類、數量和活性顯著高于非根際土壤的現象[19]。而目前關于揭示土壤微生物群落失衡與作物連作障礙發生關系的研究,沒有考慮到微生物數量和種群在土壤不同微域分布。從本研究結果來看,無論連作還是輪作花生,各種微生物數量均呈現從“根表”到“根際”再到“非根際”逐漸下降趨勢,尤其是從“根表”到“根際”層次下降幅度更加明顯。“根表”作為根土界面,其特定的生理代謝活性是影響根際微生物到“根表”定殖的關鍵因素。例如,Williams和Russo[20]研究發現根部Acidic Exopolysaccharide (EPS)基因的表達是影響根瘤菌附著大豆根表和形成生物膜的重要因素,同時發現根部分泌甘露聚糖可以介導土壤微生物大量附著在大豆根部表面。Kos等[21]也發現微生物的自凝集作用與其附著在根表面物質之間存在密切關系。總體而言,由于根域微生物變化與植物根系分泌物代謝特征關系密切,而根表是受根系分泌物代謝影響最為直接的部位,因此微生物數量在連作和輪作花生“根表”中差異巨大,說明“根表”附著的微生物種群結構與植物健康最為密切相關,未來有關連作障礙的微生物生態研究應著重于作物“根表”,因而針對連作障礙的微生物修復重點也應放在作物“根表”上。
通過對比連作和輪作地塊,發現連作花生在“根表”和“根際”層次上的真菌和細菌數量顯著較高,而在“非根際”層次無顯著差異。與本研究結果有所不同是,一些研究發現連作作物根際土壤呈現細菌數量下降、真菌數量上升的趨勢[22]。但也有研究表明連作后植物細菌數量高于輪作[23]。與“根際”相比,連作花生“根表”微生物受根系的影響及對根系的反作用明顯增強。這可能因為當根系受到病原微生物侵染發生病變時,根表組織潰爛、分泌物增多,導致根表上細菌、真菌數量與比例發生明顯變化。研究發現當病原菌侵染植物根部時不僅引起根際微生物群落組成的改變,還會導致根際生態代謝功能的變化[10,24,25]。Wu等[26]研究結果表明相比于健康三七植株,三七病株的根際微生物總量增加,并且根際微生物群落結構發生改變。而根瘤菌作為根域微生物的重要成員,根表微生物種群的變化可能影響植物結瘤。因此,“根表”微生物數量的急劇增多及組成改變可能是引起連作花生根瘤降低、根部病害上升的重要原因。但是,“根表”細菌和真菌豐度增多與連作花生根部生理代謝功能關系及“根表”根瘤菌種群變化,還需進一步研究。對于“非根際”土壤,由于距離根系相對較遠,故而受植物根系分泌物的影響很小,但其他栽培措施可能對其影響較大,這可能是引起連作和輪作地塊花生非根際土壤微生物數量無顯著差異的原因。對此,本研究從樣品采集及分析方法上對“根表”與“根際”土進行區別研究,有助于從更精確的微觀分析中發現作物連作障礙發生與微生物生態的關系。
進一步結合分子生物學鑒定來看,相對于輪作花生,連作花生根表中踝節菌(Talaromyces sp.)、黑曲霉(Aspergillus niger)和粉紅粘帚菌(Clonostachys rosea)種群豐度較高。其中,黑曲霉是花生土傳性病害—冠腐病的病原菌[7];也有研究報道踝節菌在連作太子參中大量繁殖,引起發病[9];粉紅粘帚菌是引起大豆根腐病潛在病原菌[9]。而在輪作花生根表中豐度較高的白地霉(Geotrichum candidum),有研究發現其對黃瓜枯萎病菌尖孢鐮刀菌的防治具有顯著成效[27]。值得注意的是,在鐮刀菌選擇培養基(NS)中,連作和輪作花生之間的“根表”尖孢鐮刀菌數量花針期并無明顯差異,但到了花生結莢期,連作花生“根表”尖孢鐮刀菌數量明顯高于輪作花生,這與收獲期連作花生根腐病發病率顯著高于輪作的結果一致,表明尖孢鐮刀菌可能是花生生長后期根腐病發生的重要病原菌。針對花生根表細菌,連作中沙雷氏菌屬豐度很高,而輪作花生根表的優勢種群轉變成假單胞菌,說明連作花生根表有益細菌數量大幅度下降與植株生長狀況密切相關[28]。此外,在鑒定到豐度相對較低的菌株中,連作花生根表中分離到的小球腔菌(Leptosphaeria sp.)、泛菌屬(Pantoea sp.)、紅球菌(Rhodococcus sp.)和青枯雷爾氏菌(Ralstonia sp.),有報道很多與植物發病有關[29]。相反,僅在輪作花生根表分離到的木霉(Trichoderma sp.)、類芽孢桿菌(Paenibacillus sp.)和短波單胞菌(Brevundimonas sp.)是很多病原菌的拮抗菌,尤其木霉已作為生防菌劑進行應用[30]。由此可見,輪作可能通過改善花生根表微生物結構,增加了有益/拮抗菌群的豐度,有效避免了病原菌在作物根表的定殖,從而降低了花生罹患土傳病害的風險。
目前,關于土壤微生物多樣性的研究方法已從傳統平板培養方法轉向為分子生物學方法,尤其是近年來高通量測序技術的普及,其一次性可以對高達數百萬的DNA分子序列進行測定,在微生物生態研究中具有數據通量高、微生物組成多樣性豐富等優勢[31]。同時,也面臨著海量數據分析難、測序短不能有效鑒定到“種”、數據庫中存在過多未鑒定物種以及實驗過程步驟繁瑣等問題。本研究采用傳統的微生物平板分離并結合分子生物技術,單個菌株可獲得更長的基因組片段,對各分離菌株做了更為準確的定位。并且,所分離到的各菌株可用于后續微生物生態功能研究,有助于進一步揭示連作作物根部病害與微生物生態的關系。
4 結 論
花生連作后植株生長受限、根瘤數減少、土傳性病害上升,進而導致花生產量降低。與“非根際”土壤相比,“根表”微生物受花生根系生理代謝活動影響最大,因而與連作障礙發生關系密切,是未來連作障礙微生態發生機制研究中重點關注部位。連作后花生“根表”細菌和真菌種群數量持續增多,但有益型微生物的鑒定頻度降低,而病原性微生物種群豐度較高,進而導致“根表”微生物生態系統遭到破壞,可能是引起連作下土傳性病害大幅上升關鍵原因。
致 謝 感謝中國科學院南京土壤研究所時玉博士在構建系統進化樹中的幫助!
