大鱗副泥鰍腐皮病病原菌及其拮抗菌的分離篩選
2019-09-18 07:23:40楊喬喬安賢惠韓迎亞隆小華李聯(lián)泰
水產(chǎn)科學(xué) 2019年5期
楊喬喬,安賢惠,韓迎亞,馬 臻,朱 明,2,隆小華,李聯(lián)泰
( 1.淮海工學(xué)院,江蘇 連云港 222005; 2.連云港龍?jiān)瓷锟萍加邢薰荆K 連云港 222005;3.南京農(nóng)業(yè)大學(xué),江蘇 南京 210095 )
泥鰍(Misgurnusanguillicaudatus),因其肉質(zhì)鮮美、營(yíng)養(yǎng)豐富而深受人們的喜愛,具有“水中人參”之美譽(yù)。連云港是全國(guó)最大的泥鰍養(yǎng)殖和出口基地,也是全國(guó)唯一的國(guó)家級(jí)出口泥鰍質(zhì)量安全示范區(qū)。每年出口泥鰍約1×104t,產(chǎn)值逾4×107美元。大鱗副泥鰍(Paramisgurnusdabryanus)因其體型較大,生長(zhǎng)快、抗逆性強(qiáng)、產(chǎn)量高等優(yōu)點(diǎn),成為近年新興的養(yǎng)殖品種。但隨著養(yǎng)殖面積的擴(kuò)大和養(yǎng)殖密度的增加,病害發(fā)生也愈加頻繁。其中泥鰍腐皮病是最為常見且危害較大的疾病之一。
泥鰍腐皮病傳染性強(qiáng)、潛伏期長(zhǎng)、發(fā)病率較高且危害大[1],其癥狀為鰍體局部或全身潰瘍,表面分布大面積血斑,有時(shí)也會(huì)出現(xiàn)爛身、爛鰭、食欲不振等情況。更嚴(yán)重的是,病鰍死后,其內(nèi)臟系統(tǒng)受損,大部分沉入水底,或埋于淤泥,因此不能及時(shí)處理。只有尸體被泡脹后才能浮出水面,導(dǎo)致死鰍腹腔內(nèi)大量病菌擴(kuò)散至整個(gè)池塘,從而造成更大的污染和病原傳播[2]。關(guān)于泥鰍腐皮病病原的研究,僅見蕭克宇等[3]報(bào)道的溫和氣單胞菌(Aeromonadssobria)。
對(duì)腐皮病的防控和治療,目前主要依賴傳統(tǒng)的化學(xué)藥物(包括抗生素等),其往往使病原菌產(chǎn)生耐藥性,影響治療效果;藥物殘留也會(huì)干擾水產(chǎn)動(dòng)物腸道益生菌的生長(zhǎng)和繁殖;殘留在水體中的消毒劑和抗生素破壞養(yǎng)殖水環(huán)境,導(dǎo)致水產(chǎn)品質(zhì)量降低[4]。有報(bào)道顯示,由于水產(chǎn)品藥物殘留導(dǎo)致出口受阻,給我國(guó)的水產(chǎn)品出口業(yè)造成巨大的經(jīng)濟(jì)損失[5]。利用拮抗菌來抑制病原菌的發(fā)生是一種環(huán)境友好型的病害防治方法。Kozasa[6]首次運(yùn)用1株自土壤中分離的東洋芽孢桿菌(Bacillustoyoi)處理鰻鱺(Anguillajaponica),降低了由愛德華菌(Edwardsiella)引起的鰻鱺死亡率;趙淑江等[7]從南麂島近海海洋沉積物中篩選出86株放線菌,其中有5株海洋放線菌對(duì)大黃魚(Pseudosciaenacrocea)病原性弧菌——副溶血弧菌(Vibrioparahaemolyticus)和哈維弧菌(V.harveyi)具有很強(qiáng)的抑制作用;Acurcio等[8]報(bào)道了3株腸球菌(Enterococcus),其具有很強(qiáng)的益生菌特性,對(duì)酸性條件和膽汁有很好的耐受性,并且能夠抑制李斯特氏菌(Listeia)、金黃色葡萄球菌(Staphyloccocusaureus)、大腸桿菌(Escherichiacoli)等致病菌;張歡歡等[9]自對(duì)蝦養(yǎng)殖池篩選出1株假交替單胞菌(Pseudoalteromonassp.),對(duì)鰻弧菌(V.anguillarum)、哈維弧菌及副溶血弧菌均有較強(qiáng)的抑制作用;何濤等[10]以草魚(Ctenopharyngodonidellus)病原菌維氏氣單胞菌(A.veronii)為指示菌,通過濾紙片抑菌圈法篩選出1株解淀粉芽孢桿菌(B.amyloliquefaciens),對(duì)該病原菌有較好的拮抗效果。本試驗(yàn)擬從泥鰍池中分離泥鰍腐皮病病原菌,以此為指示菌分離篩選拮抗菌,以期為泥鰍腐皮病的防治及相關(guān)生物制劑的開發(fā)奠定基礎(chǔ)。
1 材料與方法
1.1 材料
1.1.1 試驗(yàn)材料
患病泥鰍、水樣和泥樣,采自連云港市贛榆區(qū)某水產(chǎn)養(yǎng)殖公司。
1.1.2 水產(chǎn)病原菌
溫和氣單胞菌、維氏氣單胞菌、嗜水氣單胞菌(A.hydrophila)、哈維弧菌、鰻弧菌、遲緩愛德華菌(E.tarda)(海水菌株)等為本實(shí)驗(yàn)室保存菌株。
1.1.3 主要試劑
Tryptone、Yeast Extract(Oxoid LTD),革蘭氏染色液、細(xì)菌生化微量鑒定管(杭州微生物試劑有限公司),PCR所用試劑[寶生物工程(大連)有限公司],超氧化物歧化酶(SOD)測(cè)試盒、酸性磷酸酶(ACP)測(cè)試盒(南京建成生物工程研究所)。引物合成及測(cè)序由生工生物工程(上海)股份有限公司完成。
1.1.4 LB培養(yǎng)基
胰蛋白胨10 g/L,酵母提取物5 g/L,NaCl 10 g/L,瓊脂20 g/L(液體培養(yǎng)基中不加),pH 7.4~7.6,121 ℃滅菌30 min。
1.1.5 主要儀器
PCR儀(5332,德國(guó)Eppendorf公司),凝膠成像系統(tǒng)(OGGE,Bio-Rad公司),紫外可見分光光度計(jì)(T6新世紀(jì),北京普析通用儀器有限公司)。
1.2 方法
1.2.1 泥鰍腐皮病病原菌的分離
先用70%酒精棉球擦拭病灶部位表面,用無菌小刀切取病鰍約黃豆大小的病灶組織,用無菌剪刀剪碎,放入提前準(zhǔn)備好的3 mL LB液體培養(yǎng)基中,28 ℃恒溫?fù)u床培養(yǎng)2 h,然后用無菌生理鹽水10-1~10-6梯度稀釋,將10-4、10-5和10-6三個(gè)梯度分別涂布于LB培養(yǎng)基上,28 ℃培養(yǎng)24 h后觀察,將明顯可見差異的單菌落挑出,通過三區(qū)劃線接種到LB培養(yǎng)基上,并進(jìn)行連續(xù)多次三區(qū)劃線,直至得到純培養(yǎng)物[11]。
1.2.2 病原菌的篩選及回歸感染
將1.2.1分離的純培養(yǎng)物分別接種至LB液體培養(yǎng)基,28 ℃培養(yǎng)24 h后,采用分光光度法測(cè)定600 nm吸光度(OD600),根據(jù)平板菌落計(jì)數(shù)結(jié)果計(jì)算各菌株的培養(yǎng)液密度[11]。
為提高感染率,用手術(shù)刀輕輕劃傷健康泥鰍表皮組織(注意不要損傷肌肉組織),并迅速放入菌體密度為6×108cfu/mL的菌液中,浸泡15 min后,轉(zhuǎn)入未添加任何藥物的養(yǎng)殖水(3 kg/盒)中飼養(yǎng)[12]。觀察發(fā)病情況,篩選可感染腐皮病的病原菌株。
為進(jìn)一步確認(rèn)病原菌株,從試驗(yàn)中被感染的泥鰍病灶處,按照1.2.1方法再次分離病原菌并回歸感染健康泥鰍。每個(gè)菌株接種1組,每組設(shè)3個(gè)平行,每平行10尾泥鰍,以無菌LB培養(yǎng)基替代菌液為對(duì)照組。每個(gè)平行分別單獨(dú)計(jì)時(shí),保證每個(gè)平行的泥鰍感染時(shí)間一致。
自人工感染并發(fā)病的泥鰍病灶處,再次進(jìn)行細(xì)菌分離,按照1.2.1的方法進(jìn)行。通過菌落形態(tài)和顯微鏡觀察,鑒定人工感染后分離的菌株與人工感染使用的菌株是否相同。
1.2.3 腐皮病拮抗菌的分離
采集連云港市贛榆區(qū)某水產(chǎn)養(yǎng)殖公司泥鰍養(yǎng)殖池水樣和底泥,用無菌生理鹽水按不同梯度稀釋,然后涂布到LB培養(yǎng)基上,28 ℃培養(yǎng)24 h后,根據(jù)菌落顏色、形態(tài)、大小、邊緣整齊度及透明度等特征的不同,用接種環(huán)挑取單菌落進(jìn)行劃線分離,直至得到純培養(yǎng)物。分別挑取單菌落于LB液體培養(yǎng)基中,測(cè)定OD600,根據(jù)平板菌落計(jì)數(shù)結(jié)果計(jì)算各菌株的培養(yǎng)液密度,作為待測(cè)菌液。
1.2.4 拮抗菌的初篩和復(fù)篩
采用濾紙片抑菌圈法進(jìn)行初篩[13]。接種病原菌于LB液體培養(yǎng)基中,28 ℃培養(yǎng)16 h。取10 μL培養(yǎng)液加入培養(yǎng)皿中,倒入冷卻至約40 ℃的LB固體培養(yǎng)基(約20 mL),輕輕搖晃混勻。將直徑為6 mm的無菌濾紙片置于培養(yǎng)皿中,再將20 μL待測(cè)菌液滴加于濾紙片上,28 ℃培養(yǎng)24 h后觀察測(cè)定抑菌圈大小。將出現(xiàn)抑菌圈的菌株再次復(fù)篩,篩選出抑菌圈較大而且穩(wěn)定出現(xiàn)的菌株進(jìn)行后續(xù)試驗(yàn)(抑菌圈直徑用平均值±標(biāo)準(zhǔn)差表示)。
1.2.5 拮抗菌X8的傳代穩(wěn)定性
按1.2.4方法進(jìn)行操作。初代菌為分離純化保存的菌株。將初代菌重新三區(qū)劃線,挑取單菌落于LB液體培養(yǎng)基中,為二代菌。以此類推。以初代抑菌圈直徑為100%,接種9代,分別做抑菌圈試驗(yàn),觀察并測(cè)量抑菌圈直徑,試驗(yàn)重復(fù)3次。
1.2.6 病原菌和拮抗菌的菌株鑒定
1.2.6.1 形態(tài)特征觀察
群體形態(tài)觀察和個(gè)體形態(tài)觀察如革蘭氏染色、芽孢染色、莢膜染色、穿刺試驗(yàn)、細(xì)胞大小測(cè)定等均參照文獻(xiàn)[14]進(jìn)行。
1.2.6.2 生理生化試驗(yàn)
酪蛋白水解、明膠水解、淀粉水解和油脂水解等大分子水解試驗(yàn)參照文獻(xiàn)[14]進(jìn)行。微量生化管鑒定試驗(yàn)按杭州微生物試劑有限公司《細(xì)菌微量生化反應(yīng)管說明書》操作和觀察記載。
1.2.6.3 16S rRNA基因序列分析及系統(tǒng)發(fā)育樹構(gòu)建
本試驗(yàn)采用菌落PCR,從病原菌三區(qū)劃線平板上挑取單菌落,加入1 mL的無菌水,混勻后煮沸5 min,5000 r/min離心3 min,取上清液作為PCR反應(yīng)的模板。擴(kuò)增引物為細(xì)菌16S rRNA基因序列通用引物27F和1492R。反應(yīng)體系:50 U Taq酶0.5 μL,10×Buffer(含Mg2+)2.5 μL,10 mmol/L dNTP 2.5 μL,100 μmol/L引物各0.5 μL,模板1 μL,加ddH2O至25 μL。反應(yīng)程序:預(yù)變性95 ℃ 5 min;變性95 ℃ 30 s,復(fù)性55 ℃ 30 s,延伸72 ℃ 90 s,變性—復(fù)性—延伸30個(gè)循環(huán);再延伸72 ℃ 5 min。PCR產(chǎn)物送生工生物工程(上海)股份有限公司測(cè)序,在EZ.BioCloud網(wǎng)站查找模式菌株序列并下載,在Mega 6.06軟件中比對(duì)序列并構(gòu)建系統(tǒng)發(fā)育樹。
1.2.7 拮抗菌X8對(duì)不同溫度的適應(yīng)性
以0.5%的接種量接種至LB液體培養(yǎng)基中,分別置于20、25、30、35、40 ℃搖床中,180 r/min培養(yǎng)36 h。分別取20 μL發(fā)酵液,滴加于無菌濾紙片上,再將濾紙片置于混有病原菌的LB培養(yǎng)基上,28 ℃培養(yǎng)24 h,觀察測(cè)定抑菌圈大小。試驗(yàn)重復(fù)3次。
1.2.8 拮抗菌X8對(duì)不同鹽度的適應(yīng)性
以0.5%的接種量分別接種至鹽度為0、10、20、30、40、50的LB液體培養(yǎng)基中,28 ℃培養(yǎng)24 h。按照1.2.7的方法進(jìn)行操作。試驗(yàn)重復(fù)3次。
1.2.9 拮抗菌X8對(duì)泥鰍生理指標(biāo)的影響
1.2.9.1 體長(zhǎng)和體質(zhì)量增加量的測(cè)定
將泥鰍分為試驗(yàn)組和對(duì)照組兩組,每組設(shè)置3個(gè)平行,每個(gè)平行15尾泥鰍,平均體長(zhǎng)(13.2±0.3) cm,體質(zhì)量(10.1±0.2) g,置于直徑20 cm、高10 cm的飼養(yǎng)盒中,每盒保持1 L的水量。每隔2 d換水1次。水溫為(23±1) ℃。每日稱取泥鰍體質(zhì)量,按體質(zhì)量的2%進(jìn)行投食,早晚各1次。拮抗菌X8以0.5%的接種量接種至LB液體培養(yǎng)基中,30 ℃培養(yǎng)36 h,制成菌液,進(jìn)行平板菌落計(jì)數(shù),得到菌液密度。試驗(yàn)組加入1 mL菌液,使得水體中拮抗菌的密度達(dá)到105cfu/mL數(shù)量級(jí)。對(duì)照組加入1 mL無菌LB液體培養(yǎng)基。換水后,試驗(yàn)組和對(duì)照組重新添加菌液和無菌LB液體培養(yǎng)基。觀察期為28 d,觀察每組泥鰍的生長(zhǎng)狀況,測(cè)量各組總體長(zhǎng)和總體質(zhì)量(15尾合計(jì)),計(jì)算觀察期內(nèi),總體長(zhǎng)和總體質(zhì)量的增加值,分析試驗(yàn)組和對(duì)照組的差異情況。
1.2.9.2 超氧化物歧化酶和酸性磷酸酶活性的測(cè)定
28 d觀察期結(jié)束后,將各組泥鰍斷尾取血,離心后取血清,-20 ℃保存。超氧化物歧化酶和酸性磷酸酶活性的測(cè)定均按照測(cè)試盒說明書進(jìn)行。
采用SPSS 17.0軟件對(duì)數(shù)據(jù)進(jìn)行單因素方差分析,P<0.05為差異顯著,P<0.01為差異極顯著。
1.2.10 拮抗菌X8對(duì)不同水產(chǎn)病原菌的拮抗作用
活化本實(shí)驗(yàn)室保存的幾株水產(chǎn)致病性菌株:維氏氣單胞菌、溫和氣單胞菌、嗜水氣單胞菌、遲緩愛德華菌、哈維弧菌、鰻弧菌。采用濾紙片抑菌圈法(無菌LB液體培養(yǎng)基為對(duì)照),按照1.2.4進(jìn)行操作。觀察并測(cè)量抑菌圈大小。試驗(yàn)重復(fù)3次。
2 結(jié)果與分析
2.1 病原菌的分離
自患病泥鰍體表病灶處采樣,共分離出25個(gè)純培養(yǎng)物。編號(hào)分別是D1-1,D1-2-1,D1-2-2,…,D5-7,…,D7。
2.2 病原菌的篩選和回歸感染
2.2.1 病原菌的篩選
將上述分離到的25個(gè)純培養(yǎng)物,按方法1.2.2感染健康泥鰍,其中只有編號(hào)為D5-7的菌株可誘發(fā)泥鰍出現(xiàn)腐皮、爛身等典型的腐皮病癥狀,因此初步確定D5-7為病原菌株。
2.2.2 回歸感染
為進(jìn)一步證明菌株D5-7為泥鰍腐皮病的病原菌,再次自菌株D5-7引起發(fā)病的病灶部位取樣,按1.2.2的方法進(jìn)行分離和感染,同樣出現(xiàn)了與自然患病相同的腐皮病特征(圖1)。人工感染試驗(yàn)后,取發(fā)病泥鰍的病灶組織進(jìn)行分離,確實(shí)分離到與D5-7形態(tài)特性一致的菌株,表明菌株D5-7為泥鰍腐皮病的病原菌。
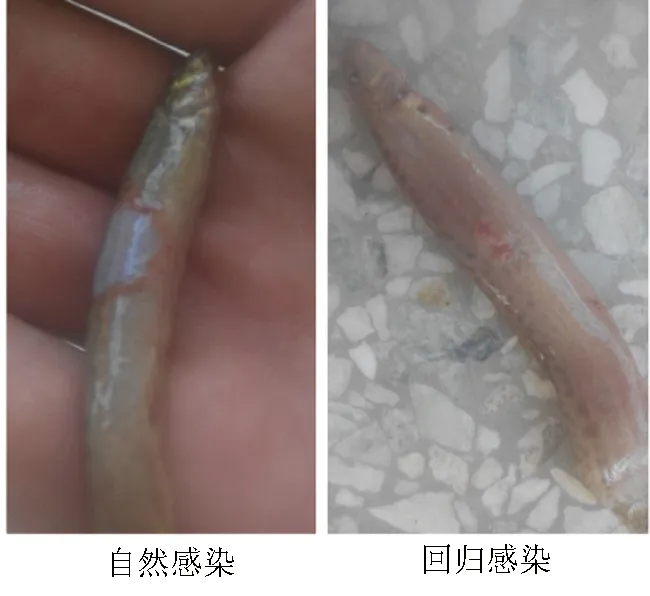
圖1 感染腐皮病的泥鰍
2.3 拮抗菌的分離
自泥鰍養(yǎng)殖池水和底泥采集的樣品經(jīng)過稀釋涂布、三區(qū)劃線,共分離出11株菌株,編號(hào)分別為L(zhǎng)11、L12、L13、L14、L15、L17、L18、L19、L20、H25、X8。
2.4 拮抗菌的篩選
2.4.1 拮抗菌的初篩和復(fù)篩
按1.2.4方法進(jìn)行操作,結(jié)果見圖2a,菌株L11、X8、L18、H25出現(xiàn)抑菌圈,直徑分別為(11.2±0.11) mm、(14.2±0.10) mm、(11.9±0.10) mm、(9.9±0.20) mm。將出現(xiàn)抑菌圈的菌株進(jìn)行復(fù)篩,篩選出抑菌圈較大且穩(wěn)定出現(xiàn)的菌株X8進(jìn)行后續(xù)試驗(yàn)(圖2b)。
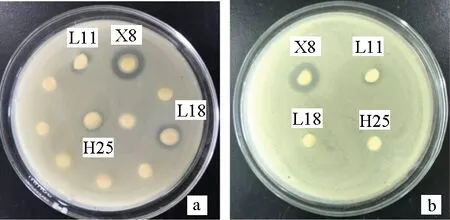
圖2 拮抗菌的初篩(a)和拮抗菌的復(fù)篩(b)
2.4.2 拮抗菌的傳代穩(wěn)定性
按1.2.4方法進(jìn)行操作。以初代抑菌圈直徑為100%,接種9代,抑菌圈直徑分別為初代的105%、109%、106%、96%、95%、94%、107%、111%、110%,證明X8的抑菌活性能夠穩(wěn)定傳代。
2.5 病原菌和拮抗菌的菌株鑒定
2.5.1 形態(tài)特征觀察
在LB平板上將病原菌D5-7三區(qū)劃線,28 ℃培養(yǎng)24 h后觀察。菌株D5-7菌落呈淡黃色,圓形,中央鼓起,邊緣整齊,光滑,半透明或接近不透明;革蘭氏染色后菌體呈紅色(圖3),可以判斷菌株D5-7為革蘭氏陰性細(xì)菌。顯微觀察其大小為(1.43±0.19) μm×(0.7±0.08) μm;無芽孢,無莢膜。穿刺試驗(yàn)結(jié)果表明,菌株D5-7無運(yùn)動(dòng)性。
在LB平板上將拮抗菌X8三區(qū)劃線,28 ℃培養(yǎng)24 h后觀察。菌株X8菌落呈淡黃色,多邊形,邊緣不整齊,表面干燥不平滑,不透明;革蘭氏染色呈紫色(圖4),可以判斷菌株X8為革蘭氏陽性細(xì)菌。顯微觀察其大小為(2.99±0.28) μm×(1.17±0.17) μm;有芽孢,無莢膜。穿刺試驗(yàn)結(jié)果表明菌株X8無運(yùn)動(dòng)性。
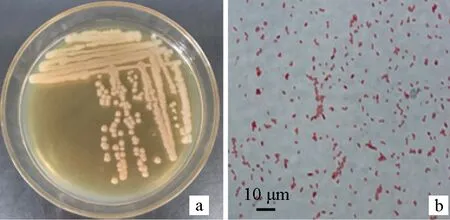
圖3 菌株D5-7的菌落形態(tài)(a)和革蘭氏染色結(jié)果(b)
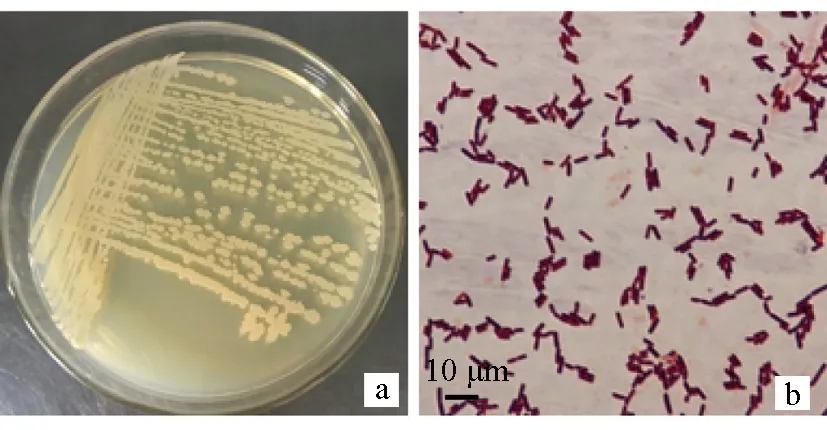
圖4 菌株X8的菌落形態(tài)(a)和革蘭氏染色結(jié)果(b)
2.5.2 生理生化試驗(yàn)
2.5.2.1 大分子水解試驗(yàn)
以短小芽孢桿菌(B.pumilus)E14[15]為對(duì)照進(jìn)行觀察,結(jié)果顯示,病原菌D5-7具有降解酪蛋白、明膠等蛋白質(zhì)的能力,但不能降解淀粉和油脂(圖5)。拮抗菌X8具有水解淀粉和明膠的能力,不能水解酪蛋白、油脂(圖6)。
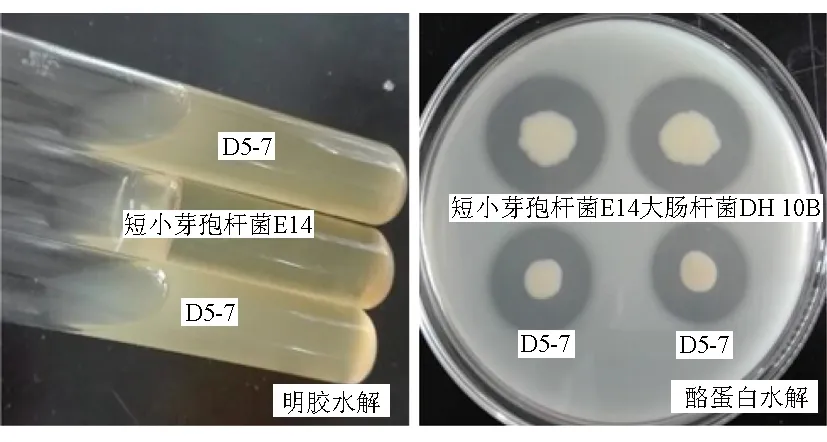
圖5 病原菌D5-7的蛋白水解試驗(yàn)
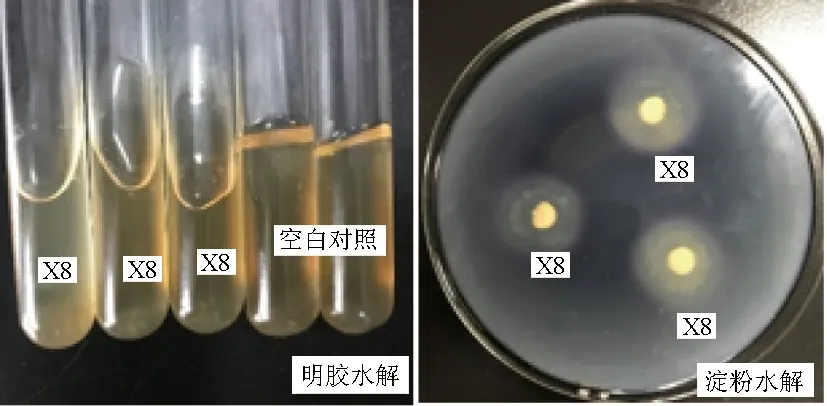
圖6 拮抗菌X8的淀粉和明膠水解試驗(yàn)
2.5.2.2 菌株D5-7和菌株X8的生理生化反應(yīng)
菌株D5-7和菌株X8的生理生化反應(yīng)見表1和表2。菌株D5-7能夠利用葡萄糖、蔗糖、甘露糖、阿拉伯糖等,不能利用丙二酸鹽、鼠李糖、阿拉伯醇、蜜二糖等。菌株X8能夠利用纖維二糖、葡萄糖、氰化鉀、丙二酸鹽等,不能利用果糖、乳糖、半乳糖、甘露醇等。
2.5.3 16S rRNA序列分析及系統(tǒng)發(fā)育樹構(gòu)建
擴(kuò)增菌株D5-7和X8的16S rRNA基因,16S rRNA基因片段大小約為1500 bp。將PCR產(chǎn)物送生工生物工程(上海)股份有限公司測(cè)序。
利用EZ.BioCloud網(wǎng)站查找模式菌株序列并下載,用Mega 6.06軟件采用臨接法(重復(fù)數(shù)為1000,步長(zhǎng)值取百分比)分別構(gòu)建菌株D5-7和X8的系統(tǒng)發(fā)育進(jìn)化樹(圖7、圖8)。
由圖7可知,病原菌D5-7與殺鮭氣單胞菌殺鮭亞種(Aeromonadssalmonicidassp.salmonicida) ATCC 33658遺傳距離最近,同源性為98%。綜合菌落和細(xì)胞形態(tài)觀察、生理生化試驗(yàn)及16S rRNA基因序列分析,將病原菌D5-7鑒定為氣單胞菌屬殺鮭氣單胞菌殺鮭亞種。
由圖8可知,拮抗菌X8與解淀粉芽孢桿菌(B.amyloliquefaciens)D15遺傳距離最近,同源性為98%。綜合菌落和細(xì)胞形態(tài)觀察、生理生化試驗(yàn)及16S rRNA基因序列分析,將拮抗菌X8鑒定為解淀粉芽孢桿菌。

表1 病原菌D5-7微量生化管反應(yīng)結(jié)果

表2 拮抗菌X8微量生化管反應(yīng)結(jié)果

圖7 基于16S rRNA基因序列構(gòu)建的菌株D5-7(LC312129)的系統(tǒng)發(fā)育樹
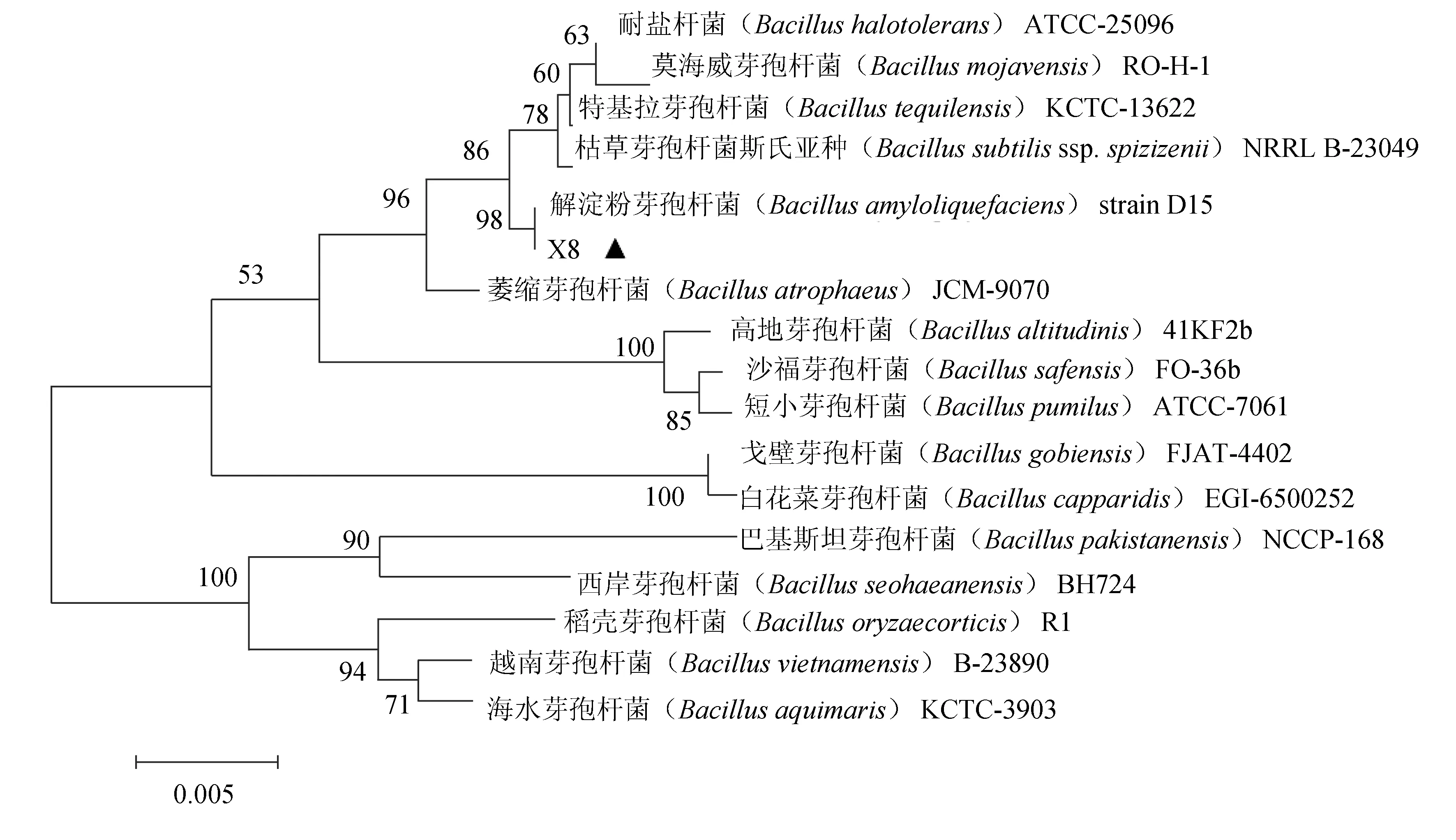
圖8 基于16S rRNA基因序列構(gòu)建的菌株X8(MH128163)的系統(tǒng)發(fā)育樹
2.6 解淀粉芽孢桿菌X8對(duì)不同溫度的適應(yīng)性
解淀粉芽孢桿菌X8在20~40 ℃之間具有抑菌活性(圖9)。在30 ℃時(shí),抑菌活性達(dá)到最高,抑菌圈直徑達(dá)(16.6±0.35) mm。
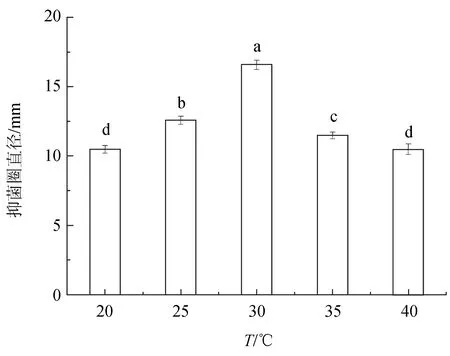
圖9 不同溫度對(duì)解淀粉芽孢桿菌X8拮抗活性的影響柱形圖上方不同小寫字母表示差異顯著(P<0.05).下同.
2.7 解淀粉芽孢桿菌X8對(duì)不同鹽度的適應(yīng)性
解淀粉芽孢桿菌X8在鹽度0~50之間具有抑菌活性(圖10)。在鹽度為0時(shí),抑菌活性最高,抑菌圈直徑達(dá)(21.7±0.18) mm。隨著鹽度的上升抑菌活性逐漸降低,在鹽度為50時(shí),抑菌活性最低,抑菌圈直徑僅為(10.1±0.37) mm。說明解淀粉芽孢桿菌X8在鹽度0~50內(nèi)具有較好的適應(yīng)性。
2.8 解淀粉芽孢桿菌X8對(duì)泥鰍生理指標(biāo)的影響
觀察期間,試驗(yàn)組與對(duì)照組泥鰍全部存活(表3)。體長(zhǎng)(cm)和體質(zhì)量(g)為試驗(yàn)前后增加的數(shù)值。試驗(yàn)組泥鰍的體長(zhǎng)和體質(zhì)量的增加值明顯大于對(duì)照組,差異達(dá)到極顯著水平(P<0.01),推測(cè)原因,可能是解淀粉芽孢桿菌X8抑制了水體中病原菌的生長(zhǎng),改善了生長(zhǎng)環(huán)境,促進(jìn)了泥鰍的健康生長(zhǎng)。
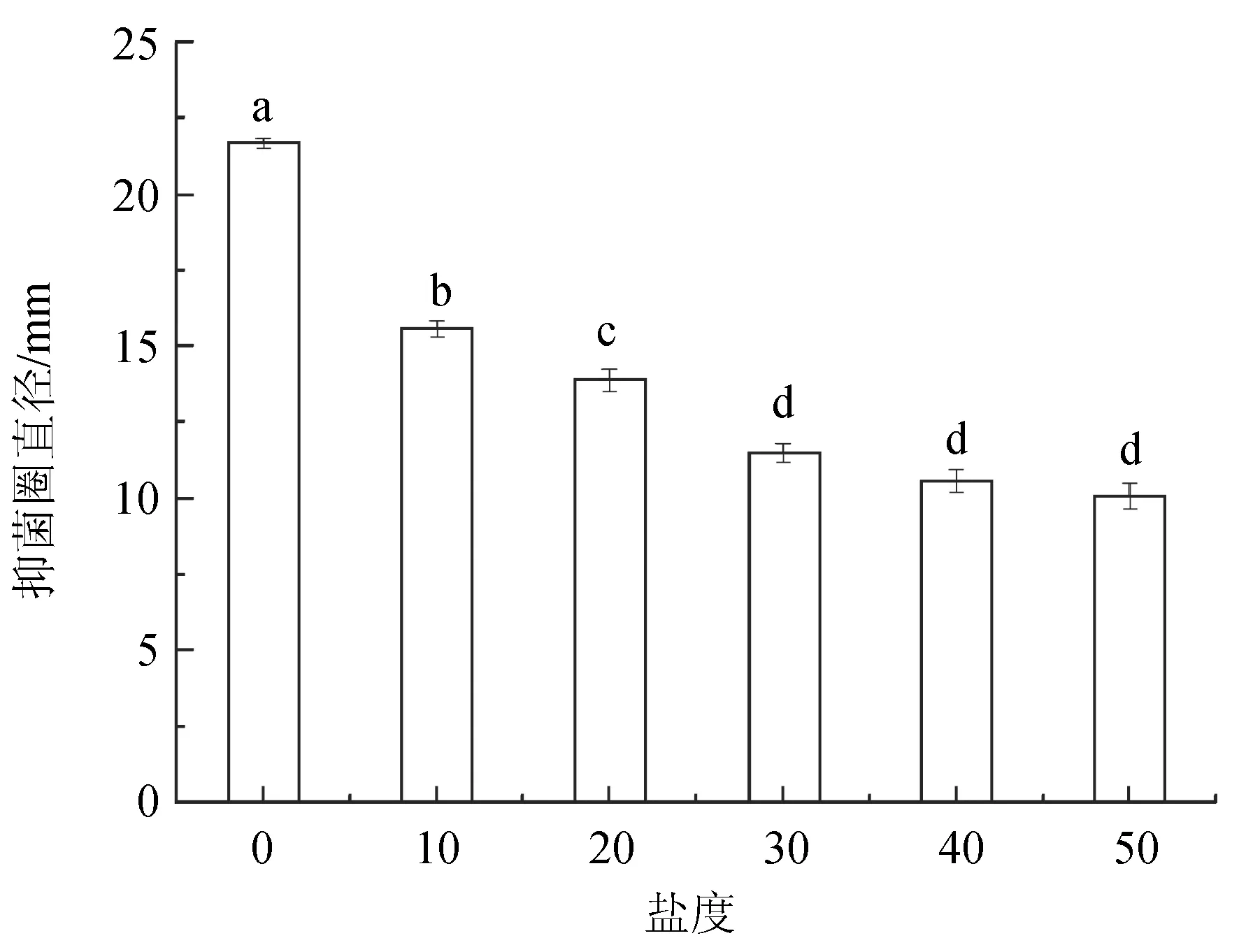
圖10 不同鹽度對(duì)解淀粉芽孢桿菌X8拮抗活性的影響
超氧化物歧化酶和酸性磷酸酶是魚類重要的兩個(gè)免疫指標(biāo)。試驗(yàn)組與對(duì)照組泥鰍血清中超氧化物歧化酶活性有所差別,但未達(dá)顯著水平(P>0.05)。試驗(yàn)組泥鰍血清中的酸性磷酸酶活性極顯著高于對(duì)照組(P<0.01)。酸性磷酸酶作為體內(nèi)巨噬細(xì)胞溶酶體的標(biāo)志酶,在體內(nèi)直接參與磷酸基團(tuán)的轉(zhuǎn)移和代謝,主要存在于各臟器、血細(xì)胞和骨髓中[16]。本試驗(yàn)中試驗(yàn)組酸性磷酸酶的增高,說明解淀粉芽孢桿菌X8可提高泥鰍的免疫力。
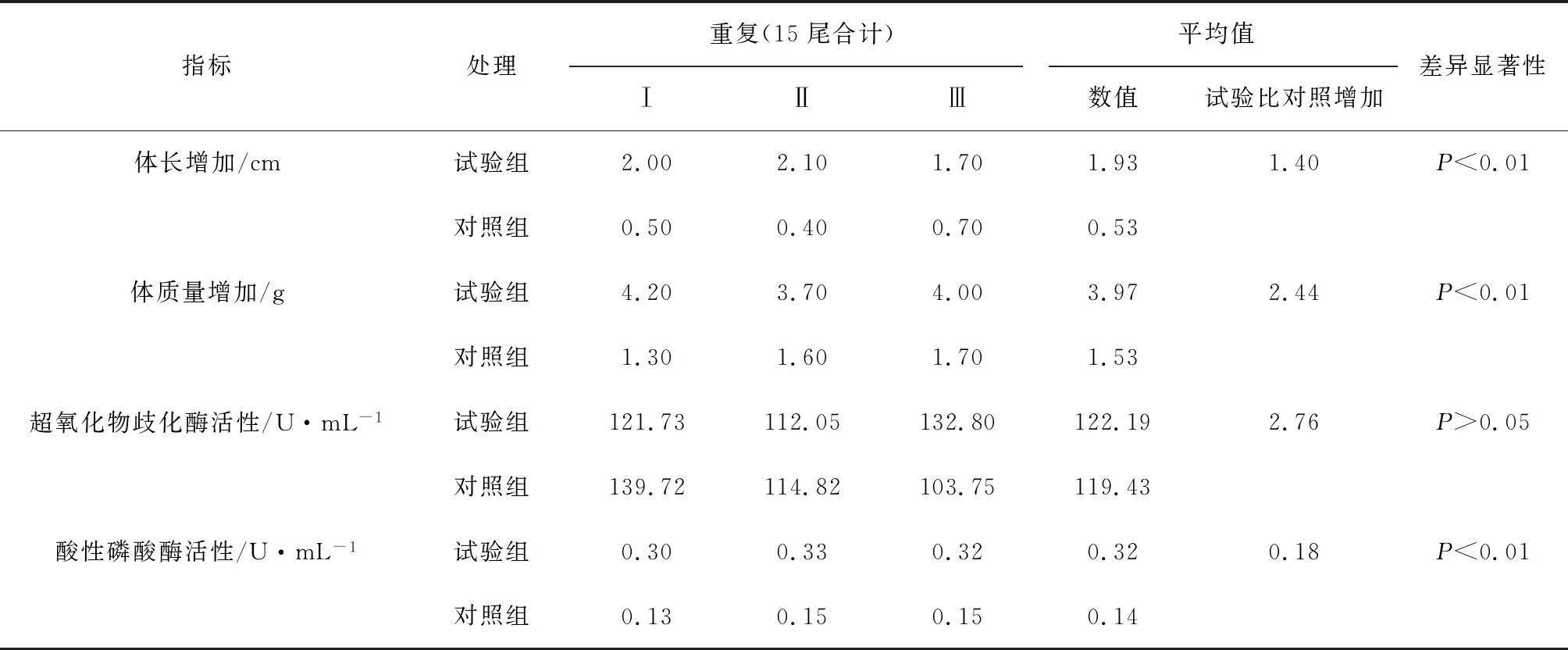
表3 解淀粉芽孢桿菌X8對(duì)泥鰍生理指標(biāo)的影響
2.9 解淀粉芽孢桿菌X8的拮抗譜
解淀粉芽孢桿菌X8對(duì)維氏氣單胞菌、溫和氣單胞菌、嗜水氣單胞菌、殺鮭氣單胞菌、遲緩愛德華菌、哈維弧菌、鰻弧菌有拮抗性(圖11),抑菌圈直徑分別達(dá)(14.7±0.10) mm、(16.8±0.15) mm、(11.3±0.20) mm、(15.9±0.15) mm、(8.2±0.05) mm、(11.8±0.10) mm、(8.2±0.08) mm。
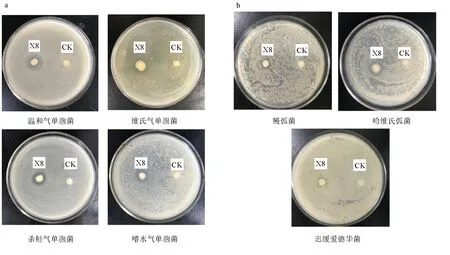
圖11 解淀粉芽孢桿菌X8對(duì)不同病原菌的抑制作用a.氣單胞菌,b.海水菌. CK為對(duì)照.
3 討 論
3.1 腐皮病病原菌的研究
誘發(fā)腐皮病的細(xì)菌性病原有嗜水氣單胞菌、溫和氣單胞菌、維氏氣單胞菌和點(diǎn)狀產(chǎn)氣單胞菌點(diǎn)狀亞種(A.punctatassp.punctata)等[3,17-18]。泥鰍腐皮病傳染性強(qiáng)、潛伏期長(zhǎng)、發(fā)病率較高且危害大,給養(yǎng)殖業(yè)帶來巨大的經(jīng)濟(jì)損失[19-20]。目前,關(guān)于泥鰍腐皮病病原菌,僅見蕭克宇等[3]報(bào)道的溫和氣單胞菌。本研究分離的殺鮭氣單胞菌也可誘發(fā)泥鰍腐皮病,目前尚未見報(bào)道。
殺鮭氣單胞菌屬氣單胞菌科、氣單胞菌屬,是一種引起鮭科魚類發(fā)生癤瘡病即腐皮病的典型細(xì)菌性病原[21]。1894年,Emmerich等[22]首次自患病溪鱒(Salvelinusfontinalis)中分離出該菌。其廣泛分布于淡水河流、湖泊、池塘等多種水環(huán)境,除美國(guó)南部地區(qū)以外,世界各地均有發(fā)現(xiàn)。近年來,被殺鮭氣單胞菌感染的水產(chǎn)動(dòng)物有半滑舌鰨(Cynoglossussemilaevis)[23]、異育銀鯽(Carassiusauratusgibelio)[24]、大西洋鮭(Salmosalar)[25]、細(xì)鱗魚(Brachymystaxlenok)[26]、大菱鲆(Scophthalmusmaximus)[27]、仿刺參(Apostichopusjaponicus)[28]、石鰈(Kareiusbicoloratus)[29]等,臨床癥狀表現(xiàn)為食欲減退,伴有出血性紅點(diǎn),嚴(yán)重者體表形成膿腫或潰瘍。本試驗(yàn)患病泥鰍除了上述病癥外,還出現(xiàn)鱗片脫落和臟器外露等癥狀。殺鮭氣單胞菌感染泥鰍的研究,擴(kuò)大了對(duì)殺鮭氣單胞菌可感染水生生物種類的認(rèn)知范圍。
3.2 腐皮病的誘發(fā)因素
誘發(fā)泥鰍腐皮病的主要因素有兩種:一是劃傷,二是溫度。當(dāng)泥鰍養(yǎng)殖池淤泥過多、池塘水質(zhì)不良、施用未經(jīng)充分發(fā)酵的糞肥,或在捕撈、運(yùn)輸過程中操作不慎引起鰍體受傷,導(dǎo)致泥鰍體表黏膜脫落或損傷,即可引起病原菌大量滋生。病原菌自泥鰍的受傷皮膚入侵,感染發(fā)病[18,30]。另外泥鰍腐皮病也為季節(jié)性發(fā)作病害,據(jù)初步觀察,江蘇連云港養(yǎng)殖泥鰍的發(fā)病時(shí)間通常為初春和秋末(即4—5月和9—10月),此時(shí)養(yǎng)殖池水溫通常為22~27 ℃,當(dāng)氣溫高于30 ℃時(shí),病情自然減弱直至消失。由此推測(cè),大鱗副泥鰍腐皮病的發(fā)生與溫度密切相關(guān),鰻鱺[31]的研究也有類似的情況。本試驗(yàn)篩選出的拮抗菌X8的適應(yīng)溫度為20~40 ℃,在生產(chǎn)上有一定的應(yīng)用價(jià)值。
3.3 拮抗菌X8為一種多功能菌株
近年來,以拮抗菌為代表的生物防治方法,具有環(huán)境友好、綠色安全等特點(diǎn),已受到越來越多的關(guān)注,不僅可以有效防治病害發(fā)生,還有助于改善水質(zhì),減少環(huán)境污染[32]。目前,國(guó)內(nèi)外應(yīng)用拮抗菌防治水產(chǎn)病害的報(bào)道較多,用來防治殺鮭氣單胞菌的研究也有報(bào)道[33-34],但使用拮抗菌防治泥鰍腐皮病的研究尚未見報(bào)道。筆者以殺鮭氣單胞菌D5-7為指示菌,自大鱗副泥鰍養(yǎng)殖池水樣中篩選并鑒定出1株拮抗菌——解淀粉芽孢桿菌X8,其抑菌圈直徑達(dá)(14.2±0.10) mm。該菌株的獲得,進(jìn)一步豐富了防治水產(chǎn)病害的拮抗菌資源。
有研究表明,解淀粉芽孢桿菌是重要的生防細(xì)菌,抑菌活性非常廣泛[35-36]。張雪等[37]研究表明,牛瘤胃源解淀粉芽孢桿菌對(duì)大腸桿菌、副溶血弧菌、藤黃八疊球菌(Sarcinalutea)和產(chǎn)氣桿菌(E.aerogenes)等細(xì)菌以及黑曲霉(Aspergillusniger)、黑根霉(Rhizopusnigricans)等霉菌均具有抑制作用。孫梅等[38]篩選出的解淀粉芽孢桿菌JSSW-LA,對(duì)多種水產(chǎn)病原菌如嗜水氣單胞菌、維氏氣單胞菌、溫和氣單胞菌、豚鼠氣單胞菌(A.caviae)均具有一定的抑制作用,其中對(duì)溫和氣單胞菌和豚鼠氣單胞菌的抑制效果顯著。本研究分離的拮抗菌解淀粉芽孢桿菌X8不僅對(duì)引起腐皮病的殺鮭氣單胞菌具有較強(qiáng)的抑制作用,對(duì)其他水產(chǎn)病原菌如維氏氣單胞菌、嗜水氣單胞菌、溫和氣單胞菌、遲緩愛德華菌、哈維弧菌、鰻弧菌等也有一定的拮抗作用。哈維弧菌、遲緩愛德華菌、鰻弧菌感染一般發(fā)生在鹽度為10~30的海水環(huán)境[39],解淀粉芽孢桿菌X8在鹽度40以下均對(duì)其有一定的抑菌活性。這與田良[40]的研究結(jié)果一致。另外,解淀粉芽孢桿菌被認(rèn)為是一種安全性較高的菌種,有望開發(fā)為水產(chǎn)養(yǎng)殖專用微生物制劑[41]。
解淀粉芽孢桿菌X8不僅對(duì)上述水產(chǎn)病害具有抑制作用,而且對(duì)泥鰍生長(zhǎng)具有良好的促進(jìn)作用。本試驗(yàn)結(jié)果表明,試驗(yàn)組比對(duì)照組體長(zhǎng)增加1.40 cm,體質(zhì)量增加2.44 g,酸性磷酸酶提高0.18 U/mL,三者均達(dá)到差異極顯著水平(P<0.01)。因此該菌株在生產(chǎn)養(yǎng)殖中具有廣闊的應(yīng)用前景。
