中國(guó)鱟幾丁質(zhì)酶的酶學(xué)性質(zhì)研究
2019-09-18 07:23:40林建城王麗英林娟娟
水產(chǎn)科學(xué) 2019年5期
關(guān)鍵詞:影響
林建城,王麗英,林娟娟
( 莆田學(xué)院 環(huán)境與生物工程學(xué)院,福建省新型污染物生態(tài)毒理效應(yīng)與控制重點(diǎn)實(shí)驗(yàn)室,福建 莆田 351100 )
Broadway等[1]根據(jù)幾丁質(zhì)酶作用機(jī)理不同,認(rèn)為其至少包含內(nèi)切幾丁質(zhì)酶(EC3.2.1.14)、外切幾丁質(zhì)酶和具外切酶性質(zhì)的N-乙酰-β-D-氨基葡萄糖苷酶(EC3.2.1.52,NAGase)等3種組分。微生物幾丁質(zhì)酶已受到廣泛關(guān)注[2],它能夠?qū)⒄婢?xì)胞壁、昆蟲體表和圍食膜的幾丁質(zhì)成分水解為幾丁寡糖和N-乙酰氨基葡萄糖,從而能有效抑制真菌的生長(zhǎng)和加速害蟲的死亡進(jìn)程[3];而在甲殼動(dòng)物的生長(zhǎng)中必須經(jīng)過(guò)一個(gè)周期性的動(dòng)態(tài)生理過(guò)程——蛻皮[4],近年研究發(fā)現(xiàn)甲殼動(dòng)物幾丁質(zhì)酶與其周期性蛻皮生理之間存在相關(guān)性[5]。Buchholz[6]研究表明,南極磷蝦(Euphausiasuperba)蛻皮與外殼幾丁質(zhì)酶活力變化相關(guān),在蛻皮前短時(shí)間內(nèi),內(nèi)切幾丁質(zhì)酶和N-乙酰-β-D-氨基葡萄糖苷酶大量合成,酶活力迅速提高,用于降解蝦幾丁質(zhì)表皮;樸順金等[7]在研究不同養(yǎng)殖期凡納濱對(duì)蝦(Litopenaeusvannamei)外殼膜N-乙酰-β-D-氨基葡萄糖苷酶的性質(zhì)中也發(fā)現(xiàn),在蝦幼體和生殖期時(shí),由于蛻殼頻繁,蝦外殼膜N-乙酰-β-D-氨基葡萄糖苷酶比活力會(huì)迅速升高。而Koga等[8]從家蠶(Bombyxmori)的表皮層中獲得N-乙酰-β-D-氨基葡萄糖苷酶,發(fā)現(xiàn)蠶表皮N-乙酰-β-D-氨基葡萄糖苷酶與其周期性的蛻皮生理、以及生活周期中蛹的形成密切相關(guān)。Arakane等[9]也發(fā)現(xiàn)昆蟲內(nèi)切幾丁質(zhì)酶可將其自身幾丁質(zhì)表皮降解為殼寡糖,再由外切幾丁質(zhì)酶繼續(xù)降解為N-乙酰氨基葡萄糖單體,并用以形成新的表皮。此后,王佩等[10]研究表明,日本沼蝦幾丁質(zhì)酶基因-3的表達(dá)水平以蛻皮前期最為顯著,從而推測(cè)與幾丁質(zhì)酶參與蝦舊殼的降解和新殼的形成有關(guān),進(jìn)一步證明了幾丁質(zhì)酶在甲殼動(dòng)物蛻皮生理中起重要作用。
中國(guó)鱟(Tachypleustridentatus)屬節(jié)肢動(dòng)物門、肢口綱、劍尾目,由于海域環(huán)境破壞和人類捕殺等原因,已成為瀕危物種,是國(guó)家二級(jí)保護(hù)動(dòng)物。隨著沿海地區(qū)工業(yè)化的不斷推進(jìn),鱟賴以生存的海域環(huán)境受到嚴(yán)重破壞,擾亂了鱟的正常生理活動(dòng)與物質(zhì)代謝。有研究表明,溫度和鹽度都會(huì)影響中國(guó)鱟胚胎發(fā)育的速度和孵化率,其胚胎發(fā)育最適溫度為28 ℃,最適鹽度為20[11];而Zn、Pb、Cu和Cd 4種重金屬離子對(duì)中國(guó)鱟的胚胎發(fā)育產(chǎn)生的影響不同,當(dāng)4種重金屬離子濃度超過(guò)漁業(yè)水域水質(zhì)標(biāo)準(zhǔn)濃度8倍時(shí),Cd2+和Cu2+對(duì)鱟的胚胎發(fā)育無(wú)明顯影響,但Zn2+和Pb2+會(huì)使鱟胚胎發(fā)育中卵徑變小,發(fā)育速度下降[12],說(shuō)明海洋環(huán)境因子的變化直接影響中國(guó)鱟的胚胎發(fā)育和幼體生長(zhǎng)。此外,有研究表明,溫度、酸堿度、金屬離子和有機(jī)溶劑等環(huán)境因素對(duì)節(jié)肢動(dòng)物幾丁質(zhì)酶會(huì)產(chǎn)生重要影響,并推測(cè)會(huì)影響節(jié)肢動(dòng)物的蛻皮生理[13-15]。而在中國(guó)鱟的胚胎發(fā)育到性成熟期間,至少需經(jīng)19次蛻皮,鱟蛻皮生理的變化直接影響鱟的正常生長(zhǎng)發(fā)育。然而,中國(guó)鱟幾丁質(zhì)酶的生理特性及環(huán)境因子對(duì)鱟幾丁質(zhì)酶的影響,目前尚未見相關(guān)研究。之前研究已自中國(guó)鱟內(nèi)臟中分離純化出N-乙酰-β-D-氨基葡萄糖苷酶[16],現(xiàn)繼續(xù)自中國(guó)鱟內(nèi)臟中分離提取內(nèi)切型幾丁質(zhì)酶,探討其酶學(xué)性質(zhì),并研究環(huán)境因子對(duì)中國(guó)鱟幾丁質(zhì)酶的影響,這對(duì)進(jìn)一步揭示鱟幾丁質(zhì)酶與蛻皮生理之間的相關(guān)性具有重要的作用,并為能更好地保護(hù)鱟資源,合理開發(fā)利用鱟資源提供科學(xué)依據(jù)。
1 材料與方法
1.1 材料
中國(guó)鱟購(gòu)自莆田市城南市場(chǎng)。酶催化的底物對(duì)硝基苯-N-乙酰-β-D-氨基葡萄糖苷,由上海醫(yī)藥工業(yè)研究院生化室合成;3,5-二硝基水楊酸為Fluka產(chǎn)品,標(biāo)準(zhǔn)物N-乙酰-D-氨基葡萄糖為Sigma產(chǎn)品,幾丁質(zhì)購(gòu)自中國(guó)醫(yī)藥集團(tuán)上海化學(xué)試劑公司,其余化學(xué)試劑均為國(guó)產(chǎn)分析純。
1.2 方法
1.2.1 中國(guó)鱟幾丁質(zhì)酶制劑的制備
取新鮮中國(guó)鱟內(nèi)臟,以1∶3(m/V)浸泡在預(yù)冷的0.01 mol/L Tris-HCl(pH 5.4)緩沖液中,在高速組織搗碎機(jī)中破碎1 min,4 ℃抽提4 h以上,勻漿液于4 ℃,12 000 r/min,離心30 min,取上清液。上清液依次采用35%、75%飽和度的(NH4)2SO4分級(jí)分離,沉淀物經(jīng)雙蒸餾水透析后于4 ℃、14 000 r/min 離心30 min,上清液為粗酶制劑。
1.2.2 膠體幾丁質(zhì)的制備
將條狀幾丁質(zhì)(甲殼素)在植物粉碎機(jī)下磨成細(xì)粉,10 g細(xì)粉幾丁質(zhì)加入100 mL濃鹽酸,4 ℃下間隔攪拌24 h,倒入500 mL預(yù)冷蒸餾水中,放置48 h,4000 r/min下離心,沉淀用蒸餾水洗至中性,用的50 mmol/L NaAc-HAc緩沖液(pH 5.4)配成5.0 mg/mL的膠體幾丁質(zhì)。
1.2.3 中國(guó)鱟幾丁質(zhì)酶活力的測(cè)定方法
中國(guó)鱟幾丁質(zhì)酶活力測(cè)定方法參照文獻(xiàn)[13],在0.5 mL的酶活力測(cè)定體系中含有50 mmol/L NaAc-HAc緩沖液(pH 5.4)和5.0 mg/mL膠體幾丁質(zhì),55 ℃下預(yù)熱10 min,后加入50 μL的酶液,在55 ℃下反應(yīng)30 min;以0.5 mL 0.5 mol/L的NaOH終止反應(yīng),離心,去沉淀,取上清液0.5 mL,加入0.5 mL 3,5-二硝基水楊酸,煮沸5 min,靜置至室溫后,測(cè)定其光吸收值(A520nm),以未加入酶制劑的為對(duì)照。以產(chǎn)物N-乙酰-D-氨基葡萄糖為標(biāo)準(zhǔn)物,制定標(biāo)準(zhǔn)曲線。以吸光值(y)為縱坐標(biāo),N-乙酰-D-氨基葡萄糖含量(x)為橫坐標(biāo),繪制標(biāo)準(zhǔn)曲線,得直線回歸方程:y=0.619x-0.151。
1個(gè)酶活力單位(U)定義為:每小時(shí)水解膠體幾丁質(zhì)產(chǎn)生1 μmol N-乙酰-D-氨基葡萄糖所需的酶量。
1.2.4 中國(guó)鱟幾丁質(zhì)酶最適pH和酸堿穩(wěn)定性的測(cè)定
在酶活力測(cè)定體系中,測(cè)定不同pH(3.6~9.0)下中國(guó)鱟幾丁質(zhì)酶活力,其中pH為3.6~5.8時(shí)選擇HAC-NaAC緩沖液,pH為5.8~7.6時(shí)采用Na2HPO4-NaH2PO4緩沖液,pH為7.6~9.0時(shí)使用Tris-HCl緩沖液,3種緩沖液濃度均為50 mmol/L,酶活力最大的試驗(yàn)組其pH值確定為酶的最適pH,其酶活力以100%表示,其余各試驗(yàn)組酶活力則以相對(duì)酶活力表示。
將酶液分別與各種不同pH(3.6~9.0)的緩沖液混合,4 ℃下各預(yù)處理1 h,然后取50 μL處理過(guò)的酶液,在酶活力測(cè)定體系中測(cè)定酶經(jīng)處理后的剩余酶活力,剩余酶活力最大的試驗(yàn)組其酶活力以100%表示,其余試驗(yàn)組的酶活力以相對(duì)酶活力表示,以分析幾丁質(zhì)酶的酸堿穩(wěn)定性。
相對(duì)酶活力/%=(a/amax)×100%
式中,a為各試驗(yàn)組(剩余)酶活力,amax為各試驗(yàn)組中最大(剩余)酶活力。
1.2.5 中國(guó)鱟幾丁質(zhì)酶最適溫度和溫度穩(wěn)定性的測(cè)定
在酶活力測(cè)定體系中,檢測(cè)不同溫度(20~90 ℃)下中國(guó)鱟幾丁質(zhì)酶活力,酶活力最大的試驗(yàn)組其溫度確定為酶的最適溫度,其酶活力以100%表示,其余試驗(yàn)組的酶活力以相對(duì)酶活力表示。
將幾丁質(zhì)酶液置于不同溫度(20~90 ℃)下預(yù)處理1 h,然后取50 μL經(jīng)熱處理過(guò)的酶,在酶活力測(cè)定體系中測(cè)定酶經(jīng)熱處理后的剩余酶活力,剩余酶活力最大的試驗(yàn)組其酶活力以100%表示,其余試驗(yàn)組的剩余酶活力以相對(duì)酶活力表示,以分析幾丁質(zhì)酶的溫度穩(wěn)定性。
1.2.6 中國(guó)鱟幾丁質(zhì)酶催化幾丁質(zhì)酶解的動(dòng)力學(xué)參數(shù)測(cè)定
在幾丁質(zhì)酶活力的測(cè)定體系中,改變底物膠體幾丁質(zhì)的質(zhì)量濃度,測(cè)定不同底物質(zhì)量濃度[S](2.0~8.0 mg/mL)下幾丁質(zhì)酶活力,采用Lineweave-Burk雙倒數(shù)作圖法,以反應(yīng)初速度倒數(shù)(1/v)對(duì)底物質(zhì)量濃度倒數(shù)(1/[S])作圖,求出中國(guó)鱟幾丁質(zhì)酶的米氏常數(shù)(Km)和最大反應(yīng)速度(Vmax)。
1.2.7 金屬離子對(duì)中國(guó)鱟幾丁質(zhì)酶活力影響的測(cè)定
在上述幾丁質(zhì)酶活力的測(cè)定體系中,分別加入0~20.0 mmol/L的Li2SO4、Na2SO4、NaCl、NaNO3和KNO3;0~5.0 mmol/L的MgSO4、CaCl2和BaCl2;0~20.0 mmol/L的FeSO4、CoCl2、NiCl2、CuSO4、ZnSO4、MnCl2、AlCl3、FeCl3、HgCl2、Pb(NO3)2和Cd(NO3)2;測(cè)定加入各金屬離子后的中國(guó)鱟幾丁質(zhì)酶活力。以未添加金屬離子的對(duì)照組酶活力為100%,計(jì)算各試驗(yàn)組的相對(duì)酶活力。
相對(duì)酶活力/%=(a/amax)×100%
式中,a為各試驗(yàn)組酶活力,amax為對(duì)照組酶活力。
1.2.8 有機(jī)溶劑對(duì)中國(guó)鱟幾丁質(zhì)酶活力影響的測(cè)定
在幾丁質(zhì)酶測(cè)活體系中,分別加入0~50.0%體積分?jǐn)?shù)的甲醇、乙醇、正丙醇、異丙醇、丙酮、二氧六環(huán)和二甲亞砜等7種有機(jī)溶劑,以未添加有機(jī)溶劑的對(duì)照組酶活力為100%,計(jì)算各試驗(yàn)組的相對(duì)酶活力。
2 結(jié)果與分析
2.1 pH對(duì)中國(guó)鱟幾丁質(zhì)酶活力的影響
由圖1可見,pH對(duì)中國(guó)鱟幾丁質(zhì)酶活力的影響呈鐘罩型曲線變化,幾丁質(zhì)酶最適pH為5.4,隨pH逐漸增大,幾丁質(zhì)酶活力也不斷下降。

圖1 中國(guó)鱟幾丁質(zhì)酶的最適pH
由圖2可見,幾丁質(zhì)酶在pH 4.6~6.0的緩沖液中預(yù)處理1 h后,相對(duì)酶活力還保留在80.7%以上,說(shuō)明幾丁質(zhì)酶在pH 4.6~6.0內(nèi)相對(duì)穩(wěn)定,經(jīng)pH 8.0緩沖液處理1 h后幾丁質(zhì)酶相對(duì)活力僅有11.5%;而在pH≤4.4的緩沖液中處理后幾丁質(zhì)酶活力也呈下降趨勢(shì),說(shuō)明該幾丁質(zhì)酶對(duì)環(huán)境中酸堿度的變化敏感。
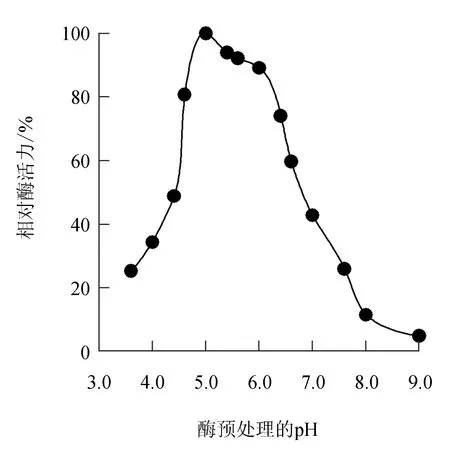
圖2 中國(guó)鱟幾丁質(zhì)酶的酸堿穩(wěn)定性
2.2 溫度對(duì)中國(guó)鱟幾丁質(zhì)酶活力的影響
由圖3可見,中國(guó)鱟內(nèi)臟幾丁質(zhì)酶的最適溫度為55 ℃,之后隨溫度升高酶活力不斷下降,溫度達(dá)80 ℃時(shí),酶活力剩余36.6%。
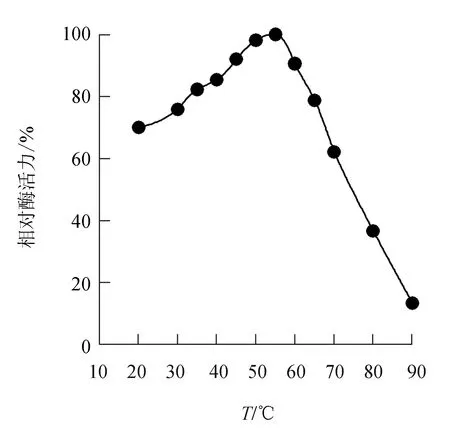
圖3 中國(guó)鱟幾丁質(zhì)酶的最適溫度
由圖4可見,幾丁質(zhì)酶在20~60 ℃預(yù)處理1 h后相對(duì)酶活力保持在88%以上,酶在65 ℃處理1 h相對(duì)酶活力還保留有79.8%,在90 ℃下處理1 h相對(duì)酶活力還剩34.3%,表明中國(guó)鱟內(nèi)臟幾丁質(zhì)酶具較好的熱穩(wěn)定性。
2.3 中國(guó)鱟幾丁質(zhì)酶米氏常數(shù)和最大反應(yīng)速度的測(cè)定
Lineweaver-Burk雙倒數(shù)作圖(圖5)顯示,反應(yīng)初速度倒數(shù)(y)與底物濃度倒數(shù)(x)的回歸方程為y=1.494x+0.687,求得催化膠體幾丁質(zhì)底物水解的中國(guó)鱟幾丁質(zhì)酶米氏常數(shù)為2.175 mg/mL,最大反應(yīng)速度為1.456 μmol/(mL·min)。

圖4 中國(guó)鱟幾丁質(zhì)酶的溫度穩(wěn)定性
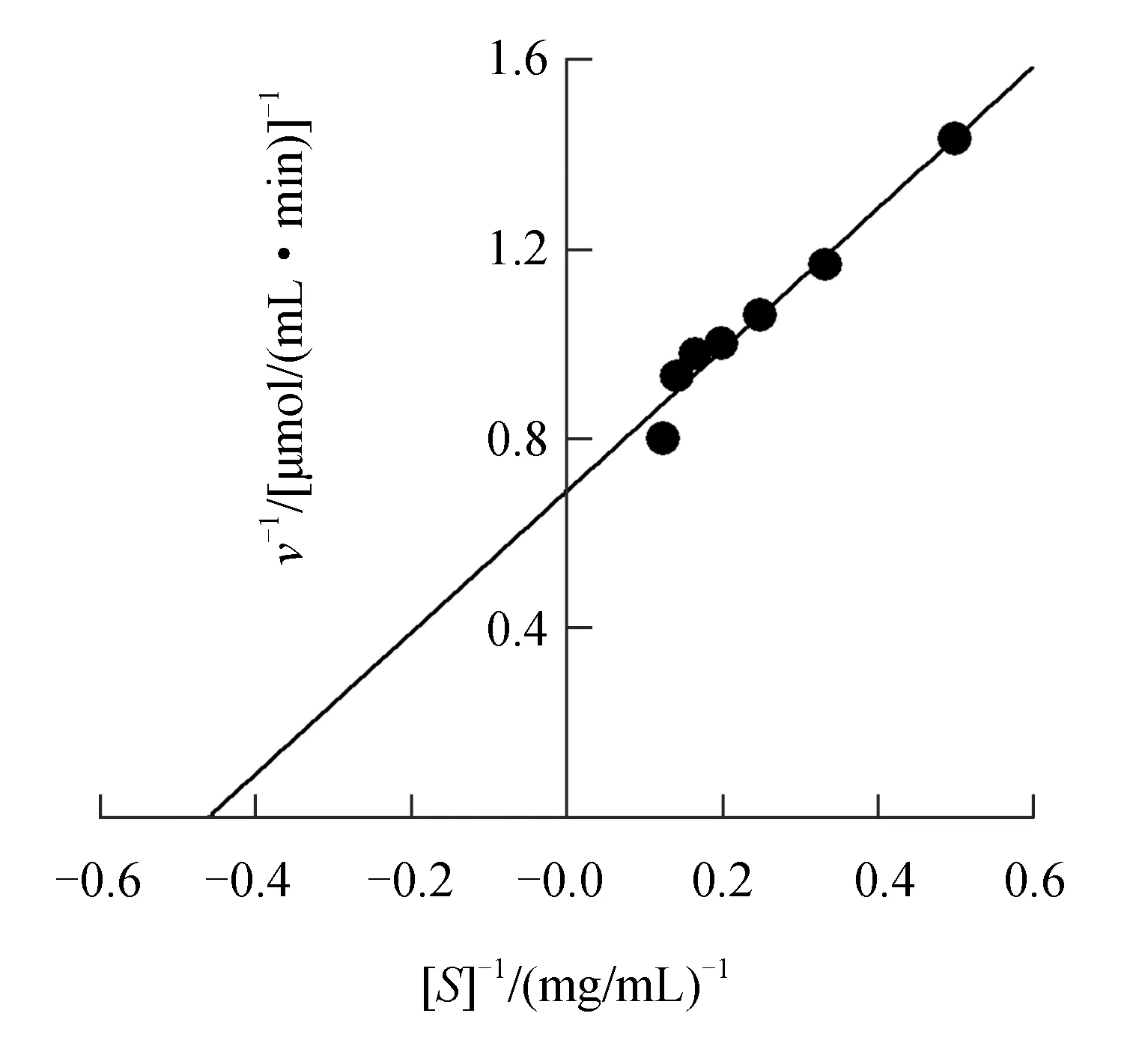
圖5 中國(guó)鱟幾丁質(zhì)酶米氏常數(shù)米氏常數(shù)的測(cè)定
2.4 金屬離子對(duì)中國(guó)鱟幾丁質(zhì)酶活力的影響
2.4.1 正一價(jià)金屬離子對(duì)酶活力的影響
2.4.2 堿土金屬離子對(duì)酶活力的影響
由圖6可見,在0~5.0 mmol/L濃度內(nèi),堿土金屬離子Ca2+和Ba2+對(duì)中國(guó)鱟幾丁質(zhì)酶有微弱的激活作用,1.0 mmol/L Ca2+和Ba2+可分別使酶活力提高10.1%、11.3%,但Mg2+對(duì)中國(guó)鱟幾丁質(zhì)酶活力影響不大。
2.4.3 過(guò)渡金屬離子對(duì)酶活力的影響
由圖7可見,過(guò)渡金屬離子Fe2+對(duì)中國(guó)鱟幾丁質(zhì)酶活力影響不大;而Co2+在低濃度時(shí)對(duì)中國(guó)鱟幾丁質(zhì)酶先表現(xiàn)為激活,后隨Co2+濃度增大逐漸轉(zhuǎn)為抑制作用,5 mmol/L Co2+可使幾丁質(zhì)酶活力提高16.3%,而20 mmol/L Co2+則可使酶活力下降14.3%。Ni2+和Mn2+對(duì)中國(guó)鱟幾丁質(zhì)酶活力均呈抑制作用,10 mmol/L的Ni2+和Mn2+可分別使幾丁質(zhì)酶活力下降27.4%和41.6%。
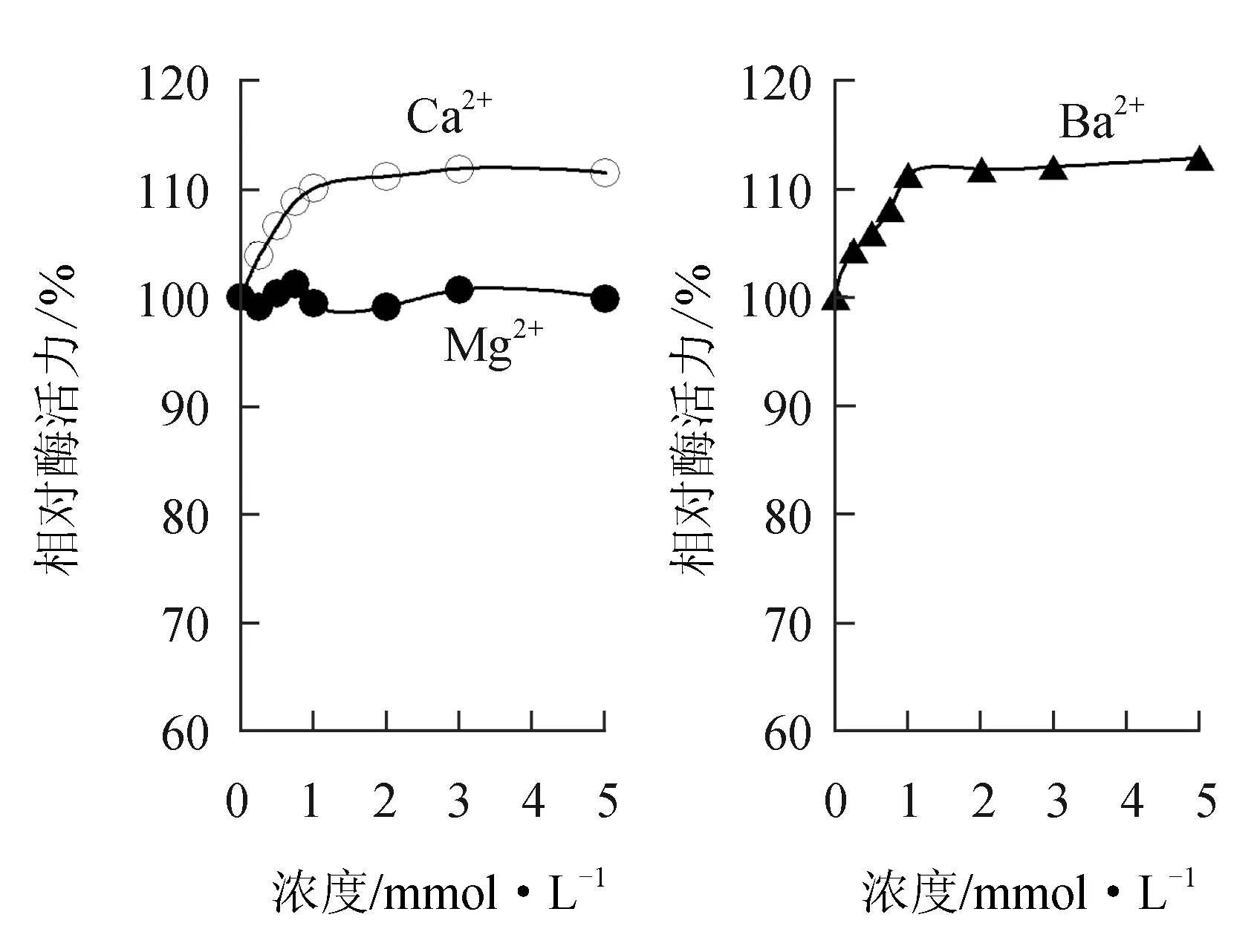
圖6 正二價(jià)堿土金屬離子對(duì)中國(guó)鱟幾丁質(zhì)酶活力的影響
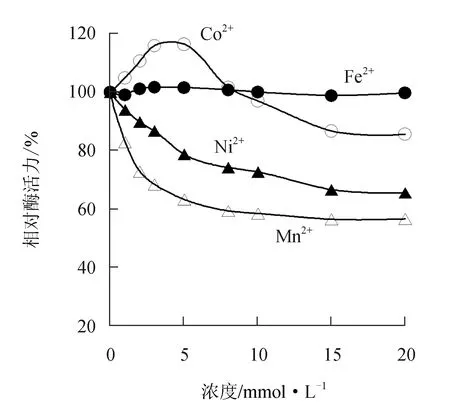
圖7 過(guò)渡金屬離子對(duì)中國(guó)鱟幾丁質(zhì)酶活力的影響
2.4.4 正三價(jià)金屬離子對(duì)酶的效應(yīng)
由圖8可見,低濃度的Fe3+對(duì)中國(guó)鱟幾丁質(zhì)酶有微弱的激活作用,但隨Fe3+濃度的增大對(duì)幾丁質(zhì)酶反而有微弱的抑制作用,5 mmol/L Fe3+能使幾丁質(zhì)酶活力提高4.3%,而20 mmol/L Fe3+對(duì)幾丁質(zhì)酶抑制了9.2%。Al3+對(duì)中國(guó)鱟幾丁質(zhì)酶活力具有一定的抑制作用,10 mmol/L的Al3+可使幾丁質(zhì)酶活力下降18.6%。
2.4.5 重金屬離子對(duì)酶活力的影響
由圖9可見,低濃度的Cu2+對(duì)中國(guó)鱟幾丁質(zhì)酶有微弱的激活作用,但隨Cu2+濃度的增大逐漸呈現(xiàn)出對(duì)幾丁質(zhì)酶的抑制作用,10 mmol/L Cu2+能使幾丁質(zhì)酶活力下降39.5%;而Cd2+、Pb2+、Zn2+和Hg2+等重金屬離子對(duì)中國(guó)鱟幾丁質(zhì)酶均呈不同程度的抑制作用,10 mmol/L的Cd2+、Pb2+、Zn2+和Hg2+可分別使幾丁質(zhì)酶活力抑制17.5%、23.9%、33.1%和83.0%,Hg2+對(duì)幾丁質(zhì)酶的抑制作用最強(qiáng)。

圖8 Fe3+和Al3+對(duì)中國(guó)鱟幾丁質(zhì)酶活力的影響
2.5 有機(jī)溶劑對(duì)中國(guó)鱟幾丁質(zhì)酶活力的影響
由表1可見,甲醇、乙醇、異丙醇和正丙醇對(duì)中國(guó)鱟幾丁質(zhì)酶均有抑制作用,抑制作用順序?yàn)榧状?乙醇>異丙醇>正丙醇;丙酮對(duì)中國(guó)鱟幾丁質(zhì)酶有一定的激活作用,10%體積分?jǐn)?shù)的丙酮會(huì)使酶活力提高16.0%;二氧六環(huán)和二甲亞砜在低濃度時(shí)對(duì)中國(guó)鱟幾丁質(zhì)酶有激活作用,隨濃度升高,對(duì)幾丁質(zhì)酶轉(zhuǎn)為抑制作用,2%體積分?jǐn)?shù)的二氧六環(huán)和二甲亞砜可分別使幾丁質(zhì)酶活力提高7.7%和14.5%,而其在30%體積分?jǐn)?shù)時(shí),可分別使幾丁質(zhì)酶活力下降11.3%和23.1%。
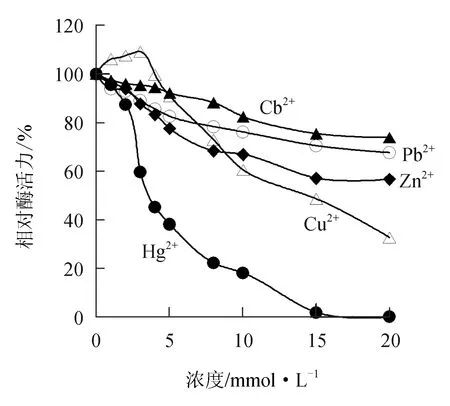
圖9 重金屬離子對(duì)中國(guó)鱟幾丁質(zhì)酶活力的影響
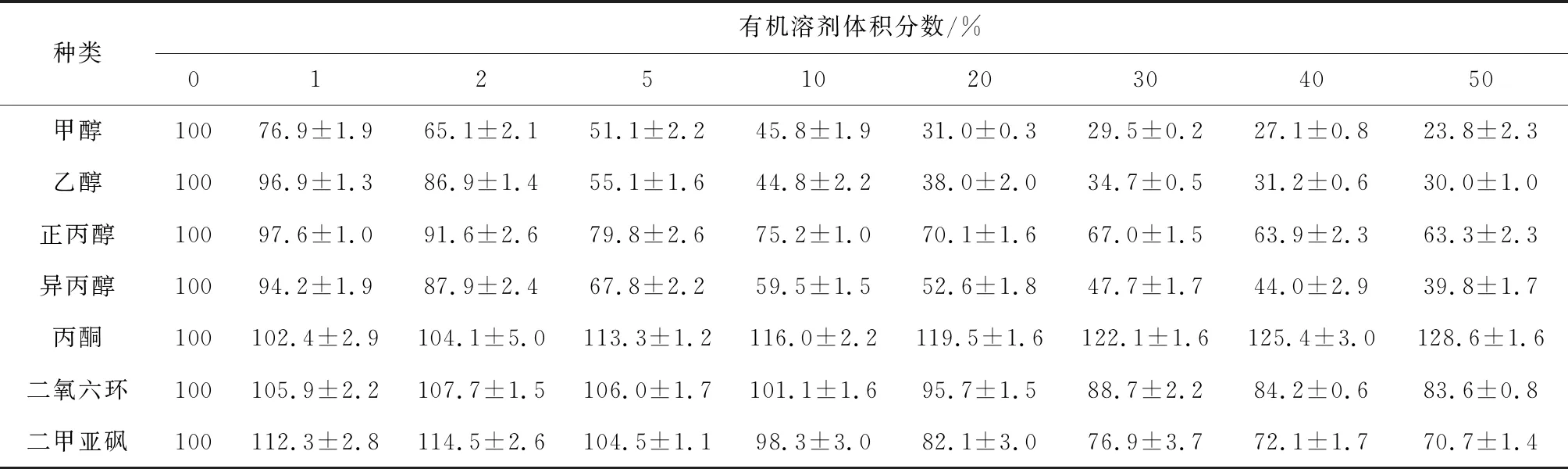
表1 7種有機(jī)溶劑對(duì)中國(guó)鱟幾丁質(zhì)酶相對(duì)酶活力的影響 %
3 討 論
3.1 幾丁質(zhì)酶的穩(wěn)定性
不同來(lái)源的幾丁質(zhì)酶對(duì)酸堿敏感度不同。凡納濱對(duì)蝦[15]幾丁質(zhì)酶酸堿穩(wěn)定性區(qū)域?yàn)閜H 4.6~7.0;而蘇云金芽孢桿菌(Bacillusthuringiensisssp.colmeri)15A3[3]幾丁質(zhì)酶B酸堿穩(wěn)定在pH 4.0~8.0。馮金榮等[17]研究的伊薩酵母(Issatchenkiaterricola)幾丁質(zhì)酶在pH 5.0~9.0內(nèi)較為穩(wěn)定,pH超出此范圍,酶將迅速失活。在本研究中,中國(guó)鱟幾丁質(zhì)酶在pH 4.6~6.0內(nèi)較為穩(wěn)定,在pH>6.5和pH<4.5下酶失活加快,對(duì)酸堿較為敏感。
蠶蛹(Bombyxmori)[18]幾丁質(zhì)酶在溫度45 ℃以上時(shí)酶失活速度加快;毛殼菌(Chaetomiumsp.)YMF1.00843[19]幾丁質(zhì)酶在溫度高于40 ℃時(shí)易失活,酶穩(wěn)定性較差。Kopparapu等[20]也從石榴(Punicagranatum)果汁中分離純化到一種熱穩(wěn)定性很好的幾丁質(zhì)酶,溫度在65 ℃內(nèi)酶基本穩(wěn)定。而中國(guó)鱟幾丁質(zhì)酶則具有較好的熱穩(wěn)定性,在20~60 ℃溫度內(nèi)酶基本穩(wěn)定,60 ℃處理1 h后酶活力還保留88%,相比中國(guó)鱟外切型幾丁質(zhì)酶——N-乙酰-β-D-氨基葡萄糖苷酶,溫度穩(wěn)定性范圍為20~50 ℃,兩者最適溫度均為55 ℃[16],因此,在鱟幼體生長(zhǎng)的適宜水溫(30 ℃左右)下,中國(guó)鱟內(nèi)切和外切幾丁質(zhì)酶活力均較低。
3.2 金屬離子對(duì)幾丁質(zhì)酶的影響
Li+和K+對(duì)海洋微生物短芽孢桿菌(Brevibacillussp.)Bsp1[21]幾丁質(zhì)酶活性有不同程度的激活作用,可分別使酶活力提高17%和3%,Na+對(duì)其又有微弱的抑制作用;而Li+對(duì)蘇云金芽孢桿菌[3]幾丁質(zhì)酶B則呈現(xiàn)一定的抑制作用,Na+對(duì)其又有微弱的激活效應(yīng)。此外,Na+和K+對(duì)中華根瘤菌(Sinorhizobiumsp.)L03[22]幾丁質(zhì)酶活性有不同程度的抑制作用;5 mmol/L Na+和K+可分別使疏綿狀嗜熱絲孢菌(Thermomyceslanuginosus)[23]幾丁質(zhì)酶活力提高45.6%和30.4%。本試驗(yàn)結(jié)果顯示,K+、Na+和Li+對(duì)中國(guó)鱟幾丁質(zhì)酶幾乎沒(méi)有影響。
堿土金屬離子常對(duì)酶呈激活效應(yīng)。Ca2+和Mg2+對(duì)毛殼菌[19]和芽孢桿菌(Bacillussp.)Hu1[24]幾丁質(zhì)酶均有激活作用;50 mmol/L Mg2+和Ba2+可顯著提高中華根瘤菌[22]幾丁質(zhì)酶的活性,Ca2+對(duì)其則呈現(xiàn)抑制作用。而Ca2+和Ba2+對(duì)凡納濱對(duì)蝦[15]幾丁質(zhì)酶活性也有一定激活作用,Mg2+對(duì)其卻沒(méi)有影響,這與本試驗(yàn)結(jié)果相似,1.0 mmol/L Ca2+和Ba2+可分別使中國(guó)鱟幾丁質(zhì)酶活力提高10.1%和11.3%,但Mg2+對(duì)酶活力影響不大。
過(guò)渡金屬離子對(duì)酶活性的影響呈現(xiàn)多樣性。Fe2+、Mn2+和Co2+對(duì)黏質(zhì)沙雷氏菌(Serratiamarcesens)[25]幾丁質(zhì)酶有不同效應(yīng),F(xiàn)e2+表現(xiàn)明顯的抑制作用,Mn2+有微弱激活作用,Co2+又幾乎沒(méi)有影響;5 mmol/L Co2+可以使短芽孢桿菌[21]幾丁質(zhì)酶活性提高1.4倍,Ni2+對(duì)其則表現(xiàn)為抑制作用;而5 mmol/L濃度的Mn2+、Co2+和Fe2+對(duì)重組幾丁質(zhì)酶rchiGR52-1[26]均有明顯的抑制作用,Ni2+對(duì)其又具有一定的促進(jìn)作用。在本試驗(yàn)中,F(xiàn)e2+對(duì)中國(guó)鱟幾丁質(zhì)酶活力影響不大,低濃度Co2+對(duì)酶表現(xiàn)為激活效應(yīng),之后隨濃度增大轉(zhuǎn)為抑制作用,Ni2+和Mn2+對(duì)酶具有不同程度的抑制作用。
Fe3+對(duì)毛殼菌[19]幾丁質(zhì)酶影響不大,對(duì)蘇云金芽孢桿菌[3]幾丁質(zhì)酶B活性有促進(jìn)作用,但又強(qiáng)烈抑制中華根瘤菌[22]幾丁質(zhì)酶的活性。在本試驗(yàn)中,低濃度Fe3+對(duì)中國(guó)鱟幾丁質(zhì)酶有微弱的激活效應(yīng),后隨濃度增大轉(zhuǎn)為抑制作用。此外,10 mmol/L Al3+可分別使蠶蛹[18]和凡納濱對(duì)蝦[15]幾丁質(zhì)酶失活5.2%和14.2%,在同樣濃度下,Al3+可使中國(guó)鱟幾丁質(zhì)酶失活18.6%。
重金屬經(jīng)常是酶的抑制劑。Zn2+、Cu2+和Hg2+能強(qiáng)烈抑制短芽孢桿菌[21]幾丁質(zhì)酶活性;10 mmol/L Cu2+、Zn2+和Cd2+可分別使黏質(zhì)沙雷氏菌[25]幾丁質(zhì)酶失活17.4%、28%和26.7%;Zn2+和Hg2+也可分別抑制疏綿狀嗜熱絲孢菌[23]幾丁質(zhì)酶活性;10 mmol/L Hg2+和Pb2+可使宛氏擬青霉(Paecilomycesvariotii)DG-3[27]產(chǎn)生的胞外幾丁質(zhì)酶(Chi46)完全失活;而凡納濱對(duì)蝦[15]幾丁質(zhì)酶在10 mmol/L的Cd2+、Pb2+和Hg2+中可分別失活11.7%、35.0%和95.6%。本試驗(yàn)結(jié)果表明,低濃度Cu2+對(duì)中國(guó)鱟幾丁質(zhì)酶表現(xiàn)為激活效應(yīng),后隨濃度增大轉(zhuǎn)為抑制作用,Cd2+、Pb2+、Zn2+和Hg2+對(duì)幾丁質(zhì)酶抑制作用順序?yàn)镠g2+>Zn2+>Pb2+>Cd2+,10 mmol/L Hg2+可使幾丁質(zhì)酶活力喪失83.0%。
目前,金屬離子對(duì)內(nèi)切型幾丁質(zhì)酶的影響機(jī)制研究報(bào)道較少,但對(duì)具有外切酶性質(zhì)的幾丁質(zhì)酶——N-乙酰β-D-氨基葡萄糖苷酶的影響機(jī)理研究較多。有研究表明,Hg2+對(duì)凡納濱對(duì)蝦[28]N-乙酰-β-D-氨基葡萄糖苷酶的抑制屬于不可逆抑制,底物對(duì)Hg2+的抑制還具有保護(hù)作用,且Hg2+是通過(guò)與半胱氨酸的巰基共價(jià)結(jié)合從而導(dǎo)致酶失活的;而Zn2+對(duì)凡納濱對(duì)蝦N-乙酰-β-D-氨基葡萄糖苷酶的抑制則表現(xiàn)為可逆的非競(jìng)爭(zhēng)性抑制作用,Zn2+可能是與N-乙酰-β-D-氨基葡萄糖苷酶蛋白的組氨酸或半胱氨酸結(jié)合,導(dǎo)致酶蛋白構(gòu)象的變化,從而引起酶的失活[29];但是,Zn2+對(duì)擬穴青蟹(Scyllaserrata)[14]N-乙酰-β-D-氨基葡萄糖苷酶的抑制則是一種可逆的混合型抑制作用;此外,Cd2+對(duì)尼羅羅非魚(Oreochromisniloticus)[30]精巢N-乙酰-β-D-氨基葡萄糖苷酶具有較強(qiáng)抑制作用,抑制類型屬于競(jìng)爭(zhēng)性抑制作用;但CuSO4對(duì)該酶的抑制則表現(xiàn)為非競(jìng)爭(zhēng)性的抑制作用[31]。
因此,不同金屬離子對(duì)幾丁質(zhì)酶有不同的影響,同一種金屬離子對(duì)不同來(lái)源的幾丁質(zhì)酶效應(yīng)也不同。
3.3 有機(jī)溶劑對(duì)幾丁質(zhì)酶的影響
酶依賴其活性中心與底物結(jié)合并產(chǎn)生催化反應(yīng),有機(jī)溶劑能滲透入酶的活性中心,降低活性中心的極性,從而增加酶與底物的靜電斥力,降低了酶與底物的結(jié)合能力。因?yàn)椴煌袡C(jī)溶劑的極性大小不同,必然對(duì)酶呈現(xiàn)不同的影響效應(yīng)。本試驗(yàn)結(jié)果顯示,甲醇、乙醇、異丙醇和正丙醇等4種有機(jī)溶劑對(duì)中國(guó)鱟幾丁質(zhì)酶均呈現(xiàn)抑制作用,其抑制作用的強(qiáng)弱與4種有機(jī)溶劑的極性大小(甲醇>乙醇>異丙醇>正丙醇)呈正相關(guān),其中甲醇極性大,對(duì)幾丁質(zhì)酶的抑制作用最強(qiáng)。甲醇、乙醇和正丙醇對(duì)凡納濱對(duì)蝦[13]體壁幾丁質(zhì)酶也有類似的抑制作用。
丙酮對(duì)蘇云金芽孢桿菌[32]幾丁質(zhì)酶活力有較強(qiáng)的激活作用,激活效果在2.5%~25%的體積分?jǐn)?shù)內(nèi)與體積分?jǐn)?shù)成正相關(guān);而在0~50%體積分?jǐn)?shù)內(nèi),丙酮對(duì)凡納濱對(duì)蝦[13]幾丁質(zhì)酶也呈激活作用。本試驗(yàn)中,丙酮也同樣具有促進(jìn)中國(guó)鱟幾丁質(zhì)酶活力的作用。丙酮是非質(zhì)子極性溶劑,酶蛋白與溶劑之間直接接觸,蛋白質(zhì)分子的表面結(jié)構(gòu)有所變化,這些變化可能使幾丁質(zhì)酶活性中心的柔性增強(qiáng),有利于酶功能的發(fā)揮。
有研究表明,低體積分?jǐn)?shù)的二氧六環(huán)和二甲亞砜對(duì)凡納濱對(duì)蝦[13]幾丁質(zhì)酶有激活作用,且隨體積分?jǐn)?shù)升高激活作用減弱并逐漸轉(zhuǎn)為抑制作用,與本試驗(yàn)中二氧六環(huán)和二甲亞砜對(duì)中國(guó)鱟幾丁質(zhì)酶的影響效應(yīng)相似。二甲亞砜對(duì)蘇云金芽孢桿菌[32]幾丁質(zhì)酶也表現(xiàn)為先激活后抑制的效應(yīng);但是,二氧六環(huán)在體積分?jǐn)?shù)小于5%時(shí),對(duì)蘇云金芽孢桿菌[32]幾丁質(zhì)酶的影響不明顯,當(dāng)體積分?jǐn)?shù)大于5%時(shí),對(duì)酶開始呈現(xiàn)激活作用。
猜你喜歡
中學(xué)生數(shù)理化·八年級(jí)物理人教版(2022年3期)2022-03-16 05:55:08
當(dāng)代陜西(2021年2期)2021-03-29 07:41:24
家庭影院技術(shù)(2020年10期)2020-12-14 07:54:18
媽媽寶寶(2017年3期)2017-02-21 01:22:28
中國(guó)塑料(2016年3期)2016-06-15 20:30:00
通信電源技術(shù)(2016年3期)2016-03-26 07:13:38
知識(shí)經(jīng)濟(jì)·中國(guó)直銷(2016年3期)2016-02-27 16:15:49
現(xiàn)代檢驗(yàn)醫(yī)學(xué)雜志(2014年6期)2014-02-02 03:02:04
閱讀與作文(小學(xué)低年級(jí)版)(2011年3期)2011-01-01 00:00:00
