大白菜微管與灰霉菌抗性研究
2019-09-23 06:10:53陳曉峰叢山王百川
江蘇農(nóng)業(yè)科學(xué) 2019年14期
關(guān)鍵詞:植物
陳曉峰 叢山 王百川
摘要:對(duì)灰霉菌(Botrytis cinerea)誘導(dǎo)抗、感大白菜葉片α-微管蛋白基因誘導(dǎo)表達(dá)的特點(diǎn)及微管動(dòng)態(tài)結(jié)構(gòu)變化進(jìn)行研究。結(jié)果表明,灰霉菌接種后大白菜中4種α-微管蛋白基因均具有上調(diào)表達(dá)特點(diǎn),且抗病品種較感病品種誘導(dǎo)表達(dá)明顯。葉片細(xì)胞微管免疫熒光觀測發(fā)現(xiàn),接種48、96 h后抗病品種較感病品種微管骨架受病原菌侵染影響較小,結(jié)構(gòu)變化不明顯。研究結(jié)果說明微管在植物對(duì)抗真菌病原菌侵染中具有一定的作用。
關(guān)鍵詞:大白菜;灰霉菌;葉片細(xì)胞;微管;α-微管蛋白;互作動(dòng)態(tài);結(jié)構(gòu)變化;作用機(jī)制
中圖分類號(hào):S436.341 ? 文獻(xiàn)標(biāo)志碼: A ?文章編號(hào):1002-1302(2019)14-0124-03
微管是真核生物細(xì)胞的重要組成部分,它和微絲、中間纖維組成植物的細(xì)胞骨架結(jié)構(gòu),在細(xì)胞形態(tài)發(fā)生、信號(hào)識(shí)別、細(xì)胞極性的建立等方面起著至關(guān)重要的作用[1-3]。在植物生長發(fā)育過程中,微管蛋白還可以調(diào)節(jié)植物細(xì)胞以不同的形態(tài)來適應(yīng)環(huán)境和功能需求,同時(shí)對(duì)植物纖維素和木質(zhì)素合成,構(gòu)建完整細(xì)胞骨架均具有重要作用[4]。植物微管蛋白主要由α-微管蛋白和β-微管蛋白基因家族組成[4-8],其貫穿整個(gè)細(xì)胞生長發(fā)育的全過程,參與微管的生長發(fā)育和動(dòng)力學(xué)變化[9-12]。
最近幾年,關(guān)于微管骨架在植物抗性研究中的作用越來越受到重視,人們對(duì)微管骨架參與植物和病原菌互作中的作用進(jìn)行了廣泛研究[13-25],發(fā)現(xiàn)通過化學(xué)物質(zhì)解聚細(xì)胞微管可阻礙過敏性壞死反應(yīng)(hypersensitive response,簡稱HR)發(fā)生,加劇病原菌的入侵,降低植物抗病力[13,22-23]。在病原菌和外源化學(xué)物質(zhì)誘導(dǎo)下,α-微管基因表達(dá)也不盡相同[2,14-17,23-25],是否該基因家族全部成員都參與了植物抗性反應(yīng)目前尚不清楚。
本研究對(duì)不同抗性大白菜材料α-微管蛋白基因受病原菌誘導(dǎo)表達(dá)的特點(diǎn)進(jìn)行分析,對(duì)微管骨架在寄主與病原菌互作中的動(dòng)態(tài)結(jié)構(gòu)變化進(jìn)行觀察,以期闡明微管在抗真菌病害體系中的作用。
1 材料與方法
1.1 材料
試驗(yàn)于2016—2018年在中國農(nóng)業(yè)大學(xué)煙臺(tái)研究院進(jìn)行。大白菜抗真菌病害品種為新煙雜3號(hào)(XYZ),感病品種為包頭蓮(BTL)。大白菜種子經(jīng)消毒后,于人工氣候箱在20~25 ℃、光周期12 h/d、相對(duì)濕度(85±5)%條件下培養(yǎng),待幼苗長至2~4張真葉時(shí)用于試驗(yàn)。
免疫熒光觀測所用果膠酶和纖維素酶,微管標(biāo)記抗體一抗為抗α-微管蛋白IgG,二抗為FITC結(jié)合的羊抗鼠IgG,均采用Sigma公司產(chǎn)品。
1.2 灰霉菌接種液的制備和接種程序
灰霉菌(Botrytis cinerea)接種液的制備和接種參照馬清華等的方法[26-27],最終配成的孢子懸浮接種液濃度為1×106個(gè)/mL。處理組每個(gè)單株葉片噴灑病原菌誘導(dǎo)液,對(duì)照組用純水代替病原菌誘導(dǎo)液。誘導(dǎo)處理后植株在100%相對(duì)濕度下保濕24 h,然后揭掉遮光物,于人工氣候箱在20~25 ℃、光周期12 h/d、相對(duì)濕度(85±5)%條件下培養(yǎng),直至采樣結(jié)束,每個(gè)處理設(shè)3個(gè)重復(fù)。
1.3 實(shí)時(shí)定量PCR檢測灰霉菌誘導(dǎo)α-微管蛋白基因表達(dá)
1.3.1 實(shí)時(shí)定量PCR反應(yīng)引物設(shè)計(jì) 在GenBank大白菜數(shù)據(jù)庫中選取所需要的α-微管蛋白基因,引物設(shè)計(jì)參考Zhang等的研究[2],大白菜actin基因?yàn)閮?nèi)標(biāo)基因,用于實(shí)時(shí)定量PCR的反應(yīng)引物序列見表1。
1.3.2 實(shí)時(shí)定量PCR反應(yīng)體系和反應(yīng)程序 采用Takara公司SYBR Premix Ex Taq試劑盒進(jìn)行實(shí)時(shí)定量PCR反應(yīng)。提取處理和對(duì)照葉片0、12、24、36、48、72、96 h的大白菜葉片RNA,利用反轉(zhuǎn)錄酶將mRNA合成cDNA。實(shí)時(shí)定量PCR反應(yīng)體系和反應(yīng)程序參考陳曉峰等的方法[28],每個(gè)處理設(shè)3次重復(fù)。
1.4 大白菜葉片細(xì)胞微管間接免疫熒光檢測
分別采集灰霉菌誘導(dǎo)接種0、48、96 h抗感病大白菜植株葉片,參考張靜宜等的方法[3]進(jìn)行微管免疫熒光觀測。
2 結(jié)果與分析
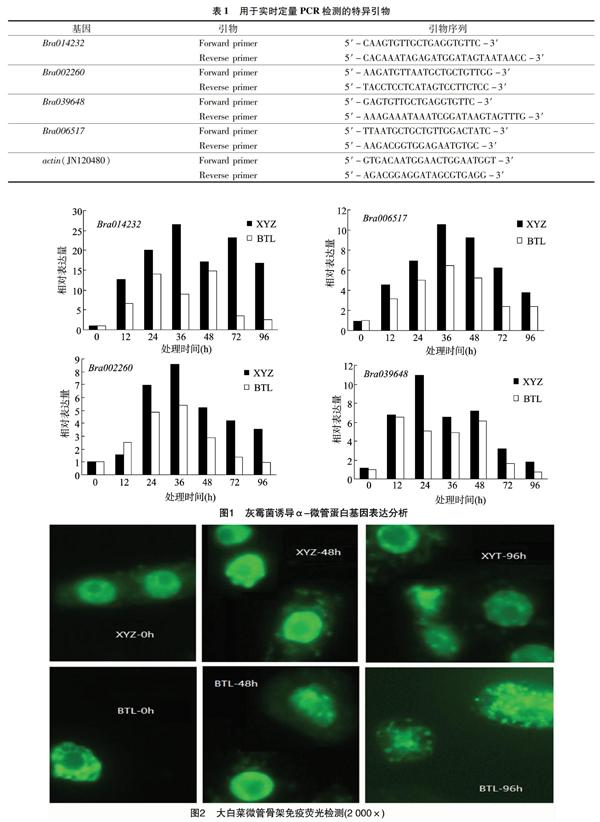
2.1 灰霉菌誘導(dǎo)α-微管蛋白基因表達(dá)分析
由圖1可知,4個(gè)α-微管蛋白基因均呈現(xiàn)誘導(dǎo)表達(dá)特點(diǎn),但表達(dá)特點(diǎn)各不相同。4個(gè)基因在抗病品種中的表達(dá)均高于感病品種,其中Bra006517和Bra002260基因呈現(xiàn)單峰表達(dá)特點(diǎn),Bra014232基因在所有試驗(yàn)材料中呈現(xiàn)雙峰表達(dá)特點(diǎn)。4個(gè)α-微管蛋白基因均在誘導(dǎo)24~48 h內(nèi)出現(xiàn)峰值表達(dá)量,其中Bra014232基因表現(xiàn)出明顯的誘導(dǎo)表達(dá)特點(diǎn),Bra002260、Bra006517、Bra039648呈現(xiàn)較弱的上調(diào)表達(dá)特點(diǎn)。感病品種中4個(gè)基因在誘導(dǎo)48 h后表達(dá)量明顯下降。
2.2 間接免疫熒光觀測大白菜葉片微管結(jié)構(gòu)差異
由圖2可知,灰霉菌誘導(dǎo)處理后,抗、感病品種葉片中微管顯微結(jié)構(gòu)觀測結(jié)果表現(xiàn)出明顯的差異,在整個(gè)誘導(dǎo)過程中,抗病品種XYZ的葉片細(xì)胞中細(xì)胞核周圍微管骨架結(jié)構(gòu)變化不明顯,部分細(xì)胞微管在病原菌誘導(dǎo)96 h后結(jié)構(gòu)出現(xiàn)不完整現(xiàn)象。感病品種BTL葉片細(xì)胞在病原菌誘導(dǎo)48 h后微管骨架結(jié)構(gòu)出現(xiàn)分散現(xiàn)象,病原菌誘導(dǎo)96 h后,絕大部分微管結(jié)構(gòu)出現(xiàn)分散降解現(xiàn)象。
3 結(jié)論與討論
微管蛋白由于在保持細(xì)胞結(jié)構(gòu)、胞內(nèi)物質(zhì)運(yùn)輸、細(xì)胞發(fā)育等過程中具有重要作用,人們對(duì)其在植物抗病體系中的作用進(jìn)行了廣泛研究。關(guān)于微管骨架與植物抗病性關(guān)系研究主要集中在病原菌與寄主過敏性壞死反應(yīng)和抗性化學(xué)物質(zhì)累積等方面[9]。研究發(fā)現(xiàn),通過oryzalin等已知的誘導(dǎo)微管解聚的化學(xué)物質(zhì)處理植物材料,這些植物對(duì)外源病原菌侵染誘導(dǎo)過敏反應(yīng)細(xì)胞的產(chǎn)生率降低,抗性降低[9,16,23]。正常條件下植物微管呈網(wǎng)狀結(jié)構(gòu)圍繞在細(xì)胞周圍,病原菌侵染植物組織后,免疫熒光觀測發(fā)現(xiàn)微管骨架分散聚集于侵染點(diǎn)或接種點(diǎn)周圍[14-18],說明微管直接或者間接參與了植物的抗性反應(yīng)。
本研究對(duì)大白菜中4種α-微管蛋白基因進(jìn)行了誘導(dǎo)表達(dá)分析,這4個(gè)基因在大白菜不同組織中存在特異表達(dá)的特點(diǎn),且受茉莉酸甲酯(MJ)、紫杉醇、赤霉素(GA3)等化學(xué)物質(zhì)誘導(dǎo)[2]。本研究中筆者所在課題組發(fā)現(xiàn)4個(gè)α-微管蛋白基因均受灰霉菌誘導(dǎo),呈上調(diào)表達(dá)特點(diǎn),且抗病品種中微管蛋白基因表達(dá)量略高于感病品種,個(gè)別基因在抗病品種中出現(xiàn)雙峰表達(dá)特點(diǎn),這都說明微管蛋白基因在抗病品種中參與了大白菜對(duì)真菌病原菌侵染的抗性反應(yīng)。對(duì)不同抗性材料葉片細(xì)胞中微管骨架的免疫熒動(dòng)態(tài)觀測發(fā)現(xiàn),抗病品種葉片細(xì)胞中微管骨架結(jié)構(gòu)比較完整,只有少量細(xì)胞出現(xiàn)微管降解現(xiàn)象,而感病品種在誘導(dǎo)處理2 d后微管骨架結(jié)構(gòu)出現(xiàn)分散降解,微管降解導(dǎo)致植物細(xì)胞結(jié)構(gòu)變化,說明微管骨架結(jié)構(gòu)完整對(duì)真菌病原菌與寄主抗性反應(yīng)的重要性。
未來須要著重研究整個(gè)微管基因家族組織和誘導(dǎo)表達(dá)特性,同時(shí)完善病原菌誘導(dǎo)植物細(xì)胞微管骨架的動(dòng)態(tài)研究。
參考文獻(xiàn):
[1]Chan J,Eder M,Crowell E F,et al. Microtubules and CESA tracks at the inner epidermal wall align independently of those on the outer wall of light-grown Arabidopsis hypocotyls[J]. Journal of Cell Science,2011,124(7):1088-1094.
[2]Zhang Y W,Jin D,Xu C,et al. Regulation of bolting and identification of the α-tubulin gene family in Brassica rapa L. ssp pekinensis[J]. Genetics and Molecular Research,2016,15(1):15017507.
[3]張靜宜,侯喜林,史公軍,等. 白菜組織細(xì)胞微管間接免疫熒光檢測體系的優(yōu)化[J]. 園藝學(xué)報(bào),2007,34(6):1551-1554.
[4]饒國棟,張建國. 植物微管蛋白基因研究進(jìn)展[J]. 世界林業(yè)研究,2013,26(3):17-20.
[5]Kopczak S D,Haas N A,Hussey P J,et al. The small genome of Arabidopsis contains at least six expressed α-tubulin genes[J]. Plant Cell,1992,4(5):539-547.
[6]Parrotta L,Cai G,Cresti M. Changes in the accumulation of α- and β-tubulin during bud development in Vitis vinifera L.[J]. Planta,2010,231(2):277-291.
[7]Oakley R V,Wang Y S,Ramakrishna W,et al. Differential expansion and expression of α- and β-tubulin gene families in Populus[J]. Plant Physiology,2007,145(3):961-973.
[8]Dixon D C,Seagull R W,Triplett B A. Changes in the accumulation of α- and β-tubulin isotypes during cotton fiber development[J]. Plant Physiology,1994,105(4):1347-1353.
[9]Nogales E . A structural view of microtubule dynamics[J]. Cellular & Molecular Life Sciences Cmls,1999,56(1/2):133-142.
[10]Nogales E,Wolf S G,Downing K H. Structure of the alpha-beta tubulin dimer by electron crystallography[J]. Nature,1998,391(6663):199-203.
[11]Nogales E,Whittaker M,Milligan R A,et al. High-resolution model of the microtubule[J]. Cell,1999,96(1):79-88.
[12]Carpenter J L,Ploense S E,Snustad D P,et al. Preferential expression of an α-tubulin gene of Arabidopsis in pollen[J]. Plant Cell,1992,4(5):557-571.
[13]姚 晶,禹 坷,陳艷利,等. 微管骨架在辣椒-黃瓜炭疽病菌非寄主互作中的作用[J]. 植物病理學(xué)報(bào),2013,43(2):136-142.
[14]Skalamera D,Heath M C. Changes in the cytoskeleton accompanying infection-induced nuclear movements and the hypersensitive response in plant cells invaded by rust fungi[J]. Plant Journal,1998,16(2):191-200.
[15]Schmelzer E. Cell polarization,a crucial process in fungal defense[J]. Trends in Plant Science,2002,7(9):411-415.
[16]Christopher-Kozjan R,Heath M C. Cytological and pharmacological evidence that biotrophic fungi trigger different cell death execution processes in host and non-host cells during the hypersensitive response[J]. Physiological and Molecular Plant Pathology,2003,62(5):265-275.
[17]Hardham A R,Jones D A,Takemoto D. Cytoskeleton and cell wall function in penetration resistance[J]. Current Opinion in Plant Biology,2007,10(4):342-348.
[18]Kobayashi I,Kobayashi Y,Yamaoka N,et al. Recognition of a pathogen and a nonpathogen by barley coleoptile cells. Ⅲ. Responses of microtubules and actin filaments in barley coleoptile cells to penetration attempts[J]. Canadian Journal of Botany,1992,70(9):1815-1823.
[19]Takemoto D,Hardham A R. The cytoskeleton as a regulator and target of biotic interactions in plants[J]. Plant Physiology,2004,136(4):3864-3876.
[20]Takemoto D,Jones D A,Hardham A R. GFP-tagging of cell components reveals the dynamics of subcellular reorganization in response to infection of Arabidopsis by oomycete pathogens[J]. Plant Journal,2003,33(4):775-792.
[21]Münch S,Lingner U,F(xiàn)loss D S,et al. The hemi-biotrophic lifestyle of Colletotrichum species[J]. Journal of Plant Physiology,2008,165(1):41-51.
[22]左 海,王海燕,馬 青. 微管骨架在小麥抗條銹菌侵染中作用的研究[C]. 中國植物病理學(xué)會(huì)2012年學(xué)術(shù)年會(huì). 北京:中國農(nóng)業(yè)科學(xué)技術(shù)出版社,2012.
[23]郝心愿,李紅莉,禹 珂,等. 微絲骨架解聚劑在小麥-黃瓜白粉菌非寄主互作中的作用[J]. 中國農(nóng)業(yè)科學(xué),2011,44(2):291-298.
[24]Li H L,Wang H Y,Hao X Y,et al. Effects of microtubule polymerization inhibitor on the hypersensitive response of wheat induced by the non-host pathogen Sphaerotheca fuliginea[J]. Agricultural Sciences in China,2010,9(3):378-382.
[25]Song X H,Ma Q,Hao X Y,et al. Roles of the actin cytoskeleton and an actin-binding protein in wheat resistance against Puccivia striiformis f. sp. tritici[J]. Protoplasma,2012,249(1):99-106.
[26]馬清華,陳曉峰,牟晉華,等. 灰霉菌侵染大白菜后ADF7和ADF10基因表達(dá)分析[J]. 南京農(nóng)業(yè)大學(xué)學(xué)報(bào),2015,38(5):742-747.
[27]王利英. 不結(jié)球白菜黑斑病抗性機(jī)制初步研究[D]. 南京:南京農(nóng)業(yè)大學(xué),2008:40.
[28]陳曉峰,隋好林,馬清華,等. 霜霉病菌誘導(dǎo)大白菜幾丁質(zhì)酶和葡聚糖酶基因的表達(dá)[J]. 山東農(nóng)業(yè)科學(xué),2015,47(2):96-99.
猜你喜歡
少兒科學(xué)周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學(xué)閱讀指南·低年級(jí)版(2021年3期)2021-03-19 06:12:40
小天使·二年級(jí)語數(shù)英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級(jí)語數(shù)英綜合(2020年11期)2020-12-16 02:57:22
學(xué)苑創(chuàng)造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領(lǐng)巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
