外來種克氏原螯蝦(Procambarus clarkii)對3種沉水植物的牧食研究
2019-09-24 07:31:56袁野張尹哲呂超超馬徐發夏成星
生態科學 2019年5期
袁野, 張尹哲, 呂超超,2,3, 馬徐發,2,3, 夏成星,2,3,*
外來種克氏原螯蝦()對3種沉水植物的牧食研究
袁野1, 張尹哲1, 呂超超1,2,3, 馬徐發1,2,3, 夏成星1,2,3,*
1. 華中農業大學水產學院, 農業部淡水生物繁育重點實驗室, 武漢 430070 2. 淡水水產健康養殖湖北省協同創新中心, 武漢 430070 3. 池塘健康養殖湖北省工程實驗室, 武漢 430070
為探討外來種克氏原螯蝦()對沉水植物牧食的偏好性和牧食強度, 采用投喂實驗的方式, 研究了3種沉水植物刺苦草()、輪葉黑藻()和伊樂藻()的適口性。結果表明, 克氏原螯蝦對輪葉黑藻的取食速率最大(7.24±0.24 mg·d-1), 其次是苦草(3.70±1.14 mg·d-1), 伊樂藻最小(0.60±0.12 mg·d-1), 說明伊樂藻的適口性最差, 輪葉黑藻的適口性最好, 苦草的適口性居中。3種沉水植物的纖維素、多酚含量和氮含量都沒有顯著性差異, 而伊樂藻卻具有更高的碳(C)含量和更高的碳氮比(C:N)。總體來說, 3種沉水植物的物理結構、碳(C)含量和碳氮比(C:N)在外來種克氏原螯蝦對沉水植物的牧食中具有重要的作用。
克氏原螯蝦; 沉水植物; 牧食; 適口性; 碳氮比
0 前言
克氏原螯蝦()俗稱小龍蝦, 隸屬于節肢動物門(Arthropoda)、甲殼綱(Crustacea)、十足目(Decapoda)、螯蝦科(Cambaridae), 原產于墨西哥東北部和美國南部地區, 是世界分布最廣泛的淡水螯蝦入侵種類[1–3], 現已廣泛分布于我國中東部地區的各種淡水水體, 尤其是長江中下游地區[4–9]。克氏原螯蝦具有較強的生態可塑性, 其生態幅寬、適應性廣, 具有食性雜、生長快、成熟早、繁殖力高等生物學特性, 使得克氏原螯蝦可以在入侵水體迅速建立種群[10–12]。克氏原螯蝦作為淡水水體重要的消費者, 對水體初級生產者和次級消費者的生物多樣性和群落結構具有較大的影響[13–15], 其可以通過取食或者挖掘等行為, 使大型水生植物大量減少, 從而可使水體由周叢藻類和沉水植物為主的清水狀態轉變為以浮游植物為主的渾濁狀態[16–17]。
植物適口性(Plant palatability)是植物一個重要的多功能性狀的綜合生態特征, 決定了牧食者的牧食選擇和植物可能承受的牧食壓力[18], 主要包括植物組織的物理性狀(Physical characteristics)和化學性狀(Chemical characteristics), 物理性狀主要包括植物體的外層結構和一些特殊的防御結構; 化學性狀主要是植物體內的營養物質含量、次生代謝產物(酚、生物堿、類三萜和皂角苷等)的含量[19–21]。植物中營養物質含量(C、N)和碳氮比(C:N)也是評價植物適口性的重要依據[22–23], 植物體具高氮(N)、低碳(C)和低碳氮比(C:N), 其適口性相對較高[24]。Bakker等[25]研究沉水植物化學計量特征對適口性影響時, 發現氮(N)含量高的植物具有更好的適口性, 且水生植物組織含氮(N)量和水生動物對其取食消耗成正相關。除了元素特征差異決定了植物的適口性以外, 纖維素和多酚含量與植物的適口性也密切相關。纖維素是植物細胞壁的主要組成成分, 纖維素含量增加, 可以使植物葉片的韌性增加, 降低植物的適口性[26]。朱順清等[27]在草食性魚類對水草的牧食研究中, 發現纖維素含量較低的苦草、輪葉黑藻和菹草比纖維素含量高的黃絲草具有更高的適口性, 更易于魚類的消化吸收, 草食性魚類也更為喜食。多酚作為廣泛存在于植物各器官中的一種復雜酚類的次生代謝產物, 含量僅次于纖維素、半纖維素和木質素, 可作為化學防御物質來抵御牧食者的牧食[22,28]。入侵歐洲和美洲地區的穗狀狐尾藻體內具有較多的酚類物質, 其高濃度的酚類不僅可以降低本身的適口性抵御牧食者牧食, 還可以使鱗翅目的幼蟲生長速度降低[29–30]。
克氏原螯蝦主要通過取食和挖掘等行為來影響水生植物的生長和群落演替, 并且受到水深和捕食者的影響[17,31–33]。Carreira等[31]在克氏原螯蝦對5種水生植物(,,,,)的牧食實驗中, 發現克氏原螯蝦更喜食, 而不喜食和, 不同種類植物的取食消耗和斷枝數量主要取決于克氏原螯蝦的大小而并非性別, 但對造成取食速率差異和植物適口性差異的機制并未進行深入探討。國內多數研究主要集中于克氏原螯蝦的人工養殖技術、食物組成、攝食節律和肌肉品質[34–36], 對克氏原螯蝦的入侵生態學研究也相對較少[7–10,37], 對克氏原螯蝦對沉水植物的牧食選擇性研究更少[38]。
本研究主要通過適口性實驗, 比較分析了入侵物種克氏原螯蝦對3種水鱉科沉水植物的取食速率以及沉水植物C、N元素和防御物質對牧食的影響, 探討了克氏原螯蝦對3種沉水植物的牧食選擇性差異及其原因, 可以為牧食理論的研究、沉水植被恢復和沉水植物過度生長控制提供數據支持和理論依據, 為克氏原螯蝦的防控提供一定的參考。
1 材料與方法
1.1 實驗材料
本研究所用沉水植物3種, 隸屬于水鱉科(Hydro-charitaceae), 分別為刺苦草(V)、輪葉黑藻()和伊樂藻(), 其中伊樂藻為外來種。所用水生植物均采集于武漢市青菱湖, 采集清洗干凈后, 培養于基質為沙子的水族缸中。實驗開始時, 取刺苦草的葉片、輪葉黑藻和伊樂藻的頂枝(均為20 cm)用清水漂洗干凈、備用。
實驗用克氏原螯蝦采集于青菱湖及周邊池塘、溝渠, 采集后帶回實驗基地室內暫養, 為防止克氏原螯蝦互相打斗受傷, 分別單獨放入具蓋塑料桶(防止逃逸)中, 并用沉水植物投喂, 定期換水, 更換植物。實驗開始前, 挑取大小相對一致、生命力強的雌性克氏原螯蝦進行實驗(保持性別一致), 實驗開始之前饑餓處理24 h。
1.2 實驗設計
本實驗在華中農業大學實驗基地室內進行。實驗時間為2016年9月11日-9月18日, 實驗周期為1周。實驗設置2個處理組(牧食組和對照組), 3個重復, 牧食組內放入1株植物和1只克氏原螯蝦(體重: (28.53±2.92) g, Mean±SE), 各處理組克氏原螯蝦大小沒有顯著差異(2,8= 0.075;= 0.929), 對照組內僅放入頂枝, 用于實驗周期內沉水植物生長指標的計算。實驗選取3種水生植物頂枝(苦草為葉片), 每株20 cm, 且每種留取大小相似的頂枝或葉片20個, 烘干、稱重計算干濕比。實驗容器為錐形塑料盒(上部直徑×下部直徑×高, 17 cm×13 cm×8.5 cm), 水深為6.5 cm。每個容器中放置帶孔盒蓋防止克氏原螯蝦逃逸。
1.3 日取食量的測定
投喂實驗開始前, 稱量每組植物鮮重和克氏原螯蝦體重, 1周實驗結束后, 將克氏原螯蝦取出后用撈網和鑷子將取食后的植物碎屑和空白組內植物取出, 晾置數分鐘后稱取鮮重。
采用Carreira[31]的方法計算克氏原螯蝦對沉水植物的日取食量, 即取食速率, 比較水生植物的適口性。
取食速率(Daily consumed biomass,B)按下式計算:
B= (W×–W)×W
式中,W為初始鮮重;為空白對照組頂枝的自生長率(實驗結束時鮮重/初始鮮重);W為實驗組結束時的鮮重;W為頂枝的干濕比。
1.4 植物化學成分的測定
用Folin-Ciocaltau方法[39]測定植物多酚含量, 取少量各處理組3種沉水植物葉子和莖烘干并磨成粉末, 然后取大約2.0—8.0 mg用來測定多酚。
稱取4.0 mg植物粉末樣品采用蒽酮比色法[40]測定纖維素含量。取大約4.0 mg的植物粉末, 利用CHNS/O元素分析儀(Elementar Vario PYRO cube, German), 采用杜馬斯燃燒法進行碳、氮連續分析, 測定碳、氮元素含量, 每個植株樣品測量3次, 每個處理測量3個重復。
1.5 數據分析
在數據分析前, 為滿足數據的正態分布和方差的齊次性, 所有的數據都經過log轉換。利用單因素方差分析(One-way ANOVA), 對克氏原螯蝦取食速率和3種沉水植物多酚、纖維素、碳含量、氮含量和碳氮比(C/N)進行顯著性分析, 利用Turkey's HSD的方法對不同處理間差異進行多重比較。數據分析采用SPSS 24.0統計分析軟件進行。
2 結果與分析
2.1 克氏原螯蝦對3種沉水植物的牧食
克氏原螯蝦對3種沉水植物的取食速率具有顯著性差異(<0.01)(圖1a), 其取食速率大小為: 輪葉黑藻>苦草>伊樂藻, 實驗周期內鮮重取食量分別為(2.39±0.57) g 、(1.89±0.27) g和(0.61±0.07) g(表1), 取食速率分別為(7.24±0.24) mg·d-1、(3.70±1.14) mg·d-1和(0.60±0.12) mg·d-1, 表明克氏原螯蝦更喜食輪葉黑藻, 其適口性最好, 而伊樂藻的適口性最差。
2.2 3種沉水植物的纖維素和多酚含量
3種沉水植物的纖維素和多酚含量沒有顯著性差異(>0.05)(圖1b,c), 輪葉黑藻、刺苦草、伊樂藻的纖維素含量分別為(128.32±12.49) mg·g-1、(126.02±29.58) mg·g-1和(149.30±9.63) mg·g-1, 多酚含量分別為(3.11±0.39) mg·g-1、(2.44±0.23) mg·g-1和(2.70±0.23) mg·g-1, 表明纖維素和多酚不是影響克氏原螯蝦對3種沉水植物牧食的主要因素。

表1 克氏原螯蝦牧食前后3種沉水植物的鮮重(Mean±SE)
2.3 3種沉水植物的碳氮含量
3種沉水植物之間的C含量和C:N比值之間具有顯著性差異, N含量無顯著性差異(圖1d,e,f)。輪葉黑藻、刺苦草和伊樂藻的碳(C)含量分別為(359.00±1.49) mg·g-1、(333.21±3.35) mg·g-1和(379.28±3.84) mg·g-1; 氮(N)含量分別為(34.90±0.96) mg·g-1、(31.62±1.93) mg·g-1和(28.95±1.47) mg·g-1; C:N比值分別為(10.30±0.24)、(10.62±0.64)和(13.16± 0.59)。伊樂藻的C含量顯著高于輪葉黑藻和刺苦草, 而伊樂藻的C:N比值顯著高于輪葉黑藻和刺苦草, 但輪葉黑藻和刺苦草之間C:N比值卻沒有顯著性差異。
3 討論
3.1 克氏原螯蝦對3種沉水植物的牧食
克氏原螯蝦作為淡水生態系統中重要的雜食性消費者[1–2,11–12], 以底棲無脊椎動物、碎屑、水生植物和藻類為食, 并且可以通過挖掘和直接切割成植物碎片來影響水生植物的生長和群落演替[41–42]。Cronin等[23]研究發現, 克氏原螯蝦更喜食絲狀或細小分枝、易碎的水生植物, 而本研究中, 輪葉黑藻相對于另外兩種沉水植物來說, 其具有細小分枝、易碎等特性, 克氏原螯蝦對其的鮮重和干重的取食量都是最高的, 結果表明, 水生植物的物理結構對克氏原螯蝦的牧食具有一定的影響。
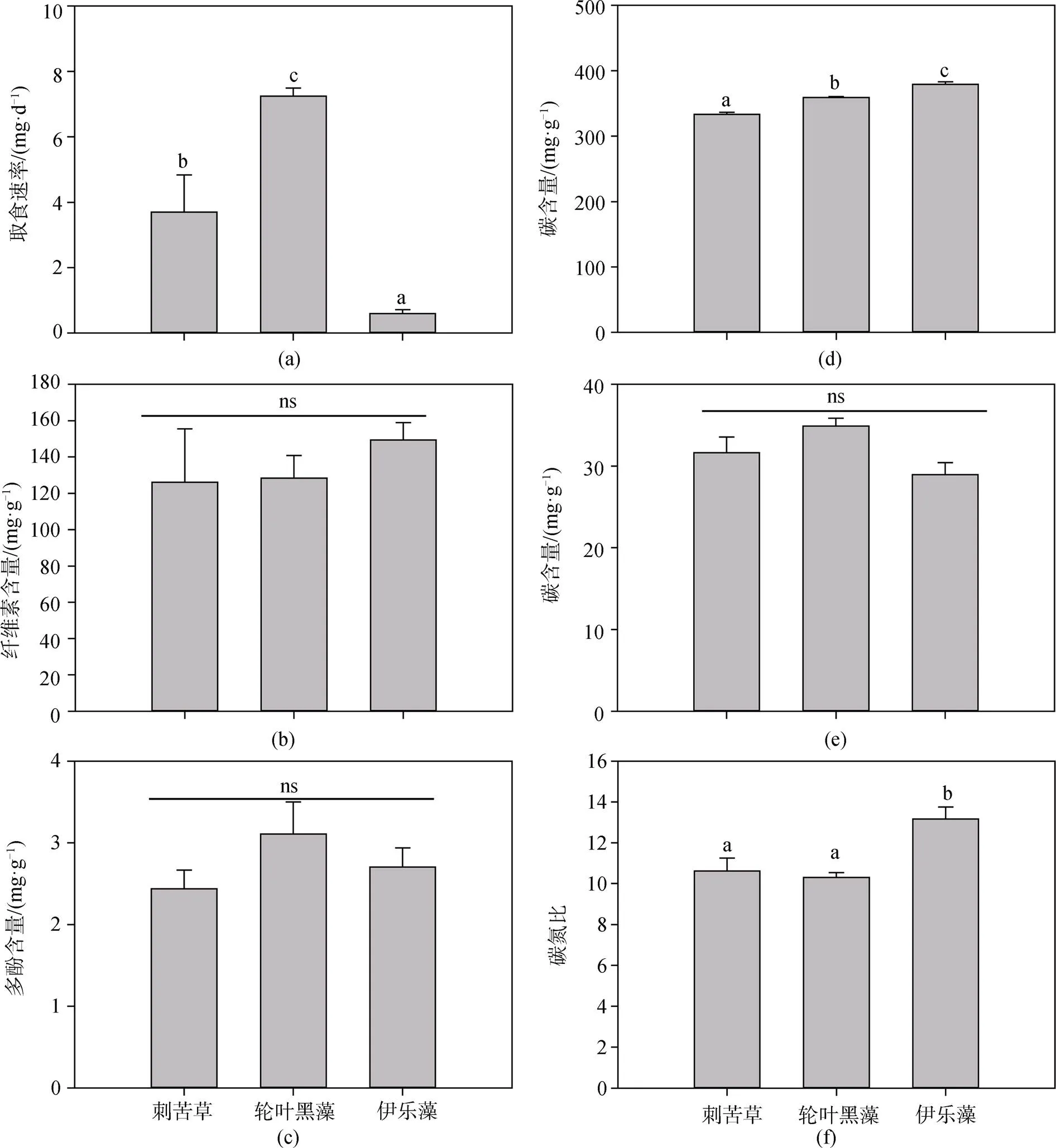
圖1 克氏原螯蝦對3種沉水植物的取食速率及3種沉水植物纖維素、多酚、碳、氮含量和碳氮比
Figure 1 The daily consumed biomass by herbivory of, and the cellulose content, polyphenol content, C content, N content and C:N ratios of three submerged plants
Carreira等[31]研究5種水生植物的適口性時發現, 克氏原螯蝦每天可以消耗單位體重0.6%—2.4%的水生植物干物質量, 取食速率為42.6—114.9 mg·d-1; 而Cronin等[23]研究發現克氏原螯蝦對水生植物的平均鮮重取食量大約為440 mg·d-1, 而本研究中克氏原螯蝦對輪葉黑藻、苦草、伊樂藻的取食速率分別為(7.24±0.24) mg·d-1、(3.70±1.14) mg·d-1和(0.60±0.12) mg·d-1, 鮮重取食速率約為232.66 mg·d-1, 取食速率相對偏低, 可能與研究的物種個體差異有關, 在Carreira等[31]和Cronin等[23]的研究中都以挺水植物為主, 而挺水植物相對沉水植物來說含有更多的碳和更低的氮[43], 克氏原螯蝦為了滿足自身的生長代謝需求, 需要攝食更多的水生植物。
3.2 纖維素和多酚含量對3種沉水植物適口性的影響
植物適口性決定了草食性動物對植物的牧食選擇性, 也決定了植物可能承受的牧食壓力[28], 纖維素作為植物細胞壁的主要組成成分, 在一定程度上增加了植物葉片的物理防御能力, 降低了植物的適口性[21], 同一種植物在不同生長發育期或不同植物在同一生長發育期, 其纖維素含量也會有所差異[44]。但本研究中, 3種沉水植物的纖維素含量卻沒有顯著性差異(圖1b), 其中, 輪葉黑藻、刺苦草、伊樂藻的纖維素含量分別為(128.32±12.49) mg·g-1、(126.02±29.58) mg·g-1和(149.30±9.63) mg·g-1, 結果表明這3種沉水植物的適口性的差異可能與其它結構性物質或防御物質有關, 如化學防御物質[44]。
多酚作為植物體內重要的化學防御物質不僅可以降低植物的適口性, 在一定程度上反應沉水植物對環境的適應能力[28], 還可以影響牧食者的生長發育[29–30]。但也有研究表明, 酚類的含量不僅受到牧食的影響, 還受到植物生活型、光照、營養等環境因素的影響, 同時酚類含量的多少并不會影響植物的適口性[19,22,45–46]。而Bolser等[47]研究發現昆蟲牧食過的植物, 對克氏原螯蝦來說, 適口性更低, 但卻不是由于多酚和蛋白質的差異造成的, 而可能是一些其它的次生代謝產物或營養物質的差異造成的[30]。本研究中, 3種沉水植物的多酚含量并沒有顯著差異(圖1c), 這與Bolser等[47]人的研究結果一致, 說明這3種沉水植物的適口性差異很可能時由于植物的其它次生代謝產物或營養物質含量的差異造成的。
3.3 C、N和C:N對3種沉水植物適口性的影響
植物適口性與植物體內的營養物質含量密切相關, Bryant等[48]提出了植物防衛物質類型(“量”型和“質”型)不同很可能是由于植物體內C、N元素的不平衡所引起的, 在高養分或碳受限的條件下, 碳基防御物質會減少, 而氮基防御物質就會相應增加。Bakker等[25]關于3種不同生境(陸生、淡水、海洋)水生植物碳氮含量及其比值對草食性動物取食速率的研究表明, 水生植物氮(N)含量高、碳氮比(C:N)較低, 對牧食者來說適口性就更高, 反之則適口性更低。本研究中, 3種沉水植物的氮(N)含量沒有顯著性差異, 但伊樂藻的碳(C)含量和碳氮比(C:N)顯著高于刺苦草和輪葉黑藻(圖1d,f), 說明碳含量在克氏原螯蝦的牧食選擇性上具有重要的作用。植物體內的碳主要用于合成含碳化合物, 主要用于增加細胞壁的厚度或韌性, 增加牧食者的消化難度[49], 而本研究中, 富碳化合物—纖維素的含量沒有顯著性差異, 說明這3種沉水植物的適口性可能與其它富碳化合物有關, 如木質素等。
4 結論
(1)對水鱉科3種沉水植物刺苦草、輪葉黑藻和伊樂藻來說, 克氏原螯蝦更喜食輪葉黑藻, 最不喜食伊樂藻, 伊樂藻的適口性最差。
(2)輪葉黑藻的適口性最好, 伊樂藻的適口性最低, 初步推斷, 可能是由于水生植物的物理結構差異和富碳化合物含量較高造成的。但由于本實驗周期較短, 影響牧食者的選擇性是否還存在其他因素, 還需要進一步深入研究。
[1] Hobbs III H H, Jass J P, Huner J V. A review of global crayfish introductions with particular emphasis on two North American species (Decapoda, Cambaridae)[J]. Crus-taceana, 1989, 56(3): 299–316.
[2] Loureiro T G, Anastácio P M S G, Araujo P B, et al. Red Swamp crayfish: biology, ecology and invasion-an overview[J]. Nauplius, 2015, 23(1): 1–19.
[3] Francesca G. Crayfish invading Europe: the case study of[J]. Marine and Freshwater Behaviour and Physiology, 2006, 39(3): 175–191.
[4] 王衛民. 軟殼克氏原螯蝦在我國開發利用的前景[J]. 水生生物學報, 1999, 23(4): 375–381.
[5] 李振宇, 解焱. 中國外來入侵種[M]. 北京: 中國林業出版社, 2002.
[6] 王亞民, 曹文宣. 中國水生外來入侵物種對策研究[J]. 農業環境科學學報, 2006, 25(1): 13–19.
[7] 曹玲亮, 周立志, 張保衛. 安徽三大水系入侵物種克氏原螯蝦的種群遺傳格局[J]. 生物多樣性, 2010, 18(4): 398–407.
[8] 王長忠, 李忠, 梁宏偉, 等. 長江下游地區4個克氏原螯蝦群體的遺傳多樣性分析[J]. 生物多樣性, 2009, 17(5): 518–523.
[9] 張萌, 白俊, 金輝, 等. 不同地理群體的克氏原螯蝦形態差異多元分析[J]. 南昌大學學報(理科版), 2016, 40(2): 188–196.
[10] 蔡鳳金, 武正軍, 何南, 等. 克氏原螯蝦的入侵生態學研究進展[J]. 生態學雜志, 2010, 29(1): 124–132.
[11] Holdich D M, Lowery R S. Freshwater crayfish: biology, management, and exploitation[M]. London: Croom Helm, 1988.
[12] Twardochleb L A, Olden J D, Larson E R. A global meta-analysis of the ecological impacts of nonnative crayfish[J]. Freshwater Science, 2013, 32(4): 1367–1382.
[13] Correia A M, Anastácio P M. Shifts in aquatic macroinvertebrate biodiversity associated with the presence and size of an alien crayfish[J]. Ecological Research, 2008, 23(4): 729–734.
[14] Gamradt S C, Kats L B, Anzalone C B. Aggression by non-native crayfish deters breeding in California Newts[J]. Conservation Biology, 1997, 11(3): 793–796.
[15] Reynolds J D, Soutygrosset C, Kozak P, et al. A review of ecological interactions between crayfish and fish, indigenous and introduced[J]. Knowledge and Management of Aquatic Ecosystems, 2011, 401(10): 1–21.
[16] Matsuzaki S I S, Usio N, Takamura N, et al. Contrasting impacts of invasive engineers on freshwater ecosystems: an experiment and meta-analysis[J]. Oecologia, 2009, 158(4): 673–686.
[17] Van der Wal J M, Dorenbosch M, Immers A K, et al. Invasive crayfish threaten the development of submerged macrophytes in lake restoration[J]. PLoS ONE, 2013, 8(10): e78579.
[18] Fraser L H, Grime J P. Interacting effects of herbivory and fertility on a synthesized plant community[J]. Journal of Ecology, 1999, 87(3): 514–525.
[19] Cronin G. Influence of macrophyte structure, nutritive value, and chemistry on the feeding choices of a generalist crayfish[M]. New York: Springer, 1998.
[20] Mith?fer A, Boland W. Plant defense against herbivores: chemical aspects[J]. Annual Review of Plant Biology, 2012, 63(1): 431–450.
[21] Turley N E, Godfrey R M, Johnson M T. Evolution of mixed strategies of plant defense against herbivores[J]. New Phytologist, 2013, 197(2): 359–361.
[22] Lodge D M. Herbivory on freshwater macrophytes[J]. Aquatic Botany, 1991, 41(1/3): 195–224.
[23] Cronin G, Lodge D M, Hay M E, et al. Crayfish feeding preferences for freshwater macrophytes: the influence of plant structure and chemistry[J]. Journal of Crustacean Biology, 2002, 22(4): 708–718.
[24] Cebrian J, Shurin J B, Borer E T, et al. Producer nutritional quality controls ecosystem trophic structure[J]. PLoS ONE, 2009, 4(3): e4929.
[25] Bakker E S, Wood K A, Pagès J F, et al. Herbivory on freshwater and marine macrophytes: a review and perspective[J]. Aquatic Botany, 2016, 135: 18–36.
[26] Wardle D A, Bonner K I, Barker G M. Linkages between plant litter decomposition, litter quality, and vegetation responses to herbivores[J]. Function Ecology, 2010, 16(5): 585–595.
[27] 朱清順, 周剛, 于能, 等. 以滆湖為例湖泊種植飼料水草的養魚效果研究[J]. 水產學報, 1993, 17(3): 189–197.
[28] Smolders A J P, Vergeer L H T, Velde G V D, et al. Phenolic contents of submerged, emergent and floating leaves of aquatic and semi-aquatic macrophyte species: why do they differ[J]? Oikos, 2000, 91(2): 307–310.
[29] Choi C, Bareiss C, Walenciak O, Gross E M. Impact of polyphenols on growth of the aquatic herbivore[J]. Journal of Chemical Ecology, 2002, 28(11): 2245–2256.
[30] Gross E M, Meyer H, Schilling G. Release and ecological impact of algicidal hydrolysable polyphenols in[J]. Phytochemistry, 1996, 41(1): 133–138.
[31] Carreira B M, Dias M P, Rebelo R. How consumption and fragmentation of macrophytes by the invasive crayfishshape the macrophyte communities of temporary ponds[J]. Hydrobiologia, 2014, 721(1): 89–98.
[32] Gherardi F, Aquiloni L, Diéguezuribeondo J, et al. Managing invasive crayfish: is there a hope[J]? Aquatic Sciences, 2011, 73(2): 185–200.
[33] Aquiloni L, Gherardi F. The use of sex pheromones for the control of invasive populations of the crayfish: a field study[J]. Hydrobiologia, 2010, 649(1): 249–254.
[34] 肖鳴鶴, 肖英平, 吳志強, 等. 養殖密度對克氏原螯蝦幼蝦生長、消化酶活力和生理生化指標的影響[J]. 水產學報, 2012, 36(7): 1088–1093.
[35] 劉琦. 克氏原螯蝦對兩種環境異質性的響應及對隱蔽所的選擇[D]. 南京: 南京大學, 2017.
[36] 田娟, 許巧情, 田羅, 等. 洞庭湖克氏原螯蝦肌肉成分分析及品質特性分析[J]. 水生生物學報, 2017, 41(4): 870–877.
[37] 武正軍, 蔡鳳金, 賈運鋒, 等. 桂林地區克氏原螯蝦對澤蛙蝌蚪的捕食[J]. 生物多樣性, 2008, 16(2): 150–155.
[38] 徐增洪, 周鑫, 水燕. 克氏原螯蝦的食物選擇性及其攝食節律[J]. 大連海洋大學學報, 2012, 27(2): 166–170.
[39] Ragan M A, Craigie J S. Phenolic compounds in brown and red algae[M]. London: Cambrige University Press, 1978.
[40] Goering H K, Van Soest P J. Forage fiber analysis: apparatus reagents, procedures and some applications. Washington D.C.: USDA, 1970.
[41] Nystr?m P, Br?nmark C, Granéli W. Patterns in benthic food webs: a role for omnivorous crayfish[J]? Freshwater Biology, 1996, 36: 631–646.
[42] Nystr?m P, Strand J. A. Grazing by a native and an exotic crayfish on aquatic macrophytes[J]. Freshwater Biology, 2010, 36(3): 673–682.
[43] Xia Chengxing, Yu Dan, Wang Zhong, et al. Stoichiometry patterns of leaf carbon, nitrogen and phosphorous in aquatic macrophytes in eastern China[J]. Ecological Engineering, 2014, 70(5): 406–413.
[44] 郭新紅, 喻達時, 王婕, 等.6種植物中木質纖維素含量的比較研究[J].湖南大學學報(自然科學版), 2008, 35(9): 76–78.
[45] 程春龍, 李俊清. 植物多酚的定量分析方法和生態作用研究進展[J]. 應用生態學報, 2006, 17(12): 2457–2460.
[46] 袁昌波, 曹特, 周存宇, 等. 洱海三種生活型水生植物葉片中碳、氮與總酚含量的比較研究[J]. 水生生物學報, 2016, 40(5): 1025–1032.
[47] Bolser R C, Hay M E, Lindquist N, et al. Chemical defenses of freshwater macrophytes against crayfish herbivory[J]. Journal of Chemical Ecology, 1998, 24(10): 1639–1658.
[48] Bryant J P, Chapin F S, Klein D R. Carbon/nutrient balance of boreal plants in relation to vertebrate herbivory[J]. Oikos, 1983, 40(3): 357.
[49] Gruner D S, Smith J E, Seabloom E W,et al. A cross-system synthesis of consumer and nutrient resource control on producer biomass[J]. Ecology letters, 2008, 11(7): 740–755.
Herbivory of the invasive crayfishon three submerged macrophytes
YUAN Ye1, ZHANG Yinzhe1, LV Chaochao1, MA Xufa1,2,3, XIA Chengxing1,2,3,*
1. Key Lab of Freshwater Animal Breeding, Ministry of Agriculture, College of Fisheries, Huazhong Agricultural University, Wuhan 430070, China 2. Freshwater Aquaculture Collaborative Innovation Center of Hubei Province, Wuhan 430070, China 3. Hubei Provincial Engineering Laboratory for Pond Aquaculture, Wuhan 430070, China
This study investigated the palatability of three species (,and) by feeding testto figure out the feeding preferences and the grazing intensity of submerged macrophytes for invasive crayfish. Our results showed that the daily consumed biomass of crayfish onwas the largest (7.24±0.24) mg·d-1, followed by(3.70±1.14) mg·d-1, and(0.60±0.12) mg·d-1, indicating that the palatability ofwas the worst,was the best, andwas in the middle. Our findings suggested that the cellulose content, polyphenol content, and nitrogen content (N) of the three submerged macrophytes were not significant difference, buthad more C and higher C:N ratio. In a word, the content of physical characteristics, C and C:N ratio of the three submerged macropyhtes played an important role on the herbivory of crayfish on submerged macrophytes.
; submerged macrophytes; herbivory; palatability; C:N ratio
10.14108/j.cnki.1008-8873.2019.05.004
Q948.8
A
1008-8873(2019)05-023-07
2018-10-19;
2018-11-14
國家自然科學基金青年科學基金項目(31500293); 中央高校基本科研業務費專項(2662015BQ038)
袁野(1990—), 男, 湖北十堰人, 碩士, 主要從事水生植物生理生態學研究, E-mail: 200831315@qq.com
夏成星, 男, 博士, 講師, 主要從事淡水生態學研究, E-mail: xiachengxing@mail.hzau.edu.cn
袁野, 張尹哲, 呂超超, 等. 外來種克氏原螯蝦()對3種沉水植物的牧食研究[J]. 生態科學, 2019, 38(5): 23-29.
YUAN Ye, ZHANG Yinzhe, LV Chaochao, et al. Herbivory of the invasive crayfishon three submerged macrophytes[J]. Ecological Science, 2019, 38(5): 23-29.
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·兒童版(2015年1期)2015-07-07 04:12:52
