小麥對鎘脅迫的生理響應及體內鎘的積累分布
2019-10-10 08:20:26于永昂王潤豪胡海燕李成偉
貴州農業科學 2019年9期
關鍵詞:植物
于永昂, 王潤豪, 胡海燕, 李成偉
(河南科技學院 生命科技學院, 河南省糧食作物基因組編輯工程技術研究中心, 河南 新鄉 453003)
近年來,由于工業“三廢”的不合理排放以及農藥化肥不合理的使用,導致土壤中的重金屬鎘含量急劇增加[1-3]。其中,土壤鎘污染超標率居首位,超標率達7.0%[4]。鎘(Cadmium,Cd)是植物生長發育的非必需元素,其毒性強、污染范圍廣、遷移性強,極易被植物吸收積累,不僅嚴重影響作物生長發育,降低其產量和品質,而且能通過食物鏈富集,危害人體健康,會產生致畸、致癌及致突變等效應[5-7]。隨著工業進程的快速發展,鎘超標農產品報道日益增多,鎘進入生態系統危害人類健康的現象日益增多,嚴重影響農產品的食用安全和人類健康[8]。鎘對土壤的污染已成為人們所面臨且亟待解決的環境污染問題之一。
植物種子在萌發期對外界環境較為敏感,鎘對植物生長發育具有明顯的毒害作用,對種子的萌發產生不同程度的刺激和抑制作用,影響植株的生長發育進程,導致植株生長遲緩、植株矮小、葉片泛黃等癥狀[9]。 鎘在植物中累積到一定程度后會阻礙光合作用和呼吸作用[10-11],降低蒸騰速率,引起細胞結構的多種損傷,改變細胞膜的透性,破壞細胞內酶及代謝作用[12],導致養分虧缺,生理生化過程紊亂,最終使植物的生長發育受到不同程度的抑制,嚴重時可導致死亡[13]。因此,植物通過抗氧化防御機制抵抗鎘的毒性[14],抗氧化酶系統可有效地預防或降低植物過氧化作用帶來的損傷并抑制 MDA的積累[15]。重金屬被植物吸收以后,通過調節生理和生物化學新陳代謝過程緩解或者去除鎘的毒害,主要通過對重金屬的吸收運輸、各部位的分配方式以及與植物體內物質的結合形態等因素克服重金屬脅迫危害[16]。如通過植物細胞壁固定或區隔在液泡內,根際 pH的改變,或通過氨基酸、蛋白質及多肽與 鎘結合,積累、儲存并固定等方式。研究表明, 鎘的亞細胞分布與植物的金屬耐性及解毒機制有關,亞細胞鎘主要分布在根系細胞壁內[17]。因此,鎘在植物中的器官積累及亞細胞分布規律是影響植物體內鎘遷移、積累特性和植物毒性的重要因素。
小麥是世界上最重要的糧食作物之一,同樣也是中國第二大糧食作物,其在全世界的糧食安全中占據重要地位[18]。小麥作為鎘積累型作物,對鎘具有較強的耐性,具有潛在的鎘污染風險。目前,鎘的植物毒性機制研究主要集中在光合特性、根系特性、種子萌發、植株酶活性及植物生理生化影響等方面[19-20],關于小麥器官及亞細胞不同組分鎘積累分配差異,小麥解毒及耐性機制尚缺乏較為深入的研究。因此,研究小麥在鎘脅迫下的植物毒性機制及應答機制,對于提高小麥對鎘的抗/耐性,改良糧食作物品質,降低鎘從植物進入食物鏈的危險性具有十分重要的理論和實踐指導意義。為此,采用水培法研究不同濃度鎘脅迫下,小麥種子的發芽率變化,在小麥幼苗器官中鎘的分布規律及其對鎘的吸收和積累差異,以期為鎘在小麥體內的遷移機理以及小麥耐鎘機制的研究提供理論依據。
1材料與方法
1.1供試材料
小麥:品種為百農207,由河南科技學院小麥遺傳改良中心提供。
1.2試驗方法
1.2.1材料處理試驗設4個鎘濃度處理,分別為0 mmol/L(蒸餾水,對照)、0.5 mmol/L、1.0 mmol/L和2.0 mmol/L。選取飽滿均勻一致的小麥種子,用1%的 NaClO消毒15 min后,用去離子水沖洗干凈,播于鋪有雙層直徑 15 cm無菌濾紙的培養皿中,每皿擺放種子50粒。每日加 10 mL不同濃度鎘溶液,于25℃培養箱內發芽,培養箱光周期為光照16 h/黑暗8 h,相對濕度(75±5)%,每個處理3次重復,7 d后開始測定發芽率、幼苗生長狀況和生理生化指標。
發芽勢= 4 d內供試種子發芽數/供試種子數×100%
發芽率= 7 d內供試種子發芽數/供試種子數×100%
1.2.2樣品的處理與測定
1) 小麥測定樣品的處理。選取5株長勢一致的小麥,用去離子水沖洗干凈,再用20 mmol/LNa2-EDTA浸泡30 min,去除根系吸附的鎘,自來水沖洗 3~4次,最后用去離子水潤洗,吸干表面水分,分成根系和地上部2個部分。根系和地上部再分成2部分,一部分在105℃殺青20 min,于 80℃烘干至恒重,稱量地上部和根干重,粉碎,用于鎘含量測定;另一部分保存于-20℃冰箱內備用,分別用于生理生化指標和植株亞細胞組分鎘含量的測定。
2) 小麥幼苗生理生化指標的測定。各處理結束后,用丙酮提取法測定小麥葉片的葉綠素(CHL)含量;用硫代巴比妥酸法測定丙二醛(MDA)含量;超氧化物歧化酶活性(SOD)采用NBT光還原法測定;過氧化物酶(POD)活性采用愈創木酚法測定;過氧化氫酶(CAT)活性采用紫外吸收法測定。
3) 小麥幼苗亞細胞內鎘含量的測定。采用WANG等[21]的方法分離根系和地上部亞細胞組分。取出預處理好的冰凍植物樣,稱取2 g剪碎,用植物樣品與提取液[0.25 mol/L蔗糖+1 mmol/L二硫赤蘚糖醇+50 mmol/L Tris-HCl緩沖液(pH 7.5)]按1∶10的比例混合后迅速將樣品研磨成勻漿。研磨時先放入少量提取液,將大塊的植物樣品盡量磨細后,再加入大量的提取液磨成勻漿,然后轉移到 50 mL離心管中,3 000 r/min離心30 s,下部沉淀和底層碎片為細胞壁組分(F1),得到的上清液在 10 000 r/min離心分離 30 min,下部沉淀為細胞器組分(F2),上清液為可溶組分(F3),全部操作在4℃下進行。
4) 樣品中鎘含量測定。稱取烘干并粉碎后的樣品0.2 g和亞細胞分離出的組分用1∶4(V/V)的HNO3-HClO4混合消煮,定容、過濾,最后采用ICP-OES測定其鎘含量。亞細胞分離出的各組分無需消化,直接用原子吸收光譜儀測定鎘含量。
1.3數據分析
所有數據采用Excel和SAS 8.0進行統計分析。
2結果與分析
2.1鎘脅迫對小麥種子萌發的影響
小麥種子在鎘脅迫條件下能否正常萌發是其能否在鎘污染土壤中生長的先決條件。用于反映種子萌發的重要參數是發芽勢和發芽率,發芽勢反映種子的發芽速度和整齊度,發芽率反映種子發芽多少,是種子質量的綜合反映。由圖1可知,隨著鎘濃度的升高,小麥種子的發芽勢和發芽率均逐漸下降。所有鎘處理小麥種子的發芽率和發芽勢均顯著低于對照。當鎘脅迫濃度為2.0 mmol/L時,其發芽勢和發芽率最低,分別為45%和53%。
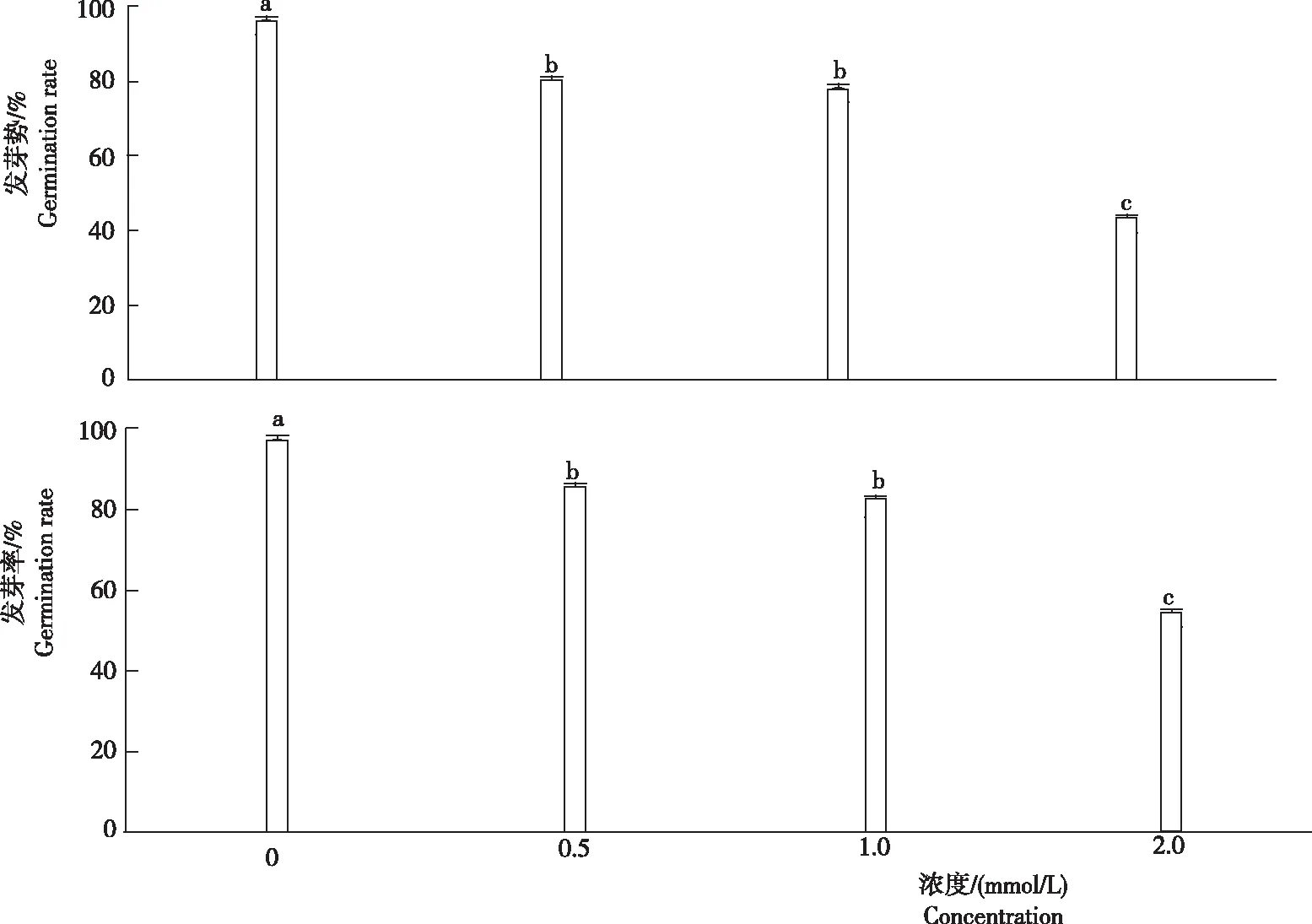
注:不同小寫字母表示處理間差異顯著(P<0.05)水平,下同。
Note:Different letters indicate significant difference atP<0.05 level,the same below.
圖1不同濃度鎘脅迫下小麥種子的發芽勢和發芽率
Fig.1 Germination potential and germination rate of wheat seeds under cadmium stress with different concentration
2.2鎘脅迫對小麥葉綠素及丙二醛含量的影響
2.2.1葉綠素葉綠素是植物進行光合作用的重要色素,其含量是判斷植物光合效率及其同化能力的重要指標之一。由圖2可知,隨著鎘濃度的增加,小麥幼苗葉片葉綠素含量逐漸降低,且各處理均顯著低于對照,鎘濃度0.5 mmol/L、1.0 mmol/L和2.0 mmol/L處理分別較對照降低41.82%、47.27%和76.36%。表明,隨著鎘濃度增加,小麥幼苗的光合作用受到抑制,進而影響幼苗的生長和發育。
2.2.2丙二醛(MDA)丙二醛含量是植物組織在逆境下遭受氧化脅迫發生膜脂過氧化的產物,其含量高低反映膜受損程度。從圖2可知,隨著鎘濃度的增加,小麥幼苗葉片中MDA含量逐漸上升。鎘濃度0.5 mmol/L、1.0 mmol/L和2.0 mmol/L處理的MDA含量較對照顯著增加,分別增加213.65%、617.33%和690.67%。表明,隨著鎘濃度的增加,引起小麥葉片內膜脂過氧化,從而引起膜的嚴重損傷。
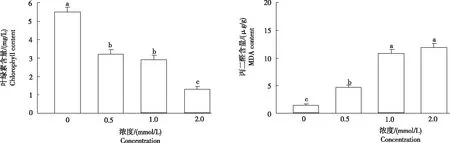
圖2 不同濃度鎘脅迫小麥幼苗葉片的葉綠素及丙二醛含量
2.3鎘脅迫對小麥抗氧化酶活性的影響
植物體內抗氧化保護酶活性的高低是反映植物對逆境適應的重要指標。從圖3可知,隨著鎘脅迫濃度的增大,小麥幼苗葉片的SOD、POD和CAT活性均呈先升后降趨勢。在鎘濃度為0.5 mmol/L時SOD活性最大,與對照相比增加60.71%,且差異顯著。當鎘濃度為2.0 mmol/L時,SOD活性最低,比對照降低9.52%,但與對照無顯著差異,其原因可能是高濃度鎘直接抑制SOD活性,或者對SOD酶基因的轉錄、翻譯過程產生抑制。CAT和POD活性在鎘脅迫下的變化規律與SOD類似,均在鎘濃度為0.5 mmol/L時達到最大,分別比對照增加154.05%和183.33%,且均與對照差異顯著;鎘濃度為2.0 mmol/L時,與對照相當。
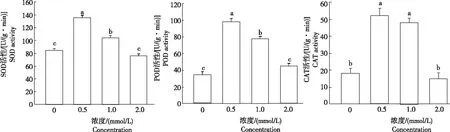
圖3 不同濃度鎘脅迫小麥的抗氧化酶活性
2.4鎘脅迫對小麥幼苗根系及地上部鎘含量的影響
從表1可知,隨著鎘處理濃度的提高,小麥根系和地上部的鎘含量顯著增加;在同一鎘濃度下,小麥根系的鎘含量顯著高于地上部分。其中,鎘濃度0.5 mmol/L、1.0 mmol/L和2.0 mmol/L處理小麥根系的鎘含量分別是地上部的16.38倍、12.80倍和8.7倍。
表1不同鎘濃度處理小麥根系和地上部的鎘含量
Table 1 Cadmium content in aboveground part and root of wheat treated with different cadmium concentrations mg/kg

鎘濃度/(mmol/L)cadmium concentration根系Root地上部Aboveground part0.52 998.86±92.21 c183.05±2.24 c1.04 076.46±103.52 b318.59±2.27 b2.04 705.93±13.32 a540.61±2.19 a
注:同列不同小寫字母表示差異顯著(P<0.05),下同。
Note:Different lowercase letters in the same column indicate significant difference atP<0.05 level.The same below.
2.5鎘在小麥幼苗亞細胞組分中的分布
從表2可知,隨著鎘濃度的升高,小麥根系和地上的細胞壁組分和細胞器組分的鎘含量均顯著上升,且差異顯著;可溶組分均呈先升后降的變化趨勢,且差異顯著。不同濃度鎘處理下,各亞細胞組分吸附鎘的比例均呈細胞器>細胞壁>可溶組分;鎘濃度為0.5 mmol/L時對小麥已產生了嚴重傷害。根系累積的速度遠遠高于地上部的速度,同一鎘濃度處理時,地上部亞細胞中的鎘含量遠低于根系。不同濃度鎘處理后,鎘在根系細胞器中的分布比例為78.07%~90.15%,在整個細胞亞細胞組分中占絕對優勢;在細胞壁中的分布比例為9.81%~21.50%。鎘處理后小麥地上部細胞器、細胞壁和可溶組分的鎘分配比例分別為50.97%~62.71%、36.93%~48.76%和0.27%~0.87%。鎘脅迫下可溶組分的鎘含量在根系和地上部的比例含量均低于1%。
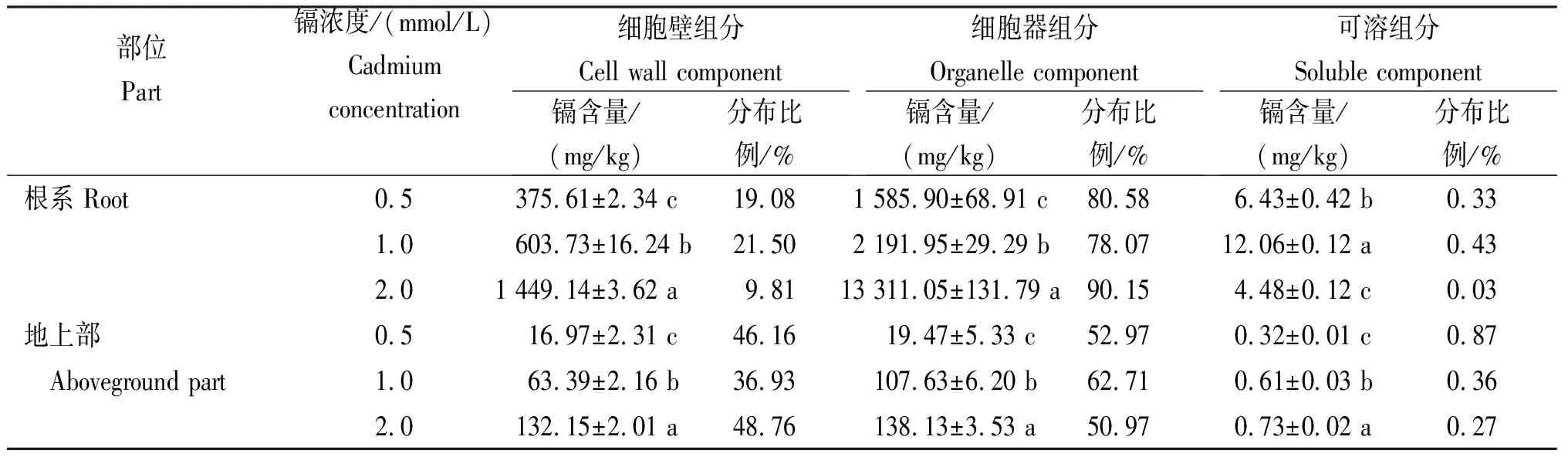
表2 不同濃度鎘處理在小麥亞細胞的分布
3結論與討論
3.1結論
在鎘脅迫下,小麥種子發芽率和發芽勢明顯降低,小麥幼苗的葉綠素含量隨著鎘脅迫濃度的增加逐漸降低,SOD、POD和CAT等抗氧化酶活性均隨著鎘濃度的增加呈先升后降趨勢,均在鎘濃度0.5 mmol/L時達最大。在同一鎘濃度下,小麥根系鎘含量顯著高于地上部;地上部亞細胞中的鎘含量遠低于根系。小麥地上部和根系大部分的鎘分布在細胞壁和細胞器中,而在可溶組分中的分布較少。
3.2討論
種子萌發是作物生長的關鍵時期和對外界脅迫最敏感的階段,種子發芽質量好壞與農作物生長和經濟效益之間有著直接的關系,通過觀察種子發芽對外源鎘的響應,可以更加深入了解鎘脅迫對植物的毒害作用[22]。鎘是重金屬元素,是植物生長過程的非必需元素,但鎘脅迫會對種子萌發和幼苗生長產生不同程度的影響。一般來說,種子的發芽勢高,其種子的生命力就較強,種子出苗整齊一致,而發芽率則能夠反映種子的出苗情況。研究結果表明,鎘脅迫處理顯著降低小麥種子的發芽勢和發芽率,抑制小麥種子的萌發。光合作用是一個高度自我調節的過程,逆境過程可能會導致植株超微結構的改變和功能的破壞,由此光合速率被抑制[23]。研究結果表明,在小麥苗期,隨著鎘濃度升高,小麥幼苗葉綠素含量降低,光合作用受到抑制,阻礙了小麥的生長和發育。
在逆境脅迫下,通常植物會產生高度反應性的活性氧自由基(ROS),而活性氧積累到一定水平便會誘發膜脂不飽和脂肪酸發生連鎖的過氧化反應,使膜結構遭受破壞、細胞功能失常,甚至造成植株死亡。植物可通過抗氧化系統、體內細胞的作用和根部截留等抗性機理來減少重金屬的毒害,而抗氧化系統在植物抵抗逆境時至關重要。研究發現,隨著鎘濃度的增加,MDA含量顯著增加,說明在鎘脅迫下,小麥幼苗中積累了大量的有害物質,對小麥的細胞膜系統造成了嚴重損害。此外,小麥的抗氧化酶系統酶活性(SOD、POD和CAT)均隨鎘脅迫濃度的增加呈先升后降的變化規律,說明小麥具有一定的適應和抵抗鎘傷害的能力,但鎘濃度較高時,小麥幼苗的細胞發生了嚴重損傷,其酶活性出現不可逆下降,與何俊瑜等[24-25]研究結果一致。隨著鎘脅迫濃度的增加,小麥根系和地上部中鎘含量顯著增加,同一鎘濃度處理下,根系中的鎘含量高于地上部,該結果與鎘在植物體內分布的普遍規律即地下部大于地上部結果一致。鎘對植物的危害程度與其在亞細胞中的分布密切相關。研究結果表明,鎘脅迫下,小麥根系和地上部中鎘主要分布在細胞器和細胞壁中,在可溶組分中較少。與大多數研究結果不一致,其原因可能是不同植物有不同的解毒機制,同時也可能與該研究采用的鎘處理濃度有著一定的關系。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
