飼糧中添加甘露寡糖對肉仔雞生長性能及組織天然免疫相關基因表達的影響
2019-10-21 06:19:28熊阿玲包龍飛許蘭嬌龍光蓮李君惠黎觀紅
中國糧油學報 2019年9期
熊阿玲 包龍飛 許蘭嬌 萬 根 龍光蓮 李君惠 黎觀紅
(江西農業大學江西省動物營養重點實驗室;江西省營養飼料開發工程研究中心,南昌 330045)
Toll樣受體(Toll-like receptors, TLRs)是表達于動物免疫細胞和上皮細胞的一類重要模式識別受體(pattern recognition receptors, PRRs),是機體最初感知微生物存在的天然免疫識別受體,是啟動哺乳動物和家禽腸道天然免疫應答的關鍵PRR,而且還作為天然免疫和獲得性免疫的橋梁而調節獲得性免疫應答[1-2]。腸道上皮細胞和免疫細胞可通過 TLRs區分病原微生物和共生有益菌產生不同免疫效應。TLRs能夠識別微生物相關分子模式(microbe-associated molecular patterns, MAMPs)并激活下游信號通路從而釋放出一系列免疫效應分子并產生一系列生理和免疫效應,釋放的免疫效應分子主要包括抗菌肽(β-防御素和cathelicidins)、細胞因子和趨化因子等[3-4]。基于TLRs在天然免疫應答中的重要作用,近年來,雞TLRs(chTLRs, chicken TLRs)的研究受到極大關注。在目前已鑒定的10種禽類chTLRs中,TLR2主要識別來自于G+細菌的細胞壁組分如肽聚糖、脂蛋白和脂磷壁酸,TLR4識別G-細菌的LPS[5]。
抗菌肽(antimicrobial peptides, AMPs)在宿主免疫調控中起到至關重要的作用,是雞抗感染天然免疫至關重要的效應分子[6-8]。β-防御素和cathelicidins是雞體內的兩個主要抗菌肽家族。到目前為止,從雞組織中鑒定出14種禽β-防御素(avian β-defensins, AvBDs),即AvBD1-14[8-9]。在雞組織中發現的cathelicidins(Cath)有4 種,即Cath-1、Cath-2, Cath-3和Cath-B1[10]。雞體內表達的抗菌肽家族中,AvBD9和Cath-B1是廣泛表達于雞組織尤其是消化道和免疫組織器官的一種重要抗菌肽,AvBD9和Cath-B1除具有很強的廣譜抗菌活性外,在體內還發揮著重要的免疫調節功能,在雞的抗感染免疫中發揮重要作用[6-7, 11-12]。
國內外學者對甘露寡糖(mannan-oligosaccharides, MOS)在畜禽生產中的應用進行了大量研究。大多數研究表明,肉雞飼糧中添加MOS可提高日增重、改善飼料利用率、降低死亡率,促進腸道發育,改善腸道微生物區系,提高肉雞腸道免疫和系統免疫功能[13-15]。但MOS調節機體免疫功能的作用機制目前還不是十分清楚。MOS是否可通過影響雞天然免疫相關基因如TLRs和抗菌肽表達而發揮其免疫調節作用?這方面的研究鮮見報道。本研究旨在探討飼糧中添加MOS對肉仔雞生長性能以及重要的天然免疫相關基因TLR2、TLR4、AvBD9和Cath-B1基因表達的影響,從天然免疫角度初步探討MOS的免疫調節作用,為MOS作為抗生素替代品的推廣應用提供參考。
1 材料與方法
1.1 試驗設計
采用單因子試驗設計,選取體重相近的1日齡雄性白羽肉仔雞(科寶500)176只,隨機分成4個處理組,每組4個重復,每個重復11只雞,試驗期為42 d。4個處理組分別飼喂在基礎飼糧中添加0(對照組)、0.3、0.6、0.9 g/kg MOS的飼糧。各組飼糧不添加任何抗生素,本試驗所用的甘露寡糖來源于食品啤酒酵母細胞壁,純度≥95%。
1.2 試驗飼糧組成及營養水平
本試驗采用的基礎飼糧為玉米—豆粕型飼糧,按照1~21、22~42日齡齡兩個階段,根據NRC(1994)和我國《雞的飼養標準》(2004)推薦的營養水平配制。基礎飼糧組成及其營養水平見表1。
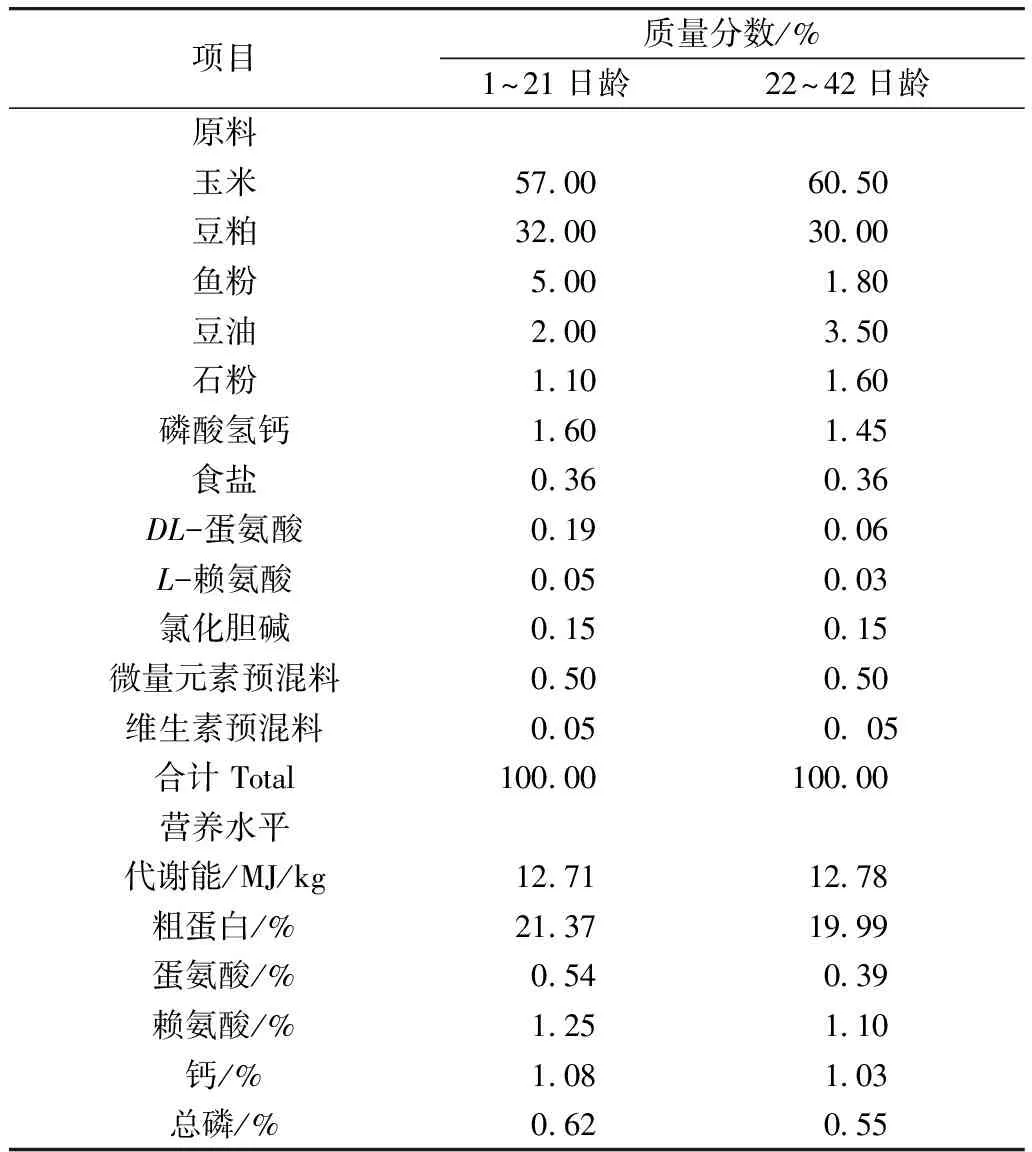
表1 基礎飼糧組成及營養水平
注: 微量元素預混料為每千克飼糧提供:鐵(FeSO4·H2O)80 mg,銅(CuSO4·5H2O)8 mg,鋅(ZnSO4·H2O)60 mg,錳(MnSO4·H2O)80 mg,碘(KI)0.35 mg,硒(Na2SeO3)0.15 mg。維生素預混料為每千克飼糧提供:維生素A 8 000 IU,維生素D33 000 IU,維生素E 10 IU,維生素K32 mg,維生素B11.5 mg,維生素B28.0 mg,維生素B62.5 mg,維生素B120.011 mg,葉酸0.9 mg,煙酸44 mg,D-泛酸鈣11 mg,生物素0.11 mg。營養水平為計算值。
1.3 飼養管理
試驗雞采用三層籠籠養。試驗前對雞舍進行充分沖洗和嚴格消毒,入雛前24 h將雞舍升溫至32~35 ℃,此后溫度每周降低2~3 ℃,直至保持在22~24 ℃為止。采用連續光照、自然通風。試驗期舍內光照、濕度和溫度根據常規飼養管理要求進行控制,雞只按正常免疫程序進行免疫。整個試驗期雞只自由采食和飲水。
1.4 組織樣品的采集與制備
分別在肉仔雞21和42日齡08 ∶00喂料前從每個重復中隨機選取兩只體重相近的雞空腹稱重,稱重后采用頸動脈放血法將肉仔雞致死,冰浴上迅速分離法氏囊、胸腺、脾臟、肝臟、回腸和盲腸,用冰冷的生理鹽水沖洗干凈并用濾紙吸干表面水分,用天平稱重法氏囊、胸腺和脾臟,用于計算器官指數。液氮速凍肝臟、脾臟、回腸和盲腸,轉入-80 ℃低溫冰箱保存備用,用于基因表達量的測定。
1.5 測定指標與方法
1.5.1 生長性能
分別于肉仔雞1、21和42 日齡對每重復肉仔雞進行空腹稱重,并在試驗期間記錄飼料耗料量,計算肉仔雞各階段的平均日增重(ADG)、平均日采食量(ADFI)和料重比(F/G)。
1.5.2 免疫器官指數
根據所屠宰試驗雞體重與免疫器官(法氏囊、胸腺、脾臟)質量計算各免疫器官指數。計算公式:免疫器官指數(g/kg)=免疫器官重(g)/雞活重(kg)。
1.5.3 組織TLR2、TLR4、AvBD9和Cath-B1基因mRNA表達量
采用Trizol法提取肉仔雞肝臟、脾臟、回腸和盲腸組織總RNA,用超微量紫外分光光度儀測定其溶度和純度,用1%瓊脂糖凝膠電泳檢測分析其完整性。對質量合格的RNA進行反轉錄,用于后續試驗。采用反轉錄試劑盒將提取的總RNA反轉錄成cDNA。根據GenBank數據庫中提供的雞GADPH、TLR2、TLR4、AvBD9和Cath-B1基因序列,用Primer Express 3.0軟件設計其引物,引物由ShineGene公司(中國上海)合成,特異性引物及探針的序列見表2。根據試劑盒說明書優化反應體系,對每個樣品 cDNA進行目的基因和內參基因熒光定量PCR反應,擴增反應在FTC2000熒光定量PCR系統(Funglyn Biotech, CA)中進行。反應總體系為50 μL:25 μL 2 × Hotstart Fluo-PCR mix, 2 × 1 μL Primers (25 pmol/μL),0.5 μL Probe(25 pmol/μL),1 μL cDNA,21.5 μL DEPC水。反應條件:94 ℃ 4 min,94 ℃ 20 s,60 ℃ 30 s循環40次。PCR擴增反應結束后,PCR儀自帶的ABI7300 SDS Software分析軟件分析所得結果Ct值。目的基因的mRNA相對表達量用2-△△Ct法分析,內參基因選用GAPDH。

表2 實時定量PCR擴增所使用的特異性引物序列、GenBank登入號及產物大小
1.6 數據處理與統計分析
采用SPSS 17.0軟件的One-way ANOVA過程進行單因素方差分析,采用Duncan氏法進行多重比較。結果以“平均值±標準差”表示,P<0.01表示差異極顯著,P<0.05表示差異顯著,P>0.05表示差異不顯著。
2 結果
2.1 甘露寡糖對肉仔雞生長性能的影響
由表3可知,飼糧中添加MOS對肉仔雞生長前期(1~21日齡)平均日采食量無顯著影響(P>0.05),但顯著提高肉仔雞生長前期平均日增重和顯著降低料重比(P<0.05或P<0.01),MOS不同添加水平組之間平均日增重和料重比無顯著差異(P>0.05)。與對照組相比,飼糧中添加0.6 g/kg MOS顯著提高肉仔雞生長后期(22~42日齡)平均日采食量(P<0.01)、平均日增重(P<0.05)和料重比(P<0.01),MOS不同添加水平組之間日增重無顯著差異。添加0.3~0.9 g/kg MOS顯著提高肉仔雞生長全期(1~42日齡)的平均日采食量和平均日增重(P<0.05或P<0.01)。

表3 飼糧中添加甘露寡糖對肉仔雞生長性能的影響
注:在同一日齡,同列數據肩標字母相同或無肩標字母表示差異不顯著(P>0.05),肩標小寫字母不同表示差異顯著(P<0.05),大寫字母不同表示差異極顯著(P<0.01)。余同。
2.2 甘露寡糖對肉仔雞免疫器官指數的影響
由表4可知,飼糧中添加0.6 g/kg和0.9 g/kg MOS顯著提高21日齡肉仔雞的法氏囊指數(P<0.05),飼糧中添加0.9 g/kg MOS顯著提高42日齡胸腺指數(P<0.05),但MOS不同添加水平組之間差異不顯著(P>0.05)。添加不同水平MOS處理組對21日齡肉仔雞胸腺指數、脾臟指數和42日齡的法氏囊指數、脾臟指數均無顯著影響(P>0.05)。
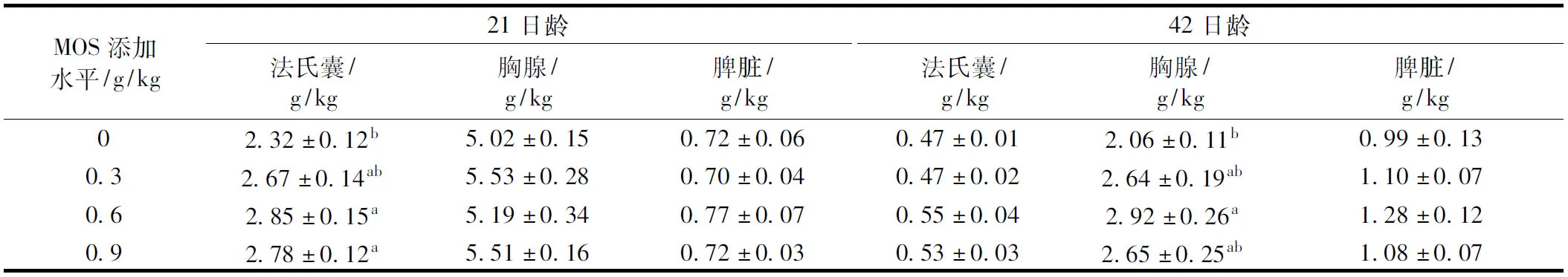
表4 甘露寡糖對肉仔雞免疫器官指數的影響
2.3 甘露寡糖對肉仔雞組織TLR2 mRNA表達的影響
由表5可知,飼糧添加MOS對21日齡肉仔雞肝臟、脾臟、回腸和盲腸以及42日齡肝臟和脾臟TLR2 mRNA表達無顯著影響(P>0.05),但顯著影響42日齡肉仔雞回腸和盲腸TLR2 mRNA表達(P<0.05或P<0.01)。與對照組相比,飼糧添加0.3和0.9 g/kg MOS分別極顯著提高42日齡肉仔雞回腸和盲腸TLR2 mRNA表達(P<0.01)。由表5亦可知,TLR2基因在所測定的肝臟、脾臟、回腸和盲腸四個組織中的表達存在組織差異性,以盲腸TLR2 mRNA表達水平最高。
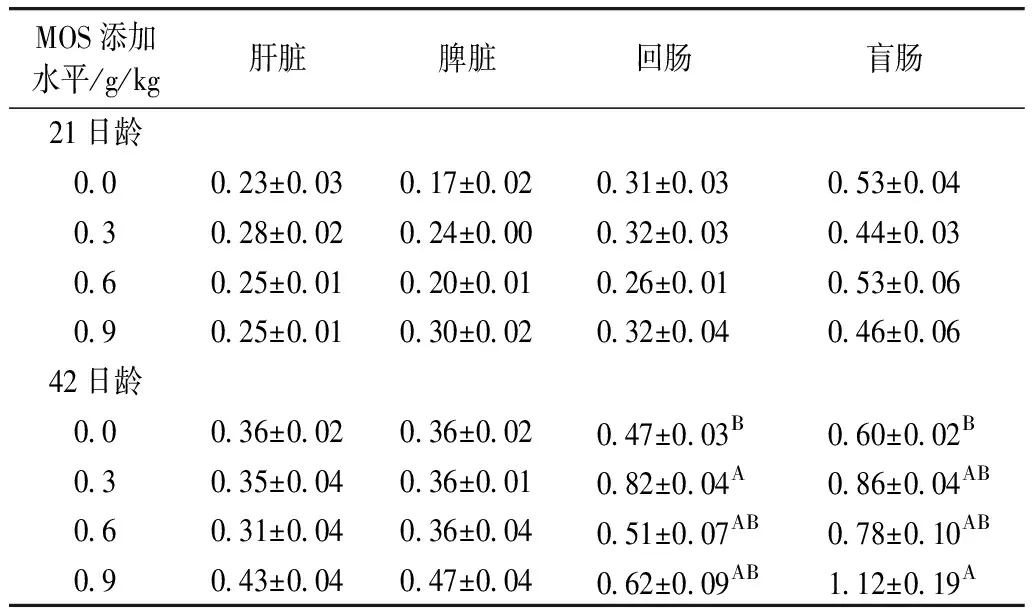
表5 飼糧中添加甘露寡糖對肉仔雞組織TLR2 mRNA表達的影響
2.4 甘露寡糖對肉仔雞組織TLR4 mRNA表達的影響
由表6可知,飼糧中添加MOS對21日齡肉仔雞脾臟、盲腸和42日齡盲腸TLR4 mRNA表達無顯著影響(P>0.05),但顯著影響21日齡肝臟、回腸和42日齡肉雞肝臟、脾臟、回腸TLR4 mRNA表達(P<0.05或P<0.01)。在21日齡,飼糧中添加0.6和0.9 g/kg MOS極顯著提高肝臟TLR4 mRNA表達(P<0.01),但0.3 g/kg MOS添加組與對照組差異不顯著(P>0.05);回腸TLR4 mRNA表達隨MOS添加量的增加而呈上升趨勢,0.9 g/kg MOS添加組顯著高于對照組(P<0.05)。在42日齡,三個MOS添加組均與對照組差異不顯著(P>0.05)。飼糧中添加0.6和0.9 g/kg MOS分別顯著提高42日齡回腸(P<0.01)和脾臟(P<0.05)TLR4 mRNA表達。由表6亦可知,在所測定的肝臟、脾臟、回腸和盲腸的四個組織中,以盲腸TLR4 mRNA表達水平最高,而肝臟表達水平最低。

表6 飼糧中添加甘露寡糖對肉仔雞組織TLR4 mRNA表達的影響
2.5 甘露寡糖對肉仔雞組織AvBD9 mRNA表達的影響
由表7可知,飼糧中添加MOS顯著影響21日齡肉仔雞脾臟、回腸和盲腸以及42日齡回腸和盲腸AvBD9 mRNA表達(P<0.05或P<0.01)。在21日齡,與對照組相比,飼糧中添加0.3 g/kg MOS顯著提高脾臟(P<0.05)和回腸(P<0.01)AvBD9 mRNA的表達;飼糧中添加0.6 g/kg MOS極顯著提高回腸和盲腸AvBD9 mRNA表達(P<0.01),而飼糧中添加0.9 g/kg MOS顯著提高盲腸AvBD9 mRNA表達(P<0.05)。在42日齡,與對照組相比,飼糧中添加0.9 g/kg MOS顯著提高回腸AvBD9 mRNA的表達(P<0.05);飼糧中添加0.3~0.9 g/kg MOS極顯著提高盲腸AvBD9 mRNA表達(P<0.01)。由表7亦

表7 飼糧中添加甘露寡糖對肉仔雞組織AvBD9 mRNA表達的影響
可知,AvBD9 mRNA表達水平在所測定的肝臟、脾臟、回腸和盲腸四個組織中存在差異,肝臟AvBD9 mRNA表達水平最高,而回腸表達水平最低。
2.6 甘露寡糖對肉仔雞組織Cath-B1 mRNA表達的影響
由表8可知,飼糧中添加MOS對21日齡肉仔雞肝臟和42日齡回腸Cath-B1 mRNA表達無顯著影響(P>0.05),但顯著影響21日齡肉仔雞脾臟、回腸和盲腸以及42日齡肝臟、脾臟和盲腸Cath-B1 mRNA表達(P<0.05或P<0.01)。在21日齡,與對照組相比,飼糧中添加0.3 g/kg MOS顯著提高脾臟(P<0.05)以及極顯著提高回腸和盲腸Cath-B1 mRNA表達(P<0.01);添加0.9 g/kgMOS極顯著提高盲腸Cath-B1 mRNA表達(P<0.01),但顯著降低脾臟Cath-B1 mRNA表達(P<0.05)。在42日齡,與對照組相比,飼糧中添加0.6 g/kg MOS極顯著提高肝臟Cath-B1 mRNA表達(P<0.01),而飼糧中添加0.9 g/kg MOS顯著提高肝臟和脾臟(P<0.05)以及極顯著提高盲腸(P<0.01)Cath-B1 mRNA表達。由表8亦可知,Cath-B1 mRNA表達水平在所測定的肝臟、脾臟、回腸和盲腸四個組織中存在差異,Cath-B1 mRNA高表達于脾臟和盲腸,而在肝臟中表達水平最低。

表8 飼糧中添加甘露寡糖對肉仔雞組織Cath-B1 mRNA表達的影響
3 討論
國內外學者對飼糧添加MOS對畜禽生長性能的影響進行了大量的研究,但不同研究者所得結果不盡一致。Pelicano等[16]報道,飼糧添加MOS可顯著提高1~21日齡肉仔雞日增重和飼料轉化率,MOS對肉仔雞生長性能的改善效應并不能持續至42日齡。Hooge 等[17]、Kumprecht等[18]和武威等[19]試驗結果均表明,MOS可顯著提高肉仔雞的生長性能。本研究發現飼糧中添加MOS能顯著改善肉仔雞生長性能,添加0.3~0.9 g/kg MOS顯著提高肉仔雞生長前期(1~21日齡)的平均日增重和降低料重比,且添加0.9 g/kg MOS的可取得最佳的料重比;添加0.6 g/kg MOS對提高肉仔雞生長后期(22~42日齡)平均日采食量和平均日增重的效果最明顯。本試驗結果與上述報道的研究結果類似。然而,Biggs等[20]和Baurhoo等[21]均報道,飼糧添加MOS對生長前期肉雞的生產性能沒有改善作用。Yang等[22]研究報道,MOS對肉雞生長后期(22~42日齡)的生產性能沒有改善作用。相反,Sohail等[23]和Cheng等[24]分別發現,飼糧中添加MOS可顯著改善熱應激條件下肉雞日增重、采食量和飼料轉化率。有關MOS對肉雞生產性能的改善作用,不同學者的研究結果不盡一致,其原因可能與MOS的來源、添加量、飼糧組成、雞的生長階段及飼養環境等因素有關。
禽類的免疫器官主要包括法氏囊、胸腺和脾臟,其中,法氏囊和胸腺為禽類的中樞免疫器官,脾臟為禽類的次級免疫器官。免疫器官指數的大小常用來反映或評價機體免疫功能狀態[25]。本研究發現,飼糧中添加0.6 g/kg可分別提高21日齡肉仔雞法氏囊指數和42日齡胸腺指數,由此說明,飼糧添加適量的MOS可提高肉雞免疫功能。同樣地, Sohail等[26]和Attia等[27]均報道,飼糧添加MOS可顯著提高肉仔雞法氏囊和胸腺指數,但對脾臟指數無影響,這與本研究結果相一致。
TLRs是啟動哺乳動物和家禽天然免疫應答的關鍵模式識別受體,并可調節獲得性免疫應答,在機體的抗感染免疫中發揮重要作用。研究表明,TLR2可識別酵母細胞壁甘露聚糖,介導宿主相關免疫效應分子的分泌和天然免疫應答的產生[28]。TLR4亦可識別葡萄糖醛酸木糖甘露聚糖(glucuronoxylomannan)、釀酒酵母(Saccharomycescerevisiae)和假絲酵母(Candidaalbicans)來源的甘露聚糖[29]。TLR2和TLR4表達的提高及其介導的下游信號通路的激活與胃腸道屏障功能的改善密切相關,由此提高腸道對病原菌入侵和對感染的抵抗力[30-31]。來源于酵母細胞壁MOS作為甘露聚糖大分子的一部分,亦可能作為MAMP而被TLRs識別而啟動機體天然免疫應答。本研究發現,在本試驗添加水平范圍內(0.3~0.9 g/kg),飼糧添加MOS可不同程度地提高肉雞回腸和盲腸中TLR2和TLR4 mRNA的表達。Yitbarek等[32]報道,產氣莢膜梭菌(Clostridiumperfringens)感染對肉雞回腸TLR2和TLR4以及盲腸扁桃體TLR4基因表達無影響,但飼糧中添加MOS顯著提高感染產氣莢膜梭菌肉雞回腸TLR2和TLR4以及盲腸扁桃體TLR4基因表達,但感染產氣莢膜梭菌的肉雞其盲腸扁桃體TLR2基因表達顯著升高,而飼糧中添加MOS對感染產氣莢膜梭菌肉雞盲腸扁桃體TLR2的表達無影響。Cheled-Shoval等[33]研究發現,出殼前3d給肉雞胚胎注射MOS可提高小腸TLR4基因表達,但對小腸TLR2基因表達無影響。本研究結果與Yitbarek等[32]報道的研究結果相一致,但與Cheled-Shoval等[33]報道的TLR2基因表達結果不一致。同樣地,Munyaka等[34]報道,飼糧添加MOS對42日齡肉仔雞回腸和盲腸扁桃體TLR2 mRNA的表達無影響,但顯著降低TLR4 mRNA的表達,而本研究發現飼糧添加MOS對盲腸TLR4 mRNA無顯著影響。功能性寡糖如菊粉和低聚果糖除直接與腸道上皮細胞和腸道免疫細胞上的碳水化合物識別受體如某些TLRs、甘露糖受體、C型凝集素受體等受體結合以啟動免疫應答而提高機體對病原菌的清除能力和保護腸道健康等作用外,部分被吸收的寡糖可引起局部如脾臟、肝臟和胸腺或系統免疫應答反應[35-36]。本研究發現,MOS除不同程度提高腸道TLR2和TLR4表達外,MOS還可不同程度地提高肝臟和脾臟TLR2和TLR4 mRNA的表達。由此表明,MOS除直接在腸道發揮免疫調節作用外,部分MOS被完整吸收后或其被腸道微生物作用后的代謝產物被吸收后調節機體其他組織尤其是免疫器官組織細胞TLRs表達并由此調節機體局部和系統免疫功能,但這需進一步的試驗證明。此外,本研究發現,TLR2和TLR4 mRNA的表達存在組織差異性,在所測的四個組織中,以盲腸TLR2和TLR4 mRNA表達量最高。
動物體內源表達的抗菌肽如β-防御素和cathelicidins是TLRs激活后產生的關鍵免疫效應分子之一[3-4]。研究表明,表達于腸道上皮細胞和免疫細胞的重要模式識別受體TLR2可識別酵母細胞壁甘露聚糖,甘露聚糖與TLR2結合后激活信號通路分子而產生免疫效應分子介導天然免疫和獲得性免疫應答反應。禽AvBD9和是Cath-B1廣泛表達于雞各種組織尤其是消化道和免疫器官的一種重要抗菌肽,具有很強的殺滅致病細菌和真菌活性,同時,在雞的天然免疫和獲得性免疫中亦發揮重要作用[6-9,12]。本研究發現,飼糧中添加適宜水平的MOS可不同程度地提高肉仔雞回腸、盲腸、肝臟和脾臟AvBD9及Cath-B1基因的表達,與相應組織中TLRs尤其是TLR2表達結果呈現相似的變化。綜上結果可知,MOS可能夠通過提高肉雞組織TLRs表達并由TLRs介導上調β-防御素和cathelicidins等抗菌肽表達而提高肉雞天然免疫防御功能。
4 結論
綜合本試驗結果,飼糧中添加適宜水平的MOS可顯著提高肉仔雞生長性能,且不同程度地提高肉雞肝臟、脾臟、回腸和盲腸中TLR2、TLR4、AvBD9和Cath-B1的表達。MOS可能通過調控肉雞組織TLRs表達并由TLRs介導上調β-防御素和cathelicidins等抗菌肽的表達而提高肉雞天然免疫防御功能。綜合生長性能、免疫器官指數和天然免疫相關基因表達指標,日糧添加0.6 g/kg MOS可取得較佳效果。
