稀土尾砂干旱脅迫對2種牧草種子萌發(fā)與幼苗生理特性的影響
2019-10-25 01:27:00張海娜魯向暉王瑞峰
江蘇農(nóng)業(yè)科學(xué) 2019年13期
張海娜 魯向暉 王瑞峰

摘要:稀土礦尾砂保水能力差,干熱化嚴重,常采用冷季草與暖季草混播的方式進行植被恢復(fù)。采用不同濃度梯度(0~30%)聚乙二醇(PEG-6000)溶液模擬稀土尾砂干旱,研究常見冷季草多年生黑麥草(Lolium perenne)和高羊茅(Festuca elata)種子萌發(fā)和幼苗生長的抗旱性,以期為稀土尾砂冷季草種的選擇提供理論依據(jù)。研究結(jié)果表明,隨干旱脅迫加劇,多年生黑麥草和高羊茅種子的發(fā)芽率、發(fā)芽勢、發(fā)芽指數(shù)呈降低趨勢,分別在20%、15%脅迫下顯著降低;幼苗期,多年生黑麥草和高羊茅的葉綠素a、葉綠素b、葉綠素a+b的含量呈現(xiàn)先降低后升高的趨勢;在中度和重度脅迫下,多年生黑麥草的葉綠素含量、丙二醛(MDA)含量、可溶性蛋白含量、游離氨基酸含量顯著高于高羊茅;相關(guān)性分析表明,MDA含量、可溶性蛋白含量和游離氨基酸含量之間存在極顯著正相關(guān)關(guān)系;隸屬函數(shù)評價顯示,多年生黑麥草較高羊茅對稀土尾砂干旱脅迫具有更強的適應(yīng)性。在稀土礦尾砂區(qū)植被恢復(fù)中可優(yōu)先選擇冷季草多年生黑麥草。
關(guān)鍵詞:稀土尾砂;多年生黑麥草;高羊茅;干旱脅迫;滲透調(diào)節(jié)物質(zhì)
中圖分類號:Q945.78;S540.1 ?文獻標志碼: A ?文章編號:1002-1302(2019)13-0204-05
離子型稀土礦是我國重要的戰(zhàn)略資源,南方紅壤區(qū)是主要產(chǎn)區(qū)。長期以來,稀土礦開采尤其早期多采用堆浸工藝,遺留大面積裸露尾砂地,形成類似“沙漠化”的景觀,水土流失嚴重[1-2]。采取有效的礦山植被修復(fù)措施來改善稀土尾砂生態(tài)環(huán)境是迫切需要解決的問題。然而,稀土尾砂地表裸露、干熱化強烈,土壤功能低下[3],尤其季節(jié)性干旱期,植物生長受到嚴重的水分脅迫,甚至威脅到植物幼苗的存活。常見的方法是將冷季草和暖季草混合播種。
多年生黑麥草(Lolium perenne)和高羊茅(Festuca elata)是典型的冷季草種類,分蘗能力強,覆綠速度快,是我國廣大溫帶地區(qū)地表覆綠的重要材料,關(guān)于這2種植物抗旱性的研究較多。在研究PEG模擬干旱脅迫對冷季型草坪草種子萌發(fā)和幼苗生長的影響中發(fā)現(xiàn),高羊茅的抗旱性高于多年生黑麥草[4-5]。也有研究采用盆栽育苗法對冷季型草種生長期的生理特性進行比較,表明高羊茅抗旱性高于多年生黑麥草[6-8];高涵等對幾種冷季草苗期的抗旱性研究顯示,多年生黑麥草高于高羊茅[9]。可見,多年生黑麥草和高羊茅抗旱性的研究多集中在草坪成坪中植物生長旺盛期或后期,而且不同時期2種草本植物的抗旱性不同。除了作為草坪草,這2種植物還被用于廢棄礦區(qū)和污染土壤的生態(tài)修復(fù)。張詩吟等研究了新疆煤礦廢棄地7種牧草的葉綠素熒光特征與抗旱性的關(guān)系,發(fā)現(xiàn)高羊茅較多年生黑麥草具有更強的抗旱性[10]。孫偉等在對湖南省鉛鋅尾礦污染區(qū)適生植物進行篩選時發(fā)現(xiàn),黑麥草對鉛和鋅有較強的遷移能力和富集能力[11]。谷超等研究了紅楓湖底泥中重金屬污染的植物修復(fù)特征,發(fā)現(xiàn)高羊茅對鋅的吸收能力較強,黑麥草對銅和鉛的吸收能力較強[12]。
種子萌發(fā)作為植物生活史的關(guān)鍵階段(包括幼苗),決定著植物對環(huán)境的適應(yīng)性和生存能力[13]。目前多年生黑麥草和高羊茅作為水土保持草種在江西稀土礦植被恢復(fù)區(qū)有所種植,但是它們對稀土尾砂生境適應(yīng)性的研究尚且缺乏。本研究采用不同濃度(0~30%)聚乙二醇(PEG-6000)模擬稀土尾砂干旱對多年生黑麥草和高羊茅種子萌發(fā)與幼苗生長的影響,比較2種植物的抗旱性,旨在為稀土尾砂礦區(qū)混播草種的選擇提供依據(jù)。
1 材料與方法
1.1 試驗材料
供試草種均采自尋烏廢棄稀土礦區(qū)周邊,采集時間為試驗前1年,室內(nèi)常溫條件保存。
供試稀土尾砂基質(zhì)取自江西省尋烏縣某廢棄稀土礦尾砂區(qū),采集表層0~20 cm范圍內(nèi)的尾砂,對其理化性狀指標進行測定。根據(jù)國際制土壤質(zhì)地分類,該尾砂土壤粒徑在 2.00~0.02 mm范圍的含量約占65%,屬于沙質(zhì)壤土,田間持水量15.02%,容重2.25 g/cm3;pH值為5.08,屬于強酸性,電導(dǎo)率高達79.12 dS/m,是原紅壤(6.9 dS/m)的10倍多,尾砂中土壤全磷含量、全氮含量、有機碳含量、有機質(zhì)含量分別為1.21 g/kg、0.20 mg/kg、6.29、10.85 g/kg,速效磷含量、速效鉀的含量分別為13.78、225.97 mg/kg。由此可見,尾砂土壤保水能力差,養(yǎng)分含量低。
1.2 試驗設(shè)計與測定方法
1.2.1 試驗設(shè)計及測定 2017年4月中旬在南昌工程學(xué)院水土保持實驗室的培養(yǎng)箱中進行種子萌發(fā)試驗。選擇飽滿、種皮完好的種子,用70%乙醇消毒5 min,蒸餾水沖洗4~5次。設(shè)定PEG-6000濃度為5%、10%、15%、20%、25%、30%共6個處理,同時以蒸餾水(CK1)、稀土尾砂浸提液(CK2)(浸提比例為1 ∶ 1,過濾后試驗備用)為對照,其中5%、10%為輕度脅迫,15%、20%為中度脅迫,25%、30%為重度脅迫。將準備好的種子置于墊有2層濾紙的培養(yǎng)皿中,分別加入配置好的處理液,每個培養(yǎng)皿放50粒種子,重復(fù)3次,置于人工氣候箱中,設(shè)定溫度為白天25 ℃,夜晚20 ℃,每天光照時間為08:00—18:00,共10 h,光照度為2 000 lx。每天定時統(tǒng)計種子發(fā)芽數(shù)(以種子露白為準),2 d計算發(fā)芽勢,9 d計算發(fā)芽率、發(fā)芽指數(shù)[7]。每2 d更換1次濾紙。
發(fā)芽率=(9 d內(nèi)種子發(fā)芽數(shù)/供試種子數(shù))×100%;
發(fā)芽勢=(2 d內(nèi)種子發(fā)芽數(shù)/供試種子數(shù))×100%;
發(fā)芽指數(shù)=∑(Gt/Dt)。
式中:Gt為t d的發(fā)芽數(shù);Dt為發(fā)芽時間,d。
1.2.2 幼苗滲透調(diào)節(jié)物質(zhì)測定 種子萌發(fā)試驗的同時,在南昌工程學(xué)院生物技術(shù)基地進行幼苗生長試驗。將消毒過的種子播種于裝有稀土尾砂的育苗盆(直徑15 cm×高10 cm)內(nèi),待種子在自然條件下生長,試驗時期為南昌雨季,無須添加水分。試驗50 d后,每盆留生長一致的幼苗15株,3個重復(fù)。將育苗盆移入遮雨棚下,每天09:00用濃度為10%、20%、30%的PEG-6000溶液對多年生黑麥草和高羊茅幼苗進行輕度、中度、重度干旱處理,處理液體積為20 mL,蒸餾水處理為對照CK。處理7 d后采集幼苗地上部分,迅速用冰盒帶回實驗室低溫-80 ℃保存,用于測定滲透調(diào)節(jié)物質(zhì)含量。其中葉綠素含量的測定采用分光光度計法;丙二醛含量采用硫代巴比妥酸法測定;可溶性蛋白質(zhì)含量采用考馬斯亮藍G-250染色法測定;游離脯氨酸含量采用磺基水楊酸法提取測定;植物組織中游離氨基酸含量采用茚三酮法測定[14]。
1.3 數(shù)據(jù)處理
采用SPSS 17.0對數(shù)據(jù)進行單因素方差分析和多重比較;用隸屬函數(shù)值法進行植物抗旱性綜合評價。
耐旱性綜合評價采用隸屬函數(shù)值法[15]。計算公式如下:
(1)
式中:u(Xij)為i物種j指標的隸屬函數(shù)值;Xij為i物種j指標的測定值;Xjmax和Xjmin分別為j指標的最大值和最小值。
如果某一指標與耐旱性呈負相關(guān),則用反隸屬函數(shù)計算:
(2)
運用公式(1)或公式(2)求出不同植物多項抗旱指標的平均隸屬函數(shù)值,平均隸屬函數(shù)值越大,表示該種植物抗旱性越強。
2 結(jié)果與分析
2.1 模擬干旱對2種植物種子萌發(fā)的影響
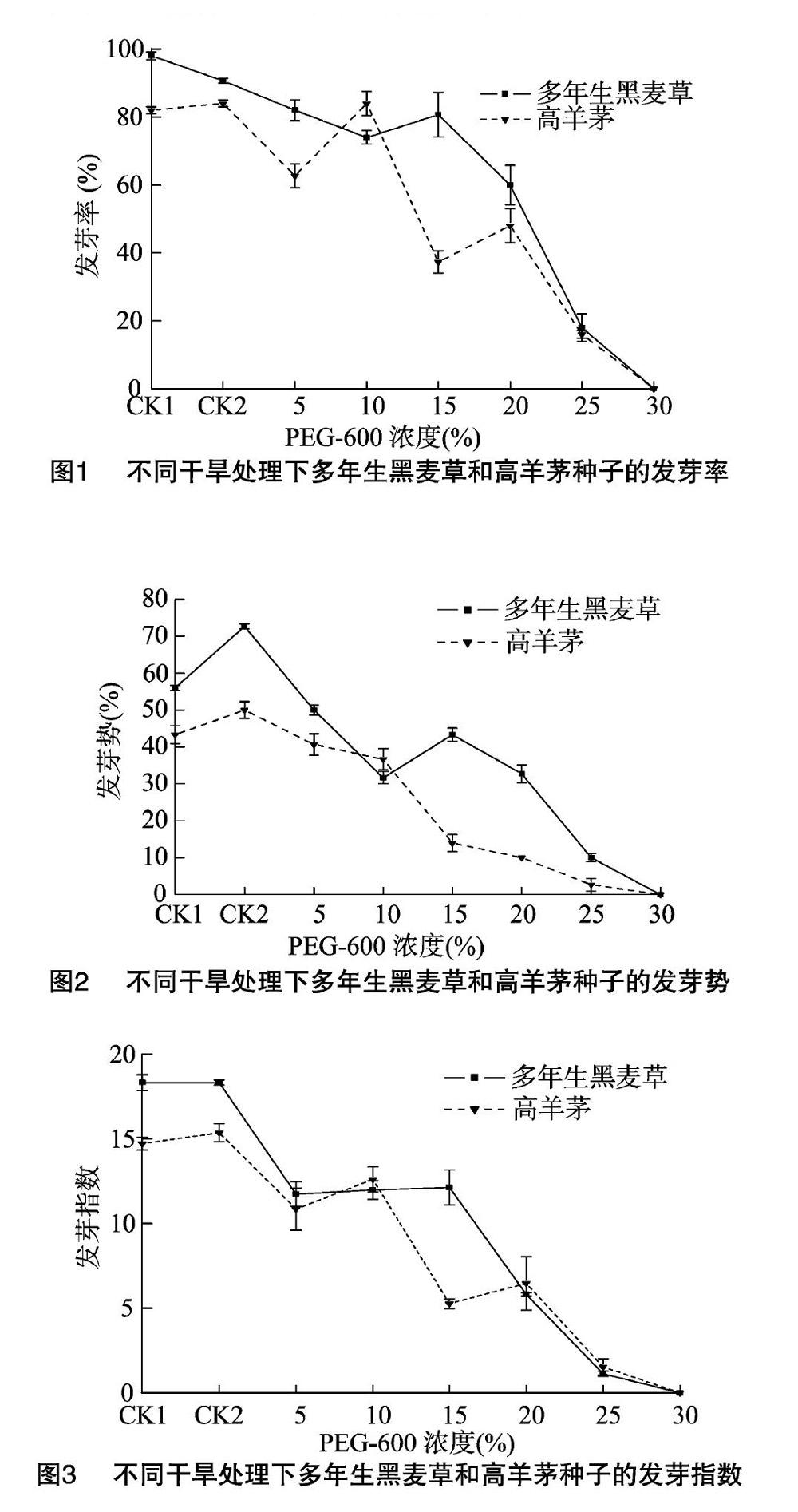
2.1.1 發(fā)芽率 不同處理下,隨著干旱程度的加強,多年生黑麥草和高羊茅的發(fā)芽率呈下降趨勢;PEG-6000濃度為5%~30%時,其發(fā)芽率低于蒸餾水(CK1)、浸提液(CK2);與蒸餾水CK1相比,浸提液CK2中多年生黑麥草的發(fā)芽率明顯降低,兩者處理的高羊茅則不差異明顯;多年生黑麥草和高羊茅的發(fā)芽率分別在20%和15%脅迫時明顯降低,說明中度脅迫大幅抑制了2種植物的種子萌發(fā);25%濃度脅迫時,二者發(fā)芽率為17%左右,30%時種子停止發(fā)芽(圖1)。
2.1.2 發(fā)芽勢和發(fā)芽指數(shù) 多年生黑麥草和高羊茅的發(fā)芽勢、發(fā)芽指數(shù)均隨著干旱程度增強呈降低趨勢。多年生黑麥草的發(fā)芽勢在CK2條件下明顯高于CK1,發(fā)芽指數(shù)在這2個條件下無明顯差異;高羊茅的發(fā)芽勢、發(fā)芽指數(shù)在CK1和CK2之間均無明顯差異,且多年生黑麥草的發(fā)芽勢、發(fā)芽指數(shù)明顯高于高羊茅。在PEG-6000的濃度為5%~30%條件下,2種植物的發(fā)芽勢和發(fā)芽指數(shù)均明顯低于CK1、CK2;相較于CK1,高羊茅的發(fā)芽勢、發(fā)芽指數(shù)在15%的脅迫下分別明顯降低了 67.69%、64.20%,多年生黑麥草則在20%脅迫下明顯降低了41.61%、68.30%。另外,輕度(5%、10%)脅迫和重度(25%、30%)脅迫條件下的發(fā)芽勢和發(fā)芽指數(shù)在2種植物之間無明顯差異(圖2、圖3)。
2.2 模擬干旱對2種植物幼苗生理特性的影響
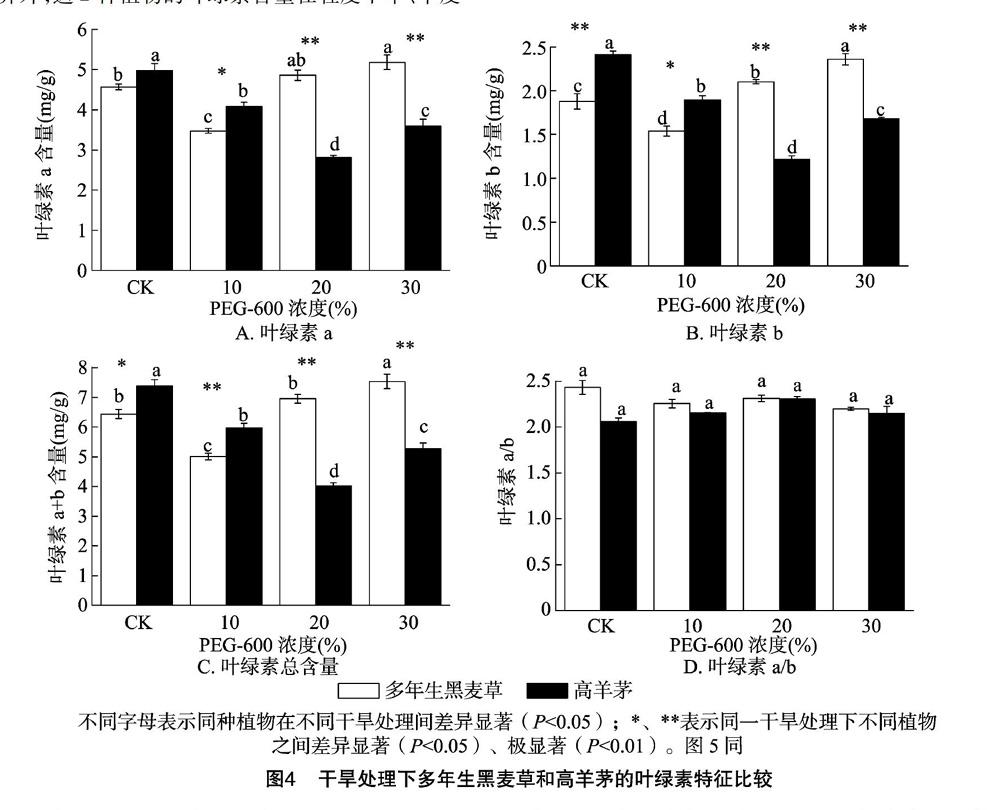
2.2.1 葉綠素含量 多年生黑麥草和高羊茅的葉綠素a、葉綠素b和葉綠素a+b含量變化趨勢隨干旱脅迫(10%~30% PEG-6000濃度)的增強基本一致,多年生黑麥草的葉綠素a、葉綠素b、葉綠素a+b含量隨著脅迫程度增強而升高,高羊茅則呈先下降后升高的趨勢(圖4-A、圖4-B、圖4-C);除了多年生黑麥草的葉綠素a含量在中度干旱和重度干旱之間無顯著差異外,這2種植物的葉綠素含量在輕度干旱、中度干旱和重度干旱之間差異顯著。對相同處理來說,對照和輕度脅迫下高羊茅的葉綠素含量高于多年生黑麥草,而中度和重度脅迫下的多年生黑麥草的葉綠素含量極顯著高于高羊茅。對葉綠素a/b來說,不管是不同處理間還是不同植物之間均無顯著差異(圖4-D)。
2.2.2 滲透調(diào)節(jié)物質(zhì) 隨干旱脅迫程度(10%~30%PEG-6000)的增強,多年生黑麥草幼苗的丙二醛含量逐漸降低,重度干旱時最低,比對照降低了28.31百分點(圖5-A),可溶性蛋白質(zhì)和游離氨基酸含量呈相似的變化趨勢,在輕度脅迫、重度脅迫條件下較高,與對照和中度脅迫時差異顯著(圖5-B、圖5-C);中度干旱、重度干旱脯氨酸含量顯著高于對照和輕度干旱(圖5-D),說明不同程度的干旱脅迫可誘使多年生黑麥草通過不同的滲透物質(zhì)來維持生長。隨干旱脅迫的增強高羊茅的丙二醛含量呈現(xiàn)增高趨勢、可溶性蛋白含量、游離氨基酸含量先降低后升高,重度脅迫下游離等基酸含量顯著高于低中度脅迫(圖5-A、圖5-B、圖5-C)。對這2種植物來說,不管是在哪種脅迫程度下,多年生黑麥草的丙二醛含量、可溶性蛋白含量、游離氨基酸的含量均顯著高于高羊茅;而且干旱脅迫下多年生黑麥草各滲透調(diào)節(jié)物質(zhì)的變化幅度高于高羊茅,說明多年生黑麥草在應(yīng)對干旱脅迫時具有更大潛力(圖5)。
2.3 多年生黑麥草和高羊茅各指標的相關(guān)性
多年生黑麥草和高羊茅幼苗生長中各指標相關(guān)性分析(表1)顯示,葉綠素a含量、葉綠素b含量和葉綠素a+b之間呈均極顯著正相關(guān),這3個指標與葉綠素a/b之間呈負相關(guān),但相關(guān)性不顯著。滲透調(diào)節(jié)物質(zhì)中,丙二醛含量、可溶性蛋白質(zhì)含量與游離氨基酸含量兩兩之間存在極顯著的正相關(guān)關(guān)系,相關(guān)系數(shù)分別為0.657、0.586、0.817。
2.4 多年生黑麥草和高羊茅對稀土尾砂干旱適應(yīng)性的綜合評價
通過11個指標對干旱處理下多年生黑麥草、高羊茅種子萌發(fā)和幼苗生長的適應(yīng)性進行綜合評價,從表2可以看出,多年生黑麥草、高羊茅各指標的平均隸屬函數(shù)值為0.639、0.329,說明多年生黑麥草對稀土尾砂干旱脅迫的適應(yīng)性高于高羊茅。
3 討論與結(jié)論
稀土尾砂廢棄地土壤結(jié)構(gòu)嚴重沙化,保水能力差,植物易遭受水分脅迫。種子為植物最重要的繁殖器官,其種子萌發(fā)和幼苗階段的生長對植物的成活起著決定性的作用,較其他階段更容易受到水分等環(huán)境因子的影響。采用PEG-6000模擬水分脅迫,是通過抑制種子吸水過程而抑制其萌發(fā),隨著干旱脅迫的增強,多數(shù)植物種子的發(fā)芽率、發(fā)芽勢、發(fā)芽指數(shù)均呈下降趨勢[16-18],也有低濃度PEG-6000處理促進植物種子萌發(fā)的報道[19]。在本試驗中,多年生黑麥草和高羊茅的發(fā)芽情況隨脅迫程度增強呈下降趨勢,說明干旱抑制了這2種植物的種子萌發(fā),輕度脅迫也未起到促進作用,這與史薇等的研究結(jié)果[16,20]一致。當PEG-6000濃度分別為20%、15%時顯著降低,說明多年生黑麥草較高羊茅在中度脅迫下具有較強的發(fā)芽能力;當濃度達到25%時,2種植物的發(fā)芽率、發(fā)芽勢、發(fā)芽指數(shù)極低,30%濃度下,種子失去萌發(fā)能力。
葉綠素含量是植物對干旱脅迫反應(yīng)敏感性的生理指標,大量的研究結(jié)果表明,干旱脅迫通過抑制葉綠素合成并加速其分解使葉綠素含量直線下降[21]。本研究顯示,隨著干旱脅迫加劇,多年生黑麥草和高羊茅的葉綠素含量先降低后升高,分別在輕度(10%)和中度(20%)干旱脅迫下最低,且中度和重度干旱脅迫下,多年生黑麥草的葉綠素含量顯著高于高羊茅,說明在中度、重度干旱條件下,多年生黑麥草受到影響較小,具有更強的光合潛力。
滲透調(diào)節(jié)作用是植物適應(yīng)水分脅迫的重要機制,其中丙二醛是膜脂過氧化的產(chǎn)物[22],通過丙二醛的變化可了解干旱脅迫對植物的破壞程度;游離氨基酸、脯氨酸和可溶性蛋白是植物體內(nèi)的主要滲透調(diào)節(jié)物質(zhì),三者均能通過降低細胞原生質(zhì)的滲透勢,促使細胞從外界吸收水分,使植物保持一定的含水量和膨壓[23],減輕脅迫對植物體的傷害[24-25]。本試驗中,不管是在輕度、中度干旱脅迫下還是在重度干旱脅迫下,多年生黑麥草的丙二醛、可溶性蛋白質(zhì)和游離氨基酸的含量均顯著高于高羊茅,且變化幅度較大,說明多年生黑麥草受到干旱脅迫時,其體內(nèi)的滲透調(diào)節(jié)物質(zhì)大量積累,減少干旱對細胞膜造成破壞,使植物具有更強的滲透調(diào)節(jié)能力。相關(guān)性分析也表明,丙二醛含量與游離氨基酸和可溶性蛋白的含量極顯著相關(guān)(r=0.586、r=0.657),這與王曉龍等的研究結(jié)果[26]一致。
也有研究指出,大多滲透調(diào)節(jié)物質(zhì)均隨干旱脅迫的增加呈遞增或者先升高后降低的趨勢[7,16],但是不同植物在不同環(huán)境條件下各不相同。本研究中多年生黑麥草和高羊茅葉片中脯氨酸含量隨脅迫加劇而上升,這與以往研究結(jié)果[21,23]一致;而可溶性蛋白質(zhì)和游離氨基酸含量則呈現(xiàn)先上升后降低然后又上升的趨勢,一方面與稀土尾砂復(fù)雜的土壤環(huán)境脅迫有關(guān),另一方面輕度脅迫(10%)下植物的滲透調(diào)節(jié)能力占主導(dǎo)地位,葉片中可溶性蛋白質(zhì)和游離氨基酸含量很高,中度脅迫(20%)時可能激發(fā)抗氧化酶系統(tǒng)活性,大量氨基酸和可溶性蛋白合成保護酶類物質(zhì)含量顯著降低[21,24],而在重度干旱脅迫下,酶活性受到抑制,其合成減弱,進而使游離氨基酸和可溶性蛋白質(zhì)含量大幅升高[17]。不同滲透調(diào)節(jié)物質(zhì)的相關(guān)性分析表明,游離氨基酸含量和可溶性蛋白質(zhì)含量呈極顯著正相關(guān),與脯氨酸呈負相關(guān),主要是由于游離氨基酸是合成植物生長所需蛋白質(zhì)和酶類的基本物質(zhì)[27],而游離氨基酸的種類有19種,不同氨基酸的調(diào)節(jié)能力和響應(yīng)時間存在差異。
在不同的PEG濃度下,各種植物的抗旱性程度不會保持不變,植物種子萌發(fā)和幼苗階段的抗旱性信息不同,應(yīng)該對生長階段的抗旱性分別評價[28],但是在實際生產(chǎn)中,種子萌發(fā)特性很好地植物可能在幼苗期由于高溫或者缺水而死亡,只有在種子萌發(fā)和幼苗階段都有較高抗旱性的植物才能很好地存活。因此,本研究中將種子萌發(fā)和幼苗階段作為整體進行隸屬函數(shù)綜合評價,結(jié)果表明,多年生黑麥草具有更強的抗旱性。
稀土尾砂干旱環(huán)境下,多年生黑麥草較高羊茅具有更強發(fā)芽率、發(fā)芽勢、發(fā)芽指數(shù),幼苗生長階段,植物通過葉綠素、丙二醛、可溶性蛋白、游離氨基酸等生理調(diào)節(jié)作用來維持生長,多年生黑麥草對稀土尾砂干旱環(huán)境有更好的適應(yīng)性。在稀土礦尾砂區(qū)采用植物措施修復(fù)時,可優(yōu)先考慮選擇冷季草多年生黑麥草,與暖季草混合進行快速覆綠。
參考文獻:
[1]Li X F,Chen Z B,Chen Z Q,et al. A human health risk assessment of rare earth elements in soil and vegetables from a mining area in Fujian Province,Southeast China[J]. Chemosphere,2013,93(6):1240-1246.
[2]Liang T,Li K X,Wang L Q. State of rare earth elements in different environmental components in mining areas of China[J]. Environmental Monitoring and Assessment,2014,186(3):1499-1513.
[3]劉文深,劉 暢,王志威,等. 離子型稀土礦尾砂地植被恢復(fù)障礙因子研究[J]. 土壤學(xué)報,2015,52(4):879-887.
[4]榮秀連,王 波,劉 刊,等. PEG-6000模擬干旱脅迫對冷季型草坪種子萌發(fā)特性影響[J]. 北方園藝,2010(8):80-82.
[5]鄭軼琦,臧國長. PEG脅迫對冷季型草坪草種子萌發(fā)和幼苗生長的影響[J]. 種子,2011,30(12):34-37.
[6]初曉輝,岳信龍,任 健,等. 水分脅迫下4種冷季型草坪草抗旱性比較研究[J]. 草原與草坪,2012,32(4):15-19.
[7]侯 敏. 干旱脅迫對3種草坪草的生長及葉綠素含量的影響[J]. 山西林業(yè)科技,2015,44(3):24-25.
[8]趙志軍,單貴蓮,段新慧,等. 干旱脅迫對3種冷季型草坪草光譜反射率及生理特征的影響[J]. 草原與草坪,2016,36(6):23-29.
[9]高 涵,吳 偉,劉秀萍,等. 水分脅迫下幾種冷季型草坪草抗旱機理研究[J]. 水土保持研究,2006,13(3):126-128.
[10]張詩吟,楊建軍,莫 愛. 干旱脅迫下七種牧草葉綠素熒光參數(shù)與抗旱性關(guān)系研究[J]. 湖北農(nóng)業(yè)科學(xué),2017,56(1):107-111.
[11]孫 偉,王 瓊,張 望,等. 湖南省鉛鋅尾礦污染區(qū)植被修復(fù)適生植物選擇研究[J]. 價值工程,2016,35(17):208-211.
[12]谷 超,梁隆超,陳 卓. 4種牧草植物對紅楓湖底泥中重金屬污染的植物修復(fù)研究[J]. 環(huán)境工程,2015,33(7):148-151.
[13]張佳寧,劉 坤. 植物調(diào)節(jié)萌發(fā)時間和萌發(fā)地點的機制[J]. 草業(yè)學(xué)報,2014,23(1):328-338.
[14]李合生. 植物生理生化實驗原理和技術(shù)[M]. 北京:高等教育出版社,2000.
[15]種培芳,蘇世平,李 毅. 4個地理種群紅砂的抗旱性綜合評價[J]. 草業(yè)學(xué)報,2011,20(5):26-33.
[16]史 薇,徐海量,趙新風(fēng),等. 脹果甘草種子萌發(fā)對干旱脅迫的生理響應(yīng)[J]. 生態(tài)學(xué)報,2010,30(8):2112-2117.
[17]Abudureheman B,Yan C,Wei Y. Fruit growth and seed germination characteristics of Nanophyton erinaceum:a dominant desert shrub[J]. International Journal of Plant Research,2012,25(1):34-40.
[18]Zeng Y J,Wang Y R,Baskin C C,et al. Testing seed germination responses to water and salinity stresses to gain insight on suitable microhabitats for restoration of cold desert shrubs[J]. Journal of Arid Environments,2014,100(1):89-92.
[19]閆興富,周立彪,思彬彬,等. 不同溫度下PEG-6000模擬干旱對檸條錦雞兒種子萌發(fā)的脅迫效應(yīng)[J]. 生態(tài)學(xué)報,2016,36(7):1989-1996.
[20]胡 紅,曹 昀,王 穎. 水分脅迫對狗牙根種子萌發(fā)及幼苗生長的影響[J]. 草業(yè)科學(xué),2013,30(1):63-68.
[21]桑子陽,馬履一,陳發(fā)菊. 干旱脅迫對紅花玉蘭幼苗生長和生理特性的影響[J]. 西北植物學(xué)報,2011,31(1):109-115.
[22]董伊晨,劉悅秋. 土壤水分對異株蕁麻(Urtica dioica)保護酶和滲透調(diào)節(jié)物質(zhì)的影響及其與葉片光合和生物量的相關(guān)性[J]. 生態(tài)學(xué)報,2009,29(6):2845-2851.
[23]李 力,劉玉民,王 敏,等. 3種北美紅楓對持續(xù)高溫干旱脅迫的生理響應(yīng)機制[J]. 生態(tài)學(xué)報,2014,34(22):6471-6480.
[24]謝志玉,張文輝,劉新成. 干旱脅迫對文冠果幼苗生長和生理生化特征的影響[J]. 西北植物學(xué)報,2010,30(5):948-954.
[25]丁玉梅,馬龍海,周曉罡,等. 干旱脅迫下馬鈴薯葉片脯氨酸、丙二醛含量變化及與耐旱性的相關(guān)性分析[J]. 西南農(nóng)業(yè)學(xué)報,2013,26(1):106-110.
[26]王曉龍,李 紅,楊 曌,等. 干旱脅迫下4種禾本科牧草根系抗旱性比較[J]. 中國飼料,2016(13):22-24,32.
[27]張海娜,方向文,蔣志榮,等. 檸條平茬處理后不同組織游離氨基酸含量[J]. 生態(tài)學(xué)報,2011,31(9):2454-2460.
[28]羅 冬,王明玖,李元恒,等. 四種豆科牧草種子萌發(fā)和幼苗生長對干旱的響應(yīng)及抗旱性評價[J]. 生態(tài)環(huán)境學(xué)報,2015,24(2):224-230.
