CiCHI提高擬南芥總黃酮含量
2019-11-21 11:09:14楊飛蕓白潔劉坤王瑞剛
生物技術通報 2019年11期
楊飛蕓 白潔 劉坤 王瑞剛
(1. 內蒙古農業大學生命科學學院 內蒙古自治區植物逆境生理與分子生物學重點實驗室,呼和浩特 010011;2. 內蒙古農業大學食品科學與工程學院,呼和浩特 010018)
類黃酮(Flavonoids)是廣泛存在于植物中的次級代謝產物,泛指2個苯環通過脂肪族三碳鏈相互聯結而成的一系列低分子量酚類化合物[1]。迄今為止,已有10000多個類黃酮類化合物被分離鑒定[2]。類黃酮分布廣泛,在植物的生長、發育、開花、結果及耐鹽、抗旱、抵抗紫外線、抗菌、抗病等方面都起著重要作用[3-4]。類黃酮類物質是具有多種生物學活性的一類植物次級代謝產物,具有重要的食用和藥用價值,一直以來都受到國內外眾多研究者的高度重視[5-6]。近年來,類黃酮研究領域一直非常活躍,并取得許多重要進展[7]。目前已發現其在人體內可以發揮抗氧化活性、保護肝臟活性、抗菌活性、抗炎活性及抗癌活性及抗病毒活性等[8-9]。
類黃酮是在胞漿多酶復合體的催化下,經由苯丙烷代謝途徑合成的[10]。參與類黃酮生物合成途徑的酶主要包括:查爾酮合成酶(Chalcone synthase,CHS)、查爾酮異構酶(Chalcone isomerase,CHI)和黃酮醇合成酶(Flavonol synthase,FLS)等[11]。查爾酮異構酶(EC 5.5.1.6)是類黃酮合成途徑中的第二個關鍵酶,催化分子內的環化反應[12]。查爾酮異構酶是一個多基因家族,但由于在進化上源自共同的祖先,因此,不同基因的編碼區同源性很高,其結構和催化機制也極其相似。但在不同物種中,其基因拷貝數和時空表達模式存在很大差異,并且受到多種因素的誘導[13]。查爾酮異構酶是植物體內花青素以及其他類黃酮合成的必需酶,通常以單體的形式存在于植物中,由于其蛋白三維折疊結構的特異性,常作為植物特有的標記基因[14]。作為類黃酮合成途徑上游的重要酶,查爾酮異構酶對調控整個代謝途徑起著重要作用,直接影響下游各種類黃酮化合物,如異黃酮、花青素等的含量。近年來應用外源CHI基因轉化植物的研究報道很多,主要集中在改變觀賞植物的花色和提高果蔬、藥用植物中類黃酮的含量等方面[15]。
中間錦雞兒(Caragana intermedia)是豆科錦雞兒屬植物,落葉灌木,具有廣泛的適應性和很強的抗逆性,在長期抵御逆境的過程中,形成了一系列適應在惡劣環境下生存的特有機制[16-17]。中間錦雞兒營養物質含量豐富,還含有類黃酮、芪類、苯丙素和萜類等多種次級代謝產物,是傳統中、蒙藥藥材,其花、根、種子等部位均可入藥[18-19]。但對其抗逆機制、類黃酮代謝及其與抗逆性之間的關系鮮見報道。
本研究以豆科已知CHI基因的保守結構域設計簡并引物,以中間錦雞兒cDNA為模板,克隆CiCHI的cDNA全長,并對其進行了序列分析和系統進化分析,通過轉基因擬南芥研究CiCHI的功能,為進一步揭示中間錦雞兒的抗逆機制提供理論依據。
1 材料與方法
1.1 材料
中間錦雞兒種子采自內蒙古自治區呼和浩特市和林格爾縣(111.82°E,40.38°N)。野生型擬南芥Columbia-0生態型(Col-0)、大腸桿菌DH5α、根癌農桿菌GV3101和植物表達載體質粒pCanG-HA由內蒙古自治區植物逆境生理與分子生物學重點實驗室保存。T載體pMD19-T Vector購自TaKaRa公司。
1.2 方法
1.2.1CiCHI的克隆及分析 采用TRIzol法[20]提取中間錦雞兒和擬南芥總RNA。去除樣品中的DNA后,反轉錄合成cDNA第一鏈。
根據豆科已知CHI基因的保守結構域設計簡并引物(表1),以中間錦雞兒cDNA為模板,使用Clontech公司的RACE試劑盒獲得3'-和5'-末端序列。通過分析、比對、拼接后得到CiCHI的全長序列。PCR引物合成及產物測序由上海生工生物公司完成。
將CiCHI的 ORF(Open reading frame) 序 列在NCBI上進行Blast比對,篩選出與其相似度最高的序列進行系統進化分析。利用Mega 5.0軟件,采用Neighbour-Joining算法構建系統進化樹[21]。不同物種的CHI基因序列分別從NCBI數據庫(https://www.ncbi.nlm.nih.gov/)、TAIR 數 據 庫(http://www.arabidopsis.org/)和植物基因組數據庫(https://phytozome.jgi.doe.gov/pz/portal.html)中獲得。
1.2.2 中間錦雞兒的處理方法 選取植物培養室(溫度22℃,16 h光照/8 h黑暗)生長一個月左右、長勢一致的中間錦雞兒幼苗進行脅迫處理,檢測CiCHI的表達量變化。每個樣品均為3株幼苗地上部分的混合樣品。紫外(UV-C)輻照處理(紫外光強度為120.5 lx,紫外燈照射距離為1 m)、NaCl和脫落酸(Abscisic acid,ABA)脅迫處理(NaCl濃度200 mmol/L,ABA濃度200 μmol/L)的取樣時間為0、0.5、1、3、6、9、12和24 h。進行3次生物學重復。提取脅迫處理樣品的總RNA,以CkEF1α(KC679842)作為內參基因,利用qRT-PCR在Light Cycler 480實時熒光定量PCR儀(Roche公司)上對CiCHI的表達量進行檢測。基因表達量以2-ΔCT法計算。
1.2.3 pCanG-CiCHI植物表達載體的構建 獲得CiCHI編碼區(ORF),并插入克隆載體pEASY-Blunt Simple中(北京全式金生物公司),將測序驗證后的序列用In-Fusion酶連入由CaMV35S啟動子驅動的植物表達載體pCanG-HA中,進行菌落PCR及雙酶切驗證,將驗證正確的重組質粒電轉化農桿菌GV3101感受態細胞,進行菌落PCR鑒定。
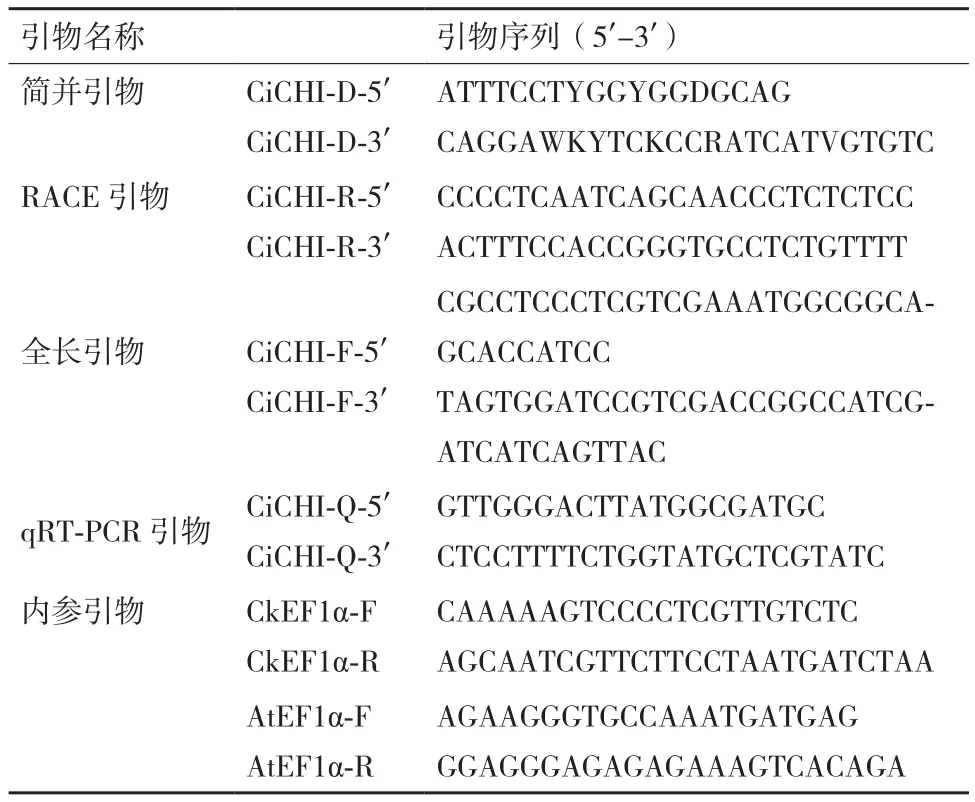
表1 引物序列
1.2.4 擬南芥遺傳轉化及純合體篩選 采用浸花法將重組植物表達載體pCanG-CiCHI轉入野生型擬南芥(Col-0)[22],收取成熟的種子種在含有 25 μg/mL卡那霉素的1/2 MS平板培養基上篩選陽性植株。提取T3代轉基因植株總RNA,反轉錄成cDNA,利用特異引物CiCHI-F-5'和CiCHI-F-3'進行PCR鑒定。利用qRT-PCR對CiCHI在轉基因株系中的表達量進行檢測。反應體系為SYBR Premix ExTaq(TaKaRa公司)10 μL、稀釋的cDNA模板5 μL、上、下游引物各 0.8 μL(10 μmol/L)和 DEPC 水 3.4 μL。反應程序為 95℃ 30 s;95℃ 5 s,60℃ 30 s,72℃ 15 s,40個循環。AtEF1α(AT5G60390)作為內參基因,基因表達量以2-ΔCT法計算,選取4株表達水平較高的不同轉化株系進行后續表型試驗。
1.2.5 轉基因擬南芥總黃酮含量檢測 剪取正常生長條件下(22℃,16 h光照/8 h黑暗)40日齡的擬南芥植株地上部分,每個株系取3株植物,用液氮研磨成粉末。采用硝酸鋁比色法進行總黃酮含量的測定[23],3次生物學重復。
2 結果
2.1 CiCHI的克隆與序列分析
據豆科已知CHI基因的保守序列設計簡并引物,擴增得到509 bp的單一條帶(圖1-A),經測序后比對,發現該片段與蒺藜苜蓿CHI基因相似度最高(84%)。利用RACE技術克隆其缺失的cDNA序列,分別得到248 bp的5'-端和473 bp的3'-端cDNA序列(圖1-B)。
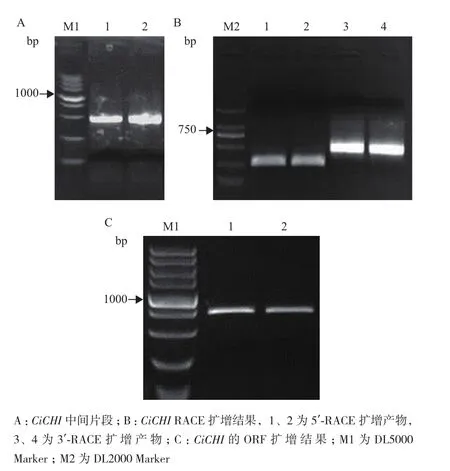
圖1 CiCHI的克隆
經測序分析正確后,利用Vector NTI 10將克隆到的序列片段進行拼接,得到CiCHI的cDNA全長序列。以中間錦雞兒cDNA為模板,使用特異引物擴增其cDNA序列(圖1-C),連接克隆載體驗證后,確定拼接結果正確。得到CiCHI的cDNA全長為1005 bp, 其 中 ORF為 678 bp,5'-UTR 為 110 bp,3'-UTR為217 bp,編碼225個氨基酸。
2.2 CiCHI蛋白的系統進化分析
為進一步了解CiCHI與其他物種該類基因之間的進化關系,將CiCHI蛋白序列與其他已知物種中相似度最高的CHI蛋白序列進行比對,并構建系統進化樹(圖2)。結果表明,CiCHI與蒺藜苜蓿的CHI最為相似,二者與其他豆科植物的CHI構成一個大分支;而同為蕓香科的蜜桔、柚子和甜橙構成一個分支。說明CiCHI與豆科植物的CHI功能最相近。進化樹中使用的序列都是NCBI中注釋為Chalcone isomerase(CHI)的基因,即查爾酮異構酶基因家族的成員,因此可以確定CiCHI為該基因家族的成員。
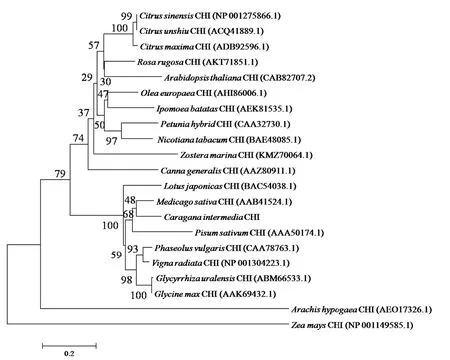
圖2 CiCHI與其他物種來源的CHI構建的系統進化樹
2.3 CiCHI脅迫處理下的表達分析
查爾酮異構酶催化類黃酮的生成,起到抵抗氧化脅迫的作用。通過檢測CiCHI在紫外、NaCl和ABA等脅迫處理下的表達模式(圖3)發現紫外照射處理下,CiCHI表達量呈先升高后降低的趨勢,在6 h時表達量最高(圖3-A)。NaCl處理下,CiCHI表達量同樣先升高后降低,在1 h時表達量最高(圖3-B)。ABA處理下,CiCHI的表達受到抑制,在1 h表達量最低(圖3-C)。
2.4 CiCHI的組織特異性分析
通過對30日齡中間錦雞兒根、莖、葉中CiCHI進行qRT-PCR分析(圖4)發現,CiCHI在所有被檢測的組織中都有不同程度的表達。在室內種植的葉中表達量最高,莖中相對較高,根中最少。在野外采摘的花中表達量高,種子中表達量低。
2.5 pCanG-CiCHI植物表達載體的構建及轉基因純合體植株的鑒定
對重組質粒進行雙酶切鑒定,可以獲得目的片段,表明載體構建成功。
通過農桿菌介導的浸花法獲得具有卡那霉素抗性的來自不同轉化的陽性植株10株,提取這些株系的總RNA并合成cDNA,利用qRT-PCR檢測CiCHI在轉基因株系中的表達水平(圖5),選取表達量較高的4個株系OE-13、OE-32、OE-37和OE-46進行后續表型檢測試驗。
2.6 轉基因擬南芥總黃酮含量的變化
按照硝酸鋁比色法的具體步驟繪制蘆丁標準曲線(圖6-A)。蘆丁質量濃度X和吸光值Y的關系為:Y=1.081X-0.004,R2=0.999,表明在蘆丁濃度0-0.96 mg/mL中,該標準曲線線性良好。按此方法進行轉基因擬南芥總黃酮含量的測定(圖6-B)。結果表明,轉基因各株系總黃酮的含量均高于野生型,且達到顯著水平。說明過表達CiCHI可以使擬南芥中總黃酮含量明顯增加。
3 討論
類黃酮是錦雞兒中迄今為止被發現的最重要的活性成分,在植物-環境相互作用中發揮著極其重要的功能。類黃酮在植物體內的生物合成幾乎完全是由氧化脅迫引起的,是植物組織在不同脅迫下的抗氧化防御系統。它們可以吸收最具能量的太陽光(UV),抑制活性氧的產生,一旦活性氧過量形成可以將其及時清除;類黃酮可以調節生長素的運輸和代謝作用,對植物在脅迫誘導下形態的發生具有重要價值[24]。本研究檢測了轉基因擬南芥中總黃酮的含量,由于CiCHI的大量表達,4個轉基因株系的總黃酮含量均顯著高于野生型,說明CiCHI參與了擬南芥的類黃酮代謝,為研究植物類黃酮代謝提供了新的基因源。
圖3 不同脅迫處理下CiCHI的表達分析

圖4 CiCHI的組織特異性表達分析
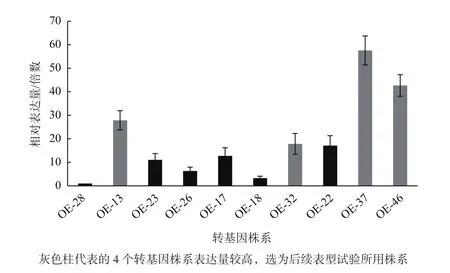
圖5 轉基因株系CiCHI表達水平檢測
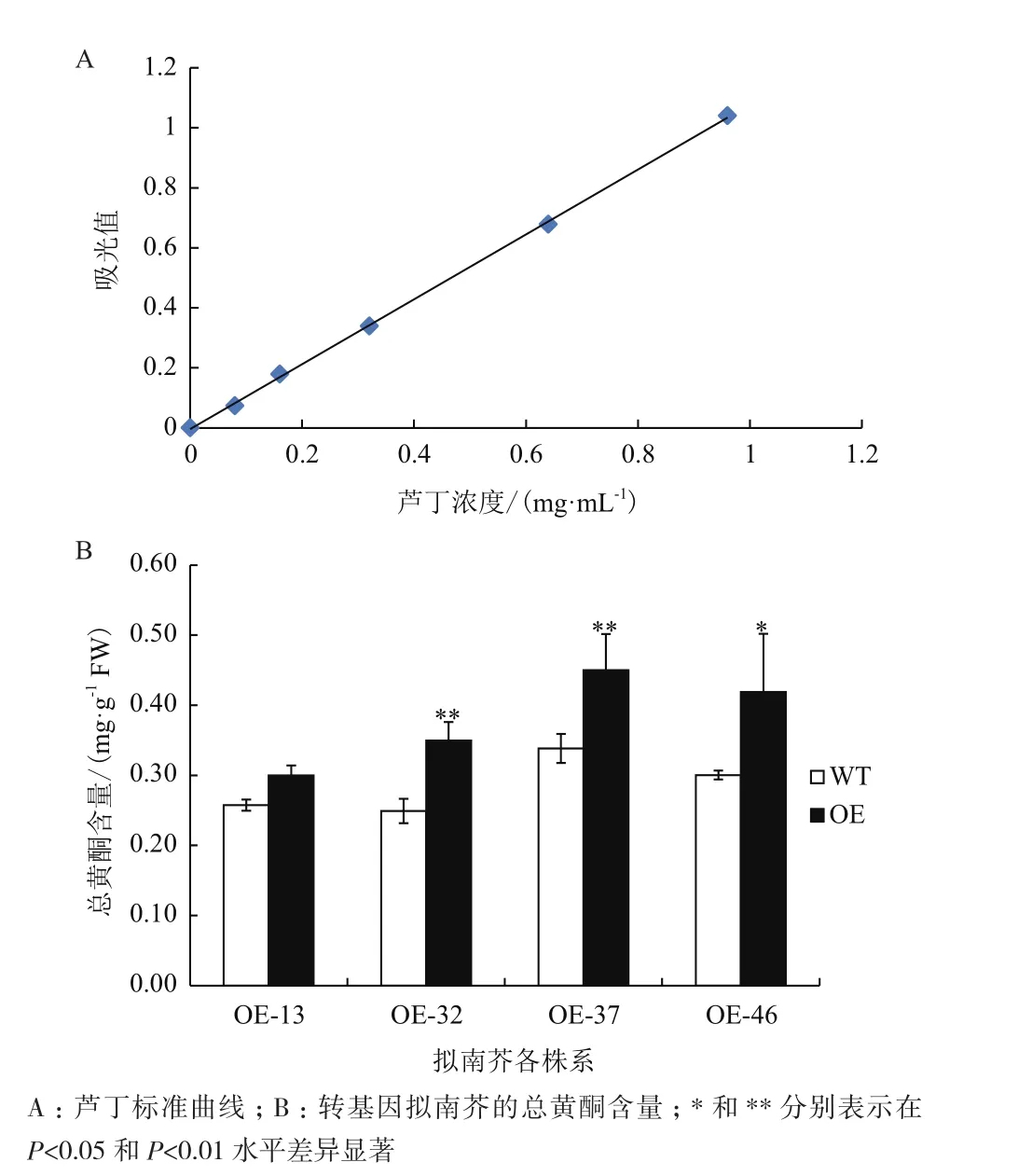
圖6 野生型和轉基因擬南芥總黃酮含量比較
查爾酮異構酶是類黃酮生物合成途徑的第二個限速酶,是植物體內類黃酮合成的必需酶,對調控整個代謝途徑起著重要作用,可使反應速率提高107倍,直接影響下游各種類黃酮化合物的生成[13]。系統進化分析顯示,CHI在整個進化過程中相對保守,序列相似性達51%-94%[14]。經過比對分析發現本研究獲得的CiCHI與蒺藜苜蓿的CHI基因相似度最高,達84%,可以確定其為查爾酮異構酶。
查爾酮異構酶基因家族不同成員的表達具有明顯的組織特異性[13]。本研究檢測了CiCHI在不同部位的表達量發現,其表達量存在很大差異。結果顯示CiCHI在室內種植的葉中表達量最高;莖中相對較高;根中最少。在野外采摘的花中表達量高,種子中表達量低,與已報道的研究結果一致[25-26]。由于根、莖、葉采自室內幼苗,花和種子采自野外自然生長的植物。因此,其生長條件、生長時間等各方面因素均不相同,使其表達量無法比較。
查爾酮異構酶家族基因受不同脅迫處理誘導,如UV、干旱、損傷和NaCl等,從而引起類黃酮含量的增加,使其抗氧化等抵御逆境的能力提高[15,27]。本研究發現中間錦雞兒CiCHI受到紫外、NaCl和ABA等處理的誘導,不同處理方式下該基因的表達模式不同。在受到紫外和NaCl脅迫時表達模式基本一致,開始受到脅迫影響時,表達量升高,生成大量類黃酮以幫助機體抵抗氧化脅迫的影響。但隨著脅迫壓力的增加,其表達量開始降低,可能其產物水平已經達到一定量并不再大量增加,此時機體也可能已經啟動了其他的脅迫響應機制。ABA處理使CiCHI的表達受到抑制。
4 結論
克隆得到中間錦雞兒CiCHI,其開放閱讀框678 bp,編碼225個氨基酸。CiCHI受到紫外、NaCl、ABA等脅迫誘導。在葉中表達量最高,莖中相對較高,根中最少;花中表達量高,種子中低。擬南芥中超表達CiCHI可使其總黃酮含量增加。
猜你喜歡
四川蠶業(2021年3期)2021-02-12 02:38:46
中成藥(2018年11期)2018-11-24 02:57:00
中成藥(2017年8期)2017-11-22 03:19:40
中成藥(2017年10期)2017-11-16 00:50:13
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
中成藥(2017年4期)2017-05-17 06:09:50
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
