基因組挖掘技術及其在真菌中的研究進展
2019-11-21 11:09:26徐杰黃建忠李力
生物技術通報 2019年11期
關鍵詞:生物
徐杰 黃建忠 李力
(福建師范大學生命科學院,福州 350108)
與傳統的以“模型導向”科研方式不同,基因組學研究以其高度的系統性,以數據為導向,開創了全新的研發方法。20世紀以來高通量基因測序技術的建立直接推動了生物信息學的發展[1],微生物基因組測序的成本不斷降低,周期縮短,越來越多的生物遺傳信息被公布,公共數據庫積累了大量的基因組序列數據,后基因組時代來臨[2]。在此基礎上,人們通過生物信息學軟件對不同基因結構進行識別鑒定、功能分析、相互關系和可能的表達調控機制,發現了大量未知的可能與合成新型次級代謝產物相關的基因簇,這為直接以基因為導向替代傳統的以生物活性為導向,尋找潛在藥物提供了可能[3]。
生物體次級代謝產物是抗生素的主要來源,其頻繁的使用會加速細菌適應性進化而產生免疫能力。研究證實,在對某一藥物產生抗性的同時,細菌對其結構類似物或衍生型藥物的抗藥性也會得到提高[4]。自 1979 年 Campbell[5]從 40000 份土壤樣品發現新型抗生素阿維菌素后至今的40年內,尚未找到其它有重要應用價值的活性化合物,以生物活性為導向的舊有藥物篩選方式陷入瓶頸。挖掘藥物的關鍵在于避免已知化合物的過度復篩,能否發現新藥物很大程度上取決于挖掘過程中在概率上的盲目程度。用科學理論指導開發方向,可降低成本和周期,避免已知化合物的重復研究。微生物中負責編碼次級代謝產物合成的酶基因通常成簇存在,通過NCBI中基礎局部比對搜索工具(BLAST)搜索相關酶類,并通過基因預測算法,如Augustus發現未知的生物合成基因簇,通過信息學工具可預測出產物結構以及可能含有的新藥物作用靶點[6-8],這為新藥開發提供了新方向。
次級代謝產物是一種具有多樣化結構的小分子,它在微生物體內不是為了滿足機體基本生長需要而產生,而是作為一種“武器”來對抗復雜的生態環境并保證在與其它微生物的競爭中處于優勢地位。然而編碼此類化合物的基因往往只在特定環境下才表達,常規培養條件下處于沉默狀態或表達量低,難以分離純化和投入工業生產。統計數據顯示微生物中表達并分離出對應代謝產物的合成基因只占總數的10%左右,微生物資源在新藥研發領域的潛力遠遠沒有被開發出來,還需要不斷地進行探索[9]。基因組挖掘是指通過高通量測序技術對生物信息學預測的可能編碼高價值天然產物的基因進行測序,了解基因表達翻譯的控制機制并通過一系列手段將其激活表達,分離純化出相應目的產物后解析結構并進行生物活性測試的技術。與傳統方法相比它將整個過程簡化為編碼次級代謝產物基因序列數據在培養基上的變現及分離柱上的純化,讓整個流程處于分子生物學的指導[10],研究主要包含以下方向。
1 基因組挖掘
1.1 傳統基因組學挖掘
基因組學挖掘天然產物的關鍵在于尋找到有價值的合成基因,酶作為生物體內代謝活動的催化劑在次級代謝產物合成途徑中發揮了重要作用,催化同一類型反應的酶系在分子結構上具有保守性,核心往往有一個高度重復的氨基酸序列,直接以負責目的代謝產物的合成酶作為靶點,系統進化樹分析含有關聯序列的酶基因,設計交聯探針或PCR引物即可定向選擇具有合成潛力的基因。最典型的就是目前研究比較深入的聚酮化合物合酶(Polyketidesynthase,PKS)和非核糖體肽合酶(Nonribsomal peptide synthetases,NRPS), 其 分子結構是由多個結構單元組成,線性排列的負責相應結構單元的模塊化合成酶基因組成了完整的生物合成基因簇。Sun等[11]對海洋黏細菌Haliangium ochraceum SMP-2進行基因組挖掘,發現了一種聚酮化合物-非核糖體肽雜合化合物Haliamide 1(圖1),通過核磁共振和HR-MS對結構進行表征和喂養實驗發現其生物合成的結構單元為苯甲氨酸鹽、丙氨酸和丙氨酸鹽,從而建立了鹵胺合成的生物模型,活性檢測發現對Hela-S3細胞具有毒性。
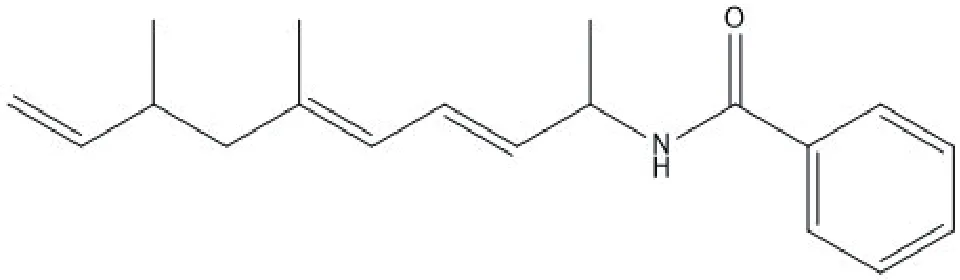
圖1 Haliamide 1結構
1.2 比較基因組學挖掘
比較基因組學是基于基因組圖譜和測序技術,對已知基因和基因組結構進行比較,了解基因功能、表達機理、物種進化的科學。在研究目的蛋白基因的同時,關注完整的基因簇或基因鄰域,分析不同分子間相關性來挖掘未知酶系和其體內的生物合成機制,促進新化合物的發現。將已知基因序列的功能基因簇通過NCBI數據庫與其它已測序的生物遺傳圖譜進行局部共線性比對驗證是否具有類似的合成路徑,挖掘已知物種未發現的合成潛力。通過將編碼目標修飾蛋白的抗藥性基因與未表征次級代謝合成基因簇孤兒BGCs相關聯,基因組定向挖掘的過程中,可預測路徑特異性小分子的生物學功能。Tang等[12]對86個鹽孢菌基因組的全基因組查詢,獲得與天然產物BGCs共定位的管家基因,篩選出一個可能具有脂肪酸合成酶抗性基因的PKS-NRPS雜合BGC(TLM),從而推測出硫代四酸類抗生素的合成途徑。
1.3 宏基因組學
宏基因組是指環境中所有微生物遺傳物質的總和,包含可培養和未可培養微生物遺傳物質基因,通過構建宏基因組文庫,繞過培養難點直接在基因層次研究未可培養微生物的合成潛力,可能會發現新的耐藥基因和抗生素機制,完善藥物靶點,以及鑒定在不同生長環境下對細菌、真菌生長起重要作用的基因,從而豐富基因庫來源[13]。操作包括從環境中提取樣品DNA,高通量測序后選擇合適的載體拼接修飾,導入宿主細胞激活表達,實現對基因組網絡的功能驗證與開發利用。Levin等[14]通過結合基因組學和生物學知識,將合成具有保守氨基酸結構的蛋白質基因和正常人體的消化道菌群數據進行對比,分析了甘氨酰自由基酶(GRE)家族,以甘氨酸為中心用自由基進行化學反應,確定了一個新的在生產L-脯氨酸通路上起重要作用的反式 4 羥基脯氨酸脫水酶,它在所有腸道微生物宏基因組樣本中都能發現,從而揭示了反式 4 羥基 L 脯氨酸的常規代謝途徑。
2 沉默基因的表達
通過基因組挖掘的方法能夠準確發現大量與合成次級代謝產物相關的基因簇,然而多數基因在常規培養條件下處于沉默狀態,必須在特定生理環境下或外來細菌病毒入侵引起免疫應答機制時才會激活。這些基因由于常規條件下不表達,因而更有可能在人為控制下激活產生骨架新穎、生物活性高的次級代謝產物,如何選用適當的方法激活這些“沉默代謝途徑”是繼基因組挖掘后的重要研究內容,也是最終獲得潛在藥物資源的關鍵。現階段激活沉默基因的策略如下。
2.1 改變培養條件、共培養和化學誘導
通過改善培養基組分、溫度、pH、光照條件、氣壓、氧濃度、微量元素,或與合適病原菌、競爭菌共培養,提高生存壓力刺激沉默基因表達。某些能夠改變微生物表觀遺傳途徑的小分子物質,如DNA甲基轉移酶抑制劑、組蛋白去乙酰化抑制劑能夠影響DNA甲基化、組蛋白修飾和RNA干擾等從而影響微生物對于次級代謝產物的合成調控,誘導沉默基因表達。Bode等[15]研究小組通過改變培養條件從6個微生物中分離出超過25個結構類型的100多個化合物;Scherlach等[16]發現Aspergillusnidulans這種產生在水稻培養基但不產生在其它介質中的新型喹諾林類生物堿;Shwab等[17]發現編碼黑曲霉組蛋白去乙酰化酶(HDAC)的hdaA缺失,導致兩個端粒近端基因簇的轉錄激活,隨后相應的分子(毒素和抗生素)水平增加。
2.2 調控基因改造
次級代謝產物合成基因簇是復雜的,它不只是單純含有合成化合物的基因,同時還包含大量參與基因表達,控制代謝途徑的調節基因。通過設計方案對調節基因進行敲除、阻遏、或使其過表達,都會影響次級代謝產物的合成效率。選擇性過表達正調控因子和降低負調控因子表達減少反饋抑制可促進基因的激活和代謝產物在發酵液中的積累[18]。Laureti等[19]過表達Streptomyces ambofaciens中的特異性調節基因Sam R0484激活了一個沉默的聚酮合酶的次級代謝基因簇表達,最終得到了新型的具有抗腫瘤活性的化合物Stambomycins。Molloy等[20]通過滅活推測的TetR轉錄抑制因子 arPRII等手段對編碼化合物基因進行鑒定,質譜和核磁共振光譜對產物進行結構表征證實其是新的生物堿類化合物,生物活性測試發現對于黏菌的生長和菌落分化具有負面作用,其中一些抗菌活性較弱。
2.3 核糖體工程
與微生物在營養缺陷時的應激機制相似,核糖體工程就是通過以不同的微生物抗性突變為靶標,高效獲得次級代謝產物合成能力得到提高的突變株的推理育種新方法。核糖體不僅是微生物體內蛋白質合成位點,還可以與藥物結合抑制蛋白質合成從而調控次級代謝產物的合成途徑,如鏈霉素和氯霉素可分別與核糖體30S亞基和核糖體50S亞基結合。在微生物的穩定生長期,與合成次級代謝產物相關基因的大量表達取決于此時的核糖體功能。以生物體中核糖體蛋白、RNA聚合酶RNAP、轉錄因子為作用靶點,通過在核糖體蛋白編碼基因上引入定點修飾和突變,改造核糖體結構來改變相關催化酶的生物學性質,可能會激活相關沉默的抗生素合成基因,影響代謝途徑[21-23]。如 Dong等[24]通過核糖體技術使海洋真菌雜色曲霉菌ZBY3獲得耐新霉素抗性后發現6種新型抗腫瘤物質;Yi等[25]通過同樣手段在紫草青霉G59中分離到一種新的環戊酮硫化物。
2.4 異源表達與強啟動子替換
異源表達一般指異源蛋白表達,對于具有復雜調控機制或不可培養微生物的基因,將目的合成基因簇整合插入到質粒、黏粒、人工染色體等載體后在異源宿主內表達。該方法可繞過自身復雜的調控機制,減少原有產物含量積累造成的反饋抑制和對特定表達環境的需求,對生長緩慢的菌種可明顯降低生產周期。目前用于挖掘新化合物的異源表達流程通常為:大腸桿菌擴增載體、酵母細胞對目的片段進行拼接、曲霉作為宿主細胞對目的基因進行表達[26]。啟動子是位于起始位點上游能增加或降低基因轉錄速率,被RNA聚合酶識別、結合和開始轉錄的一段DNA序列,它含有RNA聚合酶特異性結合和轉錄起始所需的保守序列,本身不被轉錄。在沉默基因簇前添加或替換誘導型或組成型啟動子,構建新的調控機制,通常可刺激基因激活提高表達量,利于最終工業生產,常見的有構巢曲霉A的持家啟動子gpdA,黑曲霉作為宿主異源表達時的常用強啟動子glaA等[27-30]。Wang等[31]發現在工程大腸桿菌中表達編碼嗜熱鏈球菌雙功能谷胱甘肽合成酶基因GshF,分批發酵分離積累的GSH達到以往表達最高水平。黃開華等[32]利用蛹蟲草自身強啟動子,構建天蠶菌素抗菌肽過表達載體,介導農桿菌遺傳轉化系統,在蛹蟲草中成功表達,活性測定和抑菌實驗表明相較于野生型其生物學活性明顯提高。Saha等[33]通過啟動子工程和異源表達對海洋鏈霉菌SCSIO02999中ptm基因簇進行激活,發現6種新的多環四酸酯大環酰胺PTMS,其對人癌細胞株具有較強細胞毒活性,體內基因破壞實驗和體外生化實驗表明多循環形成一個還原環化級聯,并證明PtmC是雙功能環化酶,可催化伊卡毒素內五元環形成。
3 真菌基因組挖掘進展
來源于真菌的天然產物具有廣泛的生物活性,然而其遺傳調控途徑相對細菌復雜,導致現階段對其研究較少。隨著高通量測序技術和生物信息學發展,對復雜微生物代謝通路進行分析的技術逐漸成熟,真菌作為尋找新穎骨架結構和獨特作用機制藥物的重要來源,其生物醫藥價值受到眾多科學家的關注。真菌中次級代謝產物主要分為:萜類化合物、聚酮類化合物、生物堿、非核糖體肽、氨基酸衍生物、氨基酸/聚酮雜合體細胞松弛素等。目前已從真菌中獲得的藥物包括具有抑菌、抗腫瘤、抗炎和免疫調節功能的核苷酸類抗菌素蟲草菌素;藥用真菌靈芝中分離的抗氧化抗衰老、促進核酸蛋白質代謝的靈芝多糖以及最經典的青霉素(圖2-A)和頭孢菌素(圖2-B)等,研究還發現紅豆杉內生真菌會合成毒性較小的廣譜抗腫瘤藥物紫杉醇(圖2-C)[34]。絕大多數真菌種類屬于尚未認知狀態,近幾年對于真菌挖掘潛在藥物又有了新進展[35]。
Shu等[36]從青霉Penicilin Herquei中分離到新的重氮雜環辛烷衍生物和新戊二酮衍生物,進行生物活性測試發現具有抗丙型肝炎病毒(Human antiheptitis C virus,HCV)能力,這是首次報道具有抗HCV能力的重氮環辛烷衍生物(3-A)。Chiba等[37]在篩選新的抗流感病毒時,從真菌FKI-7215發酵液中分離出草甘氨酸A(Herquline A)(圖3-C),其對于流感病毒復制有劑量依賴性的抑制作用且對病毒神經氨酸酶沒有抑制作用。Tansakul等[38]從土壤真菌黑曲霉(Penicillium herqueiPSURSPG93)中分離出新的苯丙烯酮衍生物和五個已知的phenalenone衍生物,其一個phnalenone衍生物對非洲綠猴腎成纖維細胞無細胞毒性,但抗氧化能力弱。Arunpanichlert等[39]在海草源真菌Pestalotiopsp中分離出4種新的代謝產物,包括2種巰基萜類、1種異香豆素和1種苯酚,提出了一種多烯類化合物的生物合成途徑并有較好的抗菌、抗瘧和細胞毒活性。Klaiklay等[40]從紅樹林真菌Pestalotitisp中分離到4種新的聯苯醚、3種新的色酮、1種新的黃酮、1種新的花椒酮、1種新的丁烯內酯-Pestrolide(圖3-D)和11種已知化合物。光譜技術確定了它們的結構。Pestalolide對白色念珠菌和新隱球菌具有較好的抗真菌活性。Uchoa等[41]通過改變培養條件,從巴西海岸篩選的一株黑曲霉中發現具有新的氮化骨架的天然呋喃衍生物(圖3-B),其對HCT-116細胞系具有毒活性。隨著研究的深入,真菌作為具有重大潛在價值的自然資源寶庫,在研究新型藥物和開發藥物先導化合物過程中將發揮重要作用。
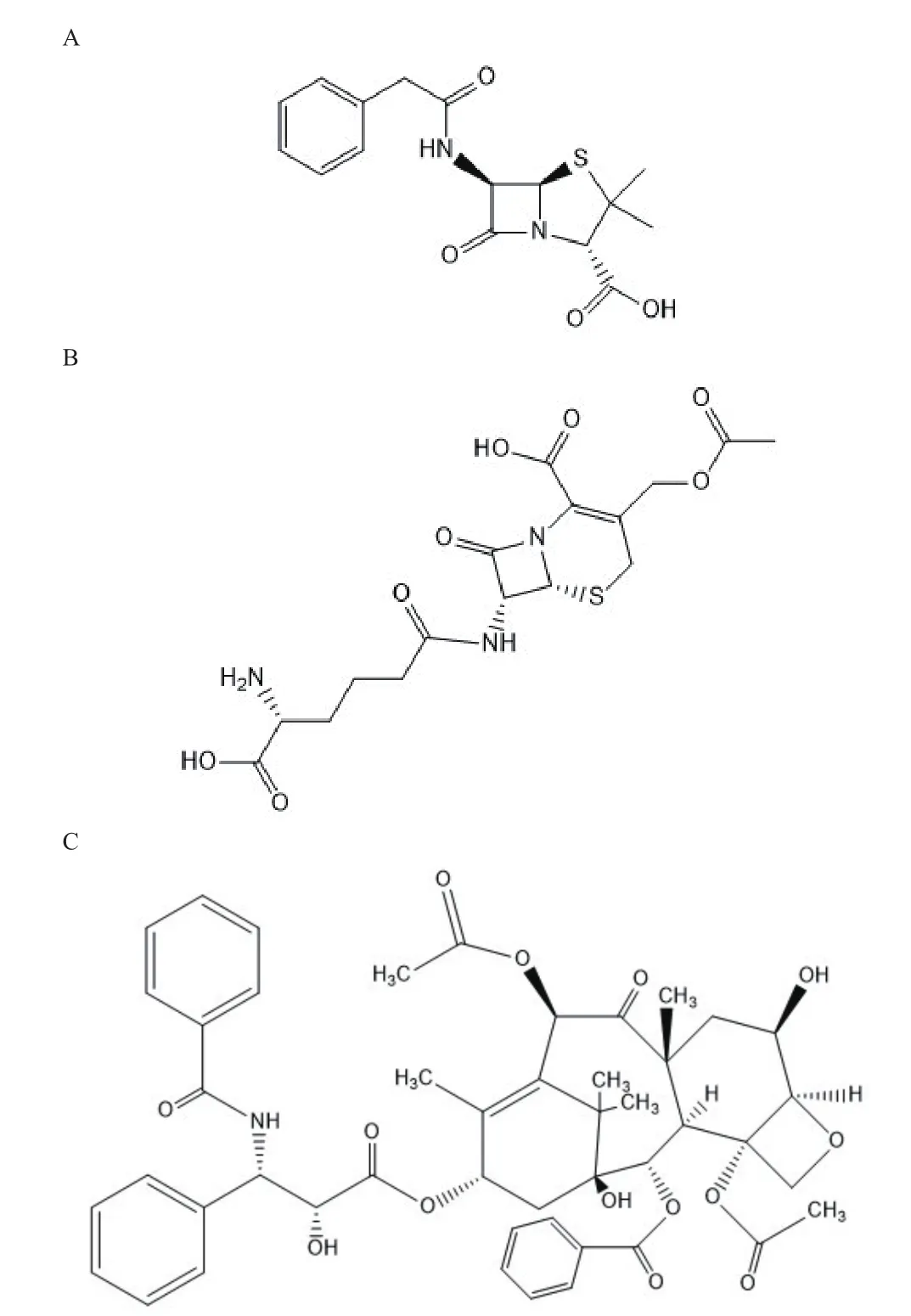
圖2 青霉素(A)、頭孢菌素C(B)、紫杉醇(C)結構
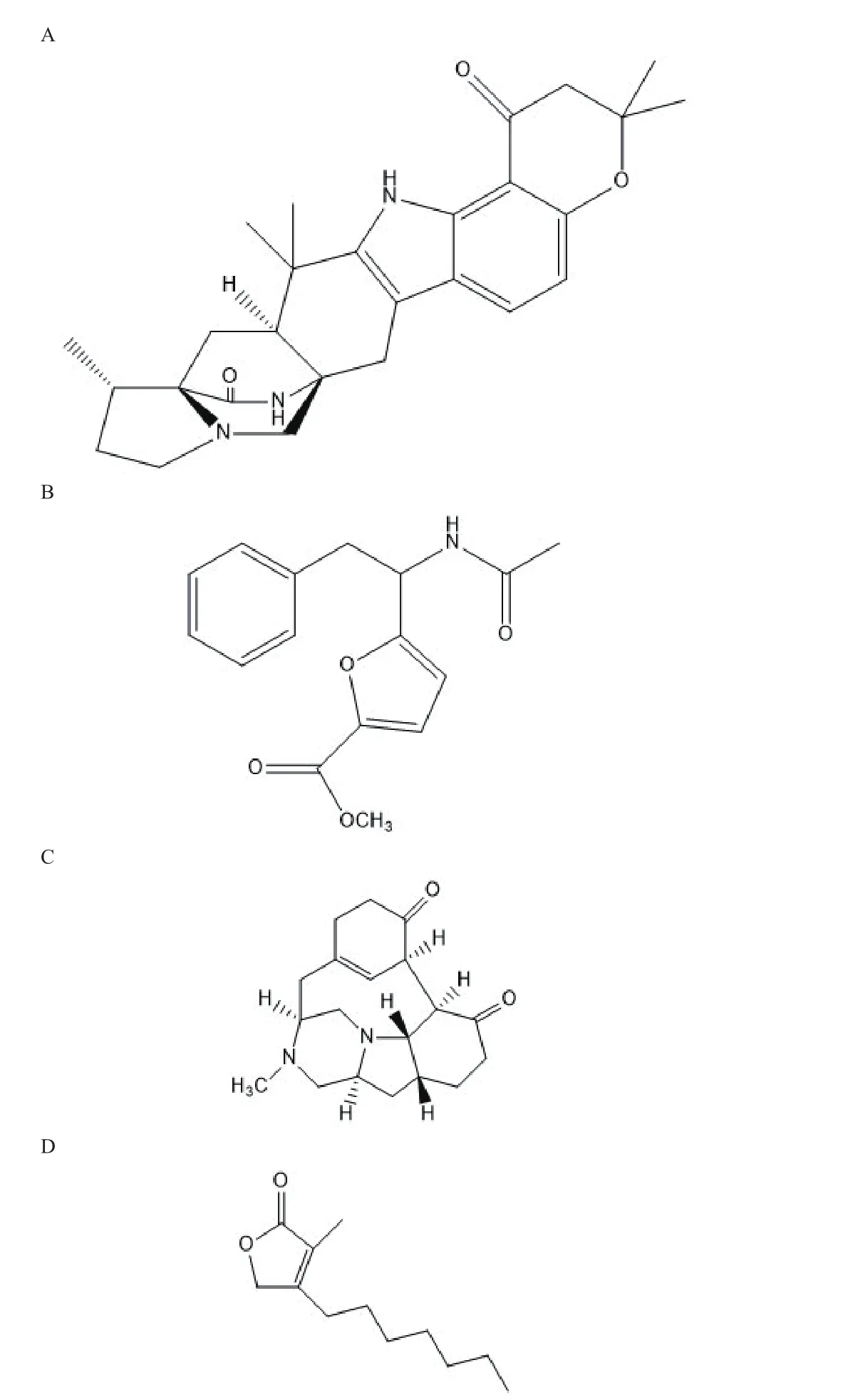
圖 3 抗HCV(A)、抗HCT-116(B)、Herquline A(C)、Pestrolide(D)結構
4 展望
分子生物學技術的發展,使人們對于生物體遺傳基因、代謝調控機制的了解愈加深刻,也越來越渴望通過基因水平定向尋找與實際需求相符的藥物,尤其近幾年計算機和數學等工具發展帶來的強大基因圖譜分析能力,更是為此提供強大的助力。隨著大量微生物遺傳圖譜公布,潛在的合成基因簇數據不斷積累,最終對于天然活性產物的挖掘將會得到井噴式發展,具有有效藥用價值的化合物也會越來越多,真菌由于自身特性也必將扮演重要的角色。盡管現如今對于微生物資源利用才剛剛起步,尚沒有完善的依托基因組挖掘技術開發藥物的流程,但可以預計,通過誘導激活沉默基因簇充分挖掘生物合成潛力,將成為未來新藥研發的重要手段,也會吸引更多國內外科研工作者的興趣。
猜你喜歡
天天愛科學(2022年9期)2022-09-15 01:12:54
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
科學大眾(2021年9期)2021-07-16 07:02:54
軍事文摘(2020年20期)2020-11-28 11:42:50
航空世界(2020年10期)2020-01-19 14:36:20
小學科學(學生版)(2018年3期)2018-04-18 12:34:19
知識經濟·中國直銷(2017年10期)2017-11-07 02:39:52
知識經濟·中國直銷(2017年3期)2017-04-16 03:08:01
文學少年(有聲彩繪)(2017年2期)2017-03-06 08:10:54
