藍色花形成的基因工程進展與育種策略
2019-11-21 11:09:26李崇暉尹俊梅
生物技術通報 2019年11期
李崇暉 尹俊梅
(1. 中國熱帶農業科學院熱帶作物品種資源研究所 農業部華南作物基因資源與種質創制重點實驗室,海口 571101;2. 中國熱帶農業科學院海口實驗站,海口 571101;3. 海南省熱帶觀賞植物種質創新利用工程技術研究中心,儋州 571737)
花色是觀賞植物最受關注的性狀,新奇的花色是育種者始終追求的目標。長期以來雜交育種和突變選種是創造新花色的主要方法。花青素是花瓣中普遍存在的花色素,能使花呈現從紅到紫紅到藍的變化,花青素在胞質中合成,在液泡中積累[1]。在高等植物中花青素的生物合成從查爾酮開始到生成花青素3-葡萄糖苷的途徑非常保守(圖1),且研究得比較透徹[1]。根據對不同觀賞植物花青素生物合成途徑的認知,每個物種表達特定的酶基因,只能積累有限種類的花青素苷,因此一個物種擁有全部種類的花色幾乎是不可能的。例如,大宗貿易花卉月季、康乃馨和菊花均因不積累飛燕草素為核心的花青素苷而缺少紫色和藍色的品種。培育藍色名貴花卉是幾輩育種者多年的夢想,基因工程技術為花色育種者打開了一扇大門,使突破物種遺傳背景限制,培育新奇的花色成為可能。藍色花的育種一直是花色研究的熱點,近年來獲得了大量的研究成果。這期間最突出的成就是月季、菊花、蝴蝶蘭等重要花卉藍色品種的誕生[2-4]。本文根據已有研究基礎[5-6]總結了藍色花形成的原因以及近15年藍色花基因工程育種的基本策略及研究進展。
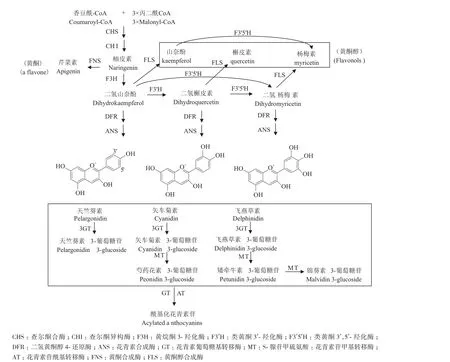
圖1 植物花青素苷生物合成途徑簡圖[2]
1 藍色花形成的化學和生理基礎
1.1 藍色花色素
藍色花最主要的成因在于花青素的化學結構,藍色花瓣中含有的花青素通常被甲基化、糖苷化和酰基化修飾,被芳香酸如對-香豆酸、咖啡酸、阿魏酸、對-羥基苯甲酸等多重酰基化是藍色花中最主要的花青素苷酰基化修飾,芳香酸酰基與花青素形成“三明治”型的分子內堆疊,這使得花青素苷藍化且更加穩定[7-8]。酰基化修飾的位置也非常重要,通常在花青素3'-和/或7-位上發生芳香酸酰基化比3-和/或5-位上的芳香酸酰基化對于藍色的形成更為重要[9]。大多數藍色花主要含有飛燕草素糖苷及其酰基化衍生物,極少數的藍色花以酰基化的矢車菊素糖苷或矮牽牛素糖苷為主。以飛燕草素為主的藍色花典型代表有鴨跖草(Commelina communis)、飛燕草(Delphiniumspp.)、 龍 膽(Gentianaspp.)、鼠尾草(Salviaspp.)、蝶豆花(Clitoria ternatea)、風信子(Hyacinthus orientalis)等[7,10-11]。以酰基化矢車菊素糖苷為主的藍色花有矢車菊(Centaureacyanus)、圓葉牽牛(Ipomoea purpurea)、 綠絨蒿(Meconopsisspp.)等[7,12-14]。以酰基化矮牽牛素糖苷為主的藍色花以琉璃唐草(Nemophila menziesii)為代表[7,15]。
鴨跖草、矢車菊、鼠尾草、琉璃唐草的藍色色素為金屬絡合花青素苷(Metalloanthocyanins),即花青素苷、黃酮和金屬離子以6∶6∶2的化學計量比自絡合形成的超分子金屬絡合色素[7]。在以飛燕草素為發色團的超分子金屬絡合色素中,Mg2+對穩定藍色的作用是足夠的;在以矢車菊素或矮牽牛素為發色團的金屬絡合色素中,除了Mg2+以外,還需要1/6當量的Fe3+輔助[7]。然而,自然界中含有金屬絡合花青素苷的藍色花卻非常少。通常的藍色花中以非化學計量比存在的金屬離子對花青素苷起到輔助著色的作用,這些藍色色素穩定性較差,在分離色素或結晶的過程中藍色便消失了[7]。例如,大花綠絨蒿(Meconopsis grandis)的色素組成為1當量的矢車菊素糖苷,2或更多當量的山奈酚衍生物(黃酮醇),1/6當量的Fe離子和過量的Mg2+[16]。
1.2 藍色呈現所需要的液泡環境
許多體外試驗表明,有了藍色花青素如飛燕草素及其衍生物通常不一定呈現藍色,可見液泡內的環境包括pH值、有機物和無機物的組成和含量等對藍色花的形成尤為重要。以繡球花為例,紅色、紫紅色、藍色等花中均只含有飛燕草素3-葡萄糖苷。進一步研究發現,藍色品種花瓣的pH值、5-O-乙酰奎寧酸與花青素苷的摩爾比率,以及Al3+的摩爾當量都顯著高于紅色細胞[17-18]。郁金香品種Murasakizuisho內層花瓣基部因大量積累鐵離子,與飛燕草素3-蕓香糖苷作用而呈藍色[19]。牽牛花(Ipomoea tricolorcv. Heavenly Blue)在花蕾期為紫紅色,完全開放為藍色,其所含色素為紫紅色的咖啡酸酰化芍藥花素糖苷,沒有金屬離子絡合。在花蕾期液泡pH值為6.6,完全開放期液泡pH值上升為7.7[20]。
目前藍色花形成的化學和生理機制尚未完全解析,根據已有研究結果,藍色花形成的因素主要有:(1)藍色花青素的生成;(2)花青素苷的多重芳香酸酰化;(3)黃酮、黃酮醇等輔助色素參與;(4)金屬離子絡合;(5)較高的液泡pH值等。
2 與藍色花形成相關的基因
2.1 與藍色色素合成相關的酶基因
在高等植物中,從查爾酮到花青素-3-葡萄糖苷的生物合成途徑非常保守,從花青素-3-葡萄糖苷之后的進一步糖苷化、酰基化修飾方式在不同植物中變化多樣,因此藍色花的花青素苷合成途徑尚未完全闡明。已知的幾種參與藍色花青素合成的關鍵酶中,細胞色素P450氧化酶家族 CYP75A或CYP75B亞家族的類黃酮3',5'-羥化酶(F3'5'H)催化二氫山奈酚B環上的3'-和5'-位的羥基化生成二氫楊梅素,是合成飛燕草素的前體。由于單一積累飛燕草素糖苷及其多酰基化的衍生物被育種者認為是培育藍色花最有效的手段,因此F3'5'H基因最受關注,并已從多種植物中克隆并鑒定[21]。而抑制藍色花自身的F3'5'H基因表達,可導致花色變成紫紅色,如龍膽[22]。
二氫黃酮醇4-還原酶(DFR)催化二氫黃酮醇生成無色花青素苷元(Leucoanthocyanidins)。不同植物中DFR的底物專一性影響了花瓣中最終積累的花青素苷元B環上的羥基化程度。例如,矮牽牛、大花蕙蘭等的DFR只能催化二氫槲皮素和二氫楊梅素,不能催化二氫山奈酚[9]。因此這些植物的DFR對于專一性地合成飛燕草素是有利的。
花青素的糖苷化和酰基化修飾對于維持其顏色的穩定非常重要,也是藍色花色素最常見的結構修飾方式。迄今鑒定出的花青素糖苷化酶有兩類,一類是依賴UDP-糖的糖苷化酶家族1,另一類是依賴酰基-葡萄糖的花青素糖苷化酶[9]。已鑒定的花青素苷酰基轉移酶也分為兩類,一類是利用酰基-CoA為供體,將酰基轉移至花青素苷糖基部分的特定位置;另一類是類絲氨酸羧肽酶(SCPL)-酰基轉移酶,利用酰基葡萄糖作為供體分子,催化花青素苷糖基部分的酰基化[9]。飛燕草的糖基轉移酶基因AA7GT或酰基轉移酶基因AA7GT-AT發生突變,均可產生由于積累中間產物而導致粉色或者紅色的花色表型[23]。
2.2 細胞pH值調控基因
矮牽牛花瓣中鑒定出7個調控pH值的基因位點PH1-PH7[24]。其中PH5編碼一種定位于液泡膜的H+P3A-ATPase質子泵。當PH5基因發生突變時,液泡中的pH值升高,花瓣從紫紅色變成藍紫色[25]。PH1基因突變體的花色與PH5基因突變體顏色相似,PH1編碼的P3B-ATPase可與PH5編碼的H+P3AATPase物理性結合,促進H+轉運活性,增加液泡的酸化程度[26]。裂葉牽牛(Ipomoea nil)開花過程中InNHX1和InNHX2基因編碼的轉運蛋白將K+/Na+轉運至液泡,提高液泡pH值,使花色變藍[27-28]。
2.3 金屬離子轉運蛋白基因
從郁金香中分離鑒定了一種編碼液泡鐵離子轉運蛋白的基因TgVit1,其編碼蛋白具有促進液泡中鐵離子的積累,與飛燕草素3-蕓香糖苷作用呈現藍色的功能[29]。隨后在矢車菊中鑒定了與TgVit1同源的基因CCViT[30]。繡球花中分離鑒定了編碼液泡膜和質膜定位的Al離子轉運蛋白的基因VALT和編碼質膜Al離子轉運蛋白1的基因PALT1,這些基因編碼的Al離子轉運蛋白與Al離子參與藍色繡球花花色的形成有關[31]。
3 藍色花卉基因工程育種實踐與策略
目前,育種者利用基因工程技術培育藍色花主要有以下幾方面的實踐(表1)。
3.1 人工創造飛燕草素合成途徑
盡管矢車菊、綠絨蒿等藍色花是由矢車菊素糖苷與金屬離子形成絡合物而呈藍色,含有芍藥花素糖苷的牽牛花由于細胞內pH值升高而產生藍色,但是,大多數藍色花的成因是積累大量的飛燕草素。而許多缺少藍色花色的植物如康乃馨、月季、菊花、百合、蝴蝶蘭等[21,32]就是因為本身缺乏飛燕草素合成途徑上的關鍵基因F3'5'H。因此通過轉基因技術,向不含飛燕草素的花卉植物中,轉入外源F3'5'H基因促進飛燕草素的生成,是全世界公認的最有可能獲得藍色花的手段。
3.1.1 轉基因受體的篩選原則 轉基因受體要有合適的遺傳背景和類黃酮組成[4]。轉入外源F3'5'H后,能否將F3'5'H酶的產物二氫楊梅素轉化成飛燕草素非常關鍵,即首先要檢測轉基因受體DFR酶是否具有催化二氫楊梅素能力,如沒有則要轉入外源的DFR基因。還要考慮受體植物內源的DFR、F3'H和FLS酶對F3'5'H酶底物的競爭。要避免底物競爭,最好有這些競爭基因功能缺失的天然突變體,或者通過基因工程手段如RNAi等下調這些基因的表達。此外,盡量選擇花瓣pH值較高、黃酮/黃酮醇含量較高的材料[21]。這樣才有可能獲得較理想的藍色花表型。
3.1.2 單一轉入外源F3'5'H基因 國內外學者對轉入外源F3'5'H基因做了大量的嘗試。將洋桔梗(Eustoma grandiflorum)F3'5'H基因轉入煙草,轉基因煙草最多僅積累了23%的3',5'-羥基化的花青素苷,花色有所加深[33]。Tanaka[21]將三色堇(Violasp. cultivar black pansy)F3'5'H基因轉入后使月季最多產生了60%的飛燕草素糖苷,花色變成深紅色。堇菜(Violaspp.)F3'5'H基因在月季品種Lavande中過表達,最多可產生44.2%的飛燕草素,然而有三分之二的轉基因株系的飛燕草素含量少于20%,花色變化不明顯[4]。蔓長春花(Vinca major)F3'5'H基因VmFH1在紅色矮牽牛中過表達,產生了深紅色帶深紫色扇形區域的花色表型[34]。風鈴草(Campanula medium)F3'5'H基因在煙草中異源表達可以特異性積累飛燕草素(最多達99%),花色也變成新奇的紫色[35]。蝶豆花F3'5'H基因轉入馬鞭草植株后,比過表達馬鞭草F3'5'H基因的植株產生的表型變化更明顯,花色由粉紅色變成紫紅色[36]。將蝴蝶蘭(Phalaenopsis)F3'5'H基因在矮牽牛花中過表達,使花色由粉色變成深粉色;蝴蝶蘭F3'5'H基因在東方百合Sorbonne花被片中瞬時過表達,產生了紫色的細胞[37]。日本Florigene公司將三色堇(Violasp. cultivar black pansy)F3'5'H基因在非洲菊中過表達,能夠積累50%的飛燕草素糖苷,花色也發生了一定的變化[8]。日本千葉大學Mii教授等[6]將鴨跖草(Commelina communis)F3'5'H基因轉入大麗花,轉基因植株產生了飛燕草素衍生物,花色呈現藍紫色[6,32]。Mii教授等還將鴨跖草F3'5'H基因轉入了蝴蝶蘭,由于粉色蝴蝶蘭花瓣中主要花青素為7-位和3'-位多重糖苷化和芳香酸酰化的矢車菊素,因此單獨過表達矢車菊F3'5'H基因的蝴蝶蘭很容易生成多重酰基化的飛燕草素,呈現出非常接近藍色的花色[6,32]。
Brugliera等[38]將 三 色 堇(Violasp. cultivar black pansy)F3'5'H基因在菊花中過表達,產生40%的飛燕草素糖苷,使花瓣顏色轉藍,然而所產生的花色并不新奇,已有育種者用傳統手段育成類似的花色。Noda等[39]篩選出菊花中高效驅動外源F3'5'H表達的啟動子為菊花F3H(CmF3H)啟動子,在此啟動子驅動下,帶有煙草NtADH基因5'UTR序列的風鈴草F3'5'H基因過表達能產生更多的飛燕草素(達到95%),花色也呈現出了藍紫色。
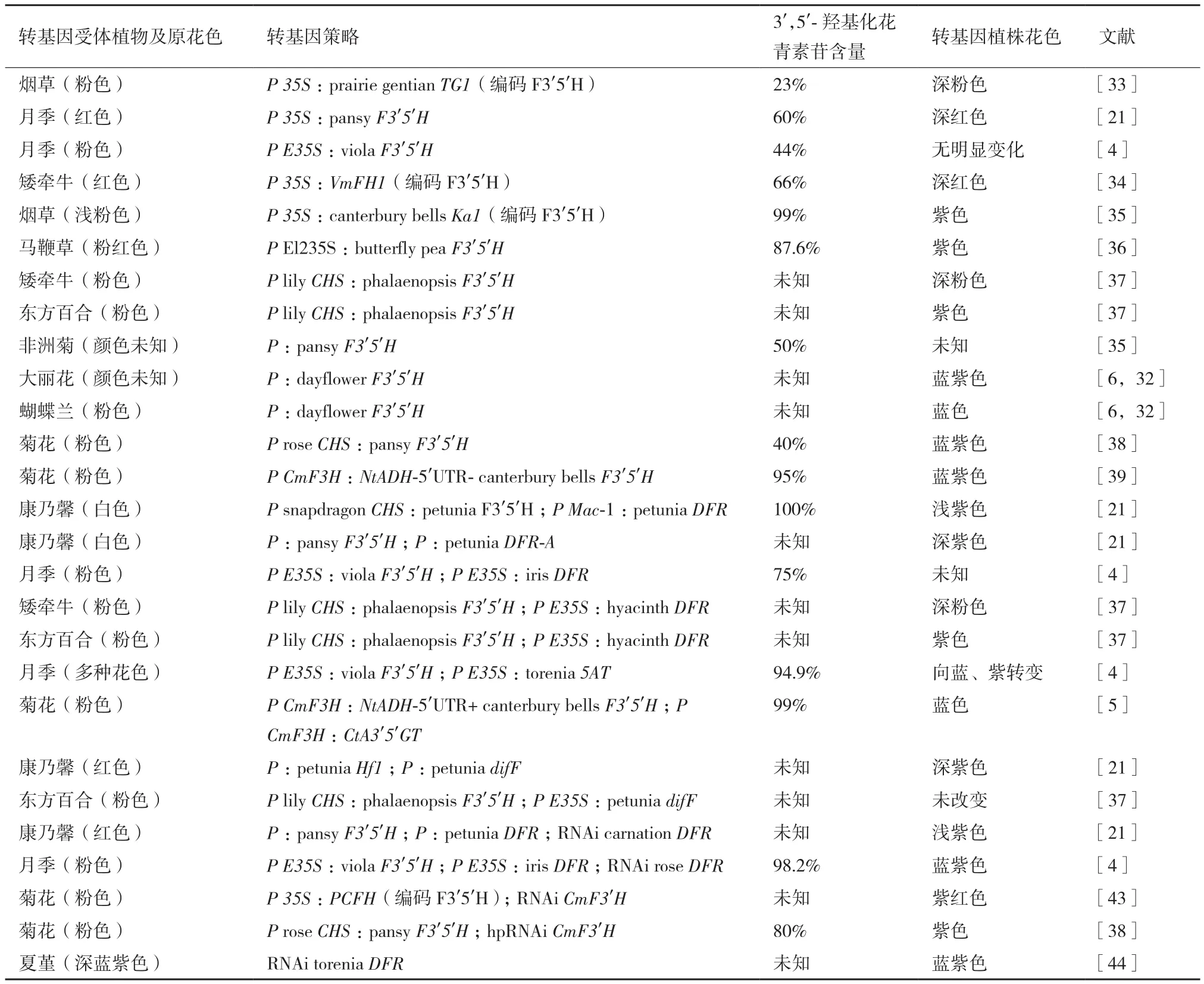
表1 已有的培育藍色花轉基因育種策略及獲得的花色表型
以上結果表明,F3'5'H基因的來源以及其轉化的受體對改變花色的結果都非常重要,因此需要篩選適合受體植物的外源基因和啟動子。然而通常僅轉入外源F3'5'H基因雖然能在目標植物花瓣中產生飛燕草素,卻往往并不能達到令人滿意的藍色效果。
3.1.3 外源F3'5'H基因與其它關鍵酶基因同時轉入
3.1.3.1 與DFR基因同時轉入 將外源F3'5'H基因和DFR基因同時轉入創造藍色花最經典的實例就是藍紫色的“Moon”系列康乃馨的誕生。以散枝型DFR和F3'H突變體白花品種White Unesco為轉基因受體,轉入矮牽牛F3'5'H基因和DFR基因,獲得了完全積累飛燕草素的淺紫色品種FLORIGENE Moondust,這是世界上首個商業化的轉基因觀賞植物[21]。隨后發現三色堇(Violasp. cultivar black pansy)的F3'5'H基因在康乃馨中表現更好,將其與矮牽牛DFR-A基因同時轉化DFR突變體品種Unesco,產生了比Moondust積累更多飛燕草素的深紫色品種FLORIGENE Moonshadow,其花瓣中含有一種芹菜素C-苷作為輔助色素,且花瓣的pH值更高,使花色看起來更藍[40]。采用同樣的策略育出了 FLORIGENE Moonvista、FLORIGENE Moonshade、FLORIGENE Moonlite、FLORIGENE Moonaqua等 眾多商業品種[21]。
月季品種Lavande中,同時過表達堇菜(Violaspp.)F3'5'H基因和鳶尾(Iris×hollandica)DFR基因比單獨過表達堇菜F3'5'H基因積累了更多的飛燕草素,最高可達75%[4]。蝴蝶蘭F3'5'H基因與風信子(Hyacinthus orientalis'Sky Jacket')DFR基因同時在矮牽牛中過表達比蝴蝶蘭F3'5'H基因單獨過表達能積累更多的飛燕草素糖苷,在東方百合花被片中瞬時過表達這兩個基因比單獨過表達F3'5'H基因產生顏色更紫的細胞[37]。
3.1.3.2 與花青素苷修飾酶基因同時轉入 除了改變受體植物的花青素苷元結構生成飛燕草素外,學者們也對飛燕草素糖苷的芳香酰基化修飾進行了嘗試。將堇菜(Violaspp.)F3'5'H基因和夏堇(Torenia hybridcultivar Summer Wave Blue)5AT(花青素苷5-羥基肉桂酸酰基轉移酶)基因共同轉化不同花色的月季品種,結果顯示各品種轉基因株系都不同程度地積累了飛燕草素糖苷,花色也發生了不同程度的藍化。只有一些(不超過44%)花青素苷被夏堇的5AT酶酰基化,且5-位酰基化只能使花青素苷的最大吸收波長紅移4 nm,因此很難觀察到由酰基化產生的花色變化[4]。
由于單獨轉化風鈴草F3'5'H基因得到的菊花花色并不是純正的藍色[39],為了讓菊花能像蝶豆花一樣積累3'-位和5'-位被糖苷化的飛燕草素糖苷,研究者進一步將蝶豆花的UDP-葡萄糖:花青素苷3',5'-O-葡萄糖基轉移酶基因(CtA3'5'GT)與風鈴草F3'5'H基因同時轉入菊花,成功生成了飛燕草素3',5'-雙糖苷化的衍生物,在木犀草素7-丙二酰葡萄糖苷輔助著色作用下,使花瓣呈現了純正的藍色[5]。應用這種方法實現了各種花型藍色菊花的培育[32]。
3.1.3.3 與輔助基因同時轉入 矮牽牛的一種在花中特異表達的基因difF,因其編碼一種細胞色素b5蛋白能夠增強F3'5'H酶活性[41],在藍色花育種中可作為F3'5'H基因的輔助基因,同時轉化目標植物。FLORIGENE公司向缺少F3'H酶活性積累天竺葵素的紅色康乃馨品種Cerise Westpearl中,同時轉入矮牽牛F3'5'H基因(Hf1)和difF,獲得了深紫色的品種FLORIGENE Moonvelvet在美國上市[21]。在東方百合中同時瞬時表達蝴蝶蘭F3'5'H基因和矮牽牛difF基因,然而被轉化細胞顏色沒有發生變化[37]。
3.1.4 轉入外源基因同時抑制內源關鍵酶基因 FLORIGENE公司利用RNAi技術抑制康乃馨紅色品種Cerise Westpearl內源DFR基因表達,并轉入三色堇(Violasp. cultivar black pansy)F3'5'H基因和矮牽牛DFR基因,產生了紫色品種FLORIGENE Moonberry和淺紫色品種FLORIGENE Moonpearl,均已上市[21]。
抑制月季品種Lavande的DFR基因,同時轉入堇菜(Violaspp.)F3'5'H基因和鳶尾(Iris×hollandica)DFR基因,2/3的轉基因株系中積累超過80%的飛燕草素,最高可達98%,成功地使月季中花青素代謝流從矢車菊素合成途徑轉向了飛燕草素合成途徑,花色由淺紫紅色變成了藍紫色[4]。至此藍色月季Applause誕生了,并于2009年上市[21]。然而Applause花色還是偏向紫色,接下來將從增加輔助色素和調節液泡pH值等方向進一步改良[21]。不久前完成的月季基因組測序獲得了當前植物基因組中質量最高的基因組信息,通過重建次生代謝及其調控通路,提出了月季花色和花香關聯的調控模型,為進一步闡明月季花色分子機制提供了基礎,能夠加速月季、薔薇科物種及其他觀賞植物的花色改良育種[42]。
菊花中F3'H基因是矢車菊素合成途徑的關鍵因,為了使花青素向飛燕草素合成途徑轉變,Huang等[43]采用RNAi技術抑制菊花CmF3'H的表達,且過表達瓜葉菊(Senecio cruentus)F3'5'H基因(PCFH),然而瓜葉菊F3'5'H酶在菊花中僅表現出3'-羥基化的功能,使轉基因菊花中的矢車菊素含量顯著增加。Brugliera 等[38]嘗試了利用 hairpin RNAi(hp RNAi)干擾菊花CmF3'H的表達,同時轉入三色堇(Violasp. cultivar black pansy)F3'5'H,使轉基因菊花中積累達80%的飛燕草素糖苷,花色由粉色變為紫色。
3.2 增加輔助色素含量
Aida等[44]研究一系列夏堇的轉基因植株發現,轉入反義DFR基因的花色比轉入反義CHS基因的花色更藍。原因是DFR基因功能受抑制導致輔助色素黃酮的含量顯著增加,因而使花色更藍。這一方法可應用于其它植物藍色花的育種。
3.3 提高液泡pH值
盡管已育成的藍色月季中積累了足夠多的飛燕草素,但是由于花瓣細胞液泡酸度過高,花色仍然呈現出一定程度的紅色色調[4]。因此要使花瓣呈現藍色,轉入基因創造一個pH值5.6-6.2微酸性的液泡環境,或者選擇天然具有較高pH值的受體材料尤為重要[32]。例如,由細胞培養得到的突變體培育而成的紫色仙客來主要含有錦葵素3,5-二葡萄糖苷,在此基礎上,通過隱性突變提高花瓣細胞pH值有望獲得藍色仙客來[32]。
3.4 增加金屬離子含量
在紫色郁金香花被片細胞中瞬時過表達液泡鐵離子轉運蛋白基因TgVit1,使鐵蛋白合成基因TgFER受抑制,阻止鐵蛋白積累并結合鐵離子,使細胞變藍[29,45]。進而 Shoji等[46]又利用花瓣特異表達的TgMYB1啟動子驅動TgVit1基因的表達,期望獲得藍色郁金香。此外,Kurihara等[47]將TgVit1在積累飛燕草素3,5-二葡萄糖苷的突變體仙客來中瞬時表達,獲得了藍色細胞,有望通過穩定表達得到藍色仙客來。
3.5 藍色花卉基因工程育種策略
綜合上述藍色花育種實踐,如要成功地利用基因工程創造有商業價值的花卉需要具備以下條件:(1)分離到有效的基因;(2)建立高效的目標植物遺傳轉化體系;(3)通過啟動子序列、目標基因的來源和終止子序列等方面來優化目標基因的表達;(4)選擇遺傳背景合適的受體品種[21]。根據藍色花形成的因素,創造藍色花應從以下方面入手:積累飛燕草素糖苷,其糖基被芳香有機酸酰化,或者加強花青素苷與輔助色素、和/或金屬離子之間的相互作用。
4 小結
盡管眾多育種實踐證實了通過基因工程的手段創造藍色花的可行性,然而讓目標植物特異地大量積累某一類色素并呈現藍色非常困難。首先需要考慮藍色花青素苷通常有復雜的糖苷化和酰基化修飾,這些修飾酶在異源植物中的活性是否能夠正常發揮,還需要考慮調節液泡pH值以及有輔助色素和金屬離子共存,這就需要對目標植物的花色形成機制有全面的認識。近年來日趨普及的全基因組測序工作將為全面解析植物花色形成的機制奠定堅實基礎。此外,對于非模式植物來說,很多觀賞植物普遍存在遺傳轉化效率低、童期長、轉入基因表達變異大等問題,攻克這些難題才有希望獲得更多新奇的商品化的藍色花卉。
