長江安慶段仔稚魚群落特征調查研究
2019-11-26 02:17:24李新豐丁隆強何曉輝方弟安徐東坡
水生生物學報 2019年6期
關鍵詞:優勢
李新豐 丁隆強 何曉輝 劉 熠 任 鵬 方弟安, 徐東坡,
(1. 上海海洋大學水產科學國家級實驗教學示范中心, 上海 201306; 2. 農業農村部長江下游漁業資源環境科學觀測實驗站,中國水產科學研究院淡水漁業研究中心, 無錫 214081)
魚類早期資源調查具有樣本量大、對資源破壞小、采樣簡單、費用較低的優勢, 使其成為進行魚類種群豐度評估研究重要手段之一[1], 仔稚魚作為魚類早期生活史中重點研究對象, 對仔稚魚在自然水域環境下的豐度動態變化研究可以了解魚類早期棲息地分布及發育過程中的環境需求等基礎資料[2], 進而為漁業資源的合理開發以及魚類資源保護政策制定提供科學依據。長江流域幅員遼闊,魚類區系種類組成復雜, 長江干流分布擁有豐富的產卵場、育幼場[3—5], 國內學者自20世紀60年代開始大規模的進行長江魚類早期資源調查, 時至今日已經積累了大量的仔稚魚類的形態特征、密度變化、群聚結構等較為完備的基礎資料。近年來, 國內學者對長江流域的魚類早期資源研究主要集中在中上游江段, 如段辛斌等[6]就三峽蓄水產漂流性卵的魚類產卵場影響進行了分析研究; 黎明政等[7]對長江上游武穴段的魚類早期資源群落結構做了調查; 萬力等[8]探討了漢中的產漂流性早期魚類的時空分布特征; 劉明典等[9]闡述了長江中游宜昌江段魚類早期資源群聚特征及四大家魚的產卵場的現狀; 對長江口魚類早期的資源量、群落特征和產卵場分布亦有不少研究報道[10—12]。然而關于長江下游段的魚類早期資源的研究報道寥寥可數, 僅見劉熠等[13]在2016年對長江湖口段仔稚魚群落結構特征的報道, 因此, 基于上述背景本文選取安慶江段作為魚類早期資源調查水域, 以期為長江下游早期資源研究提供基礎資料。
長江下游安慶段, 位于安徽省境內, 全長約243 km, 多沙洲, 屬于分汊型河段, 漁業資源豐富,該江段擁有長江江豚自然保護區、大口鯰保護區、長江刀鱭國家級水產種質資源保護區等重要生態區域, 是重要的漁業資源水域[14], 歷史資料顯示20世紀90年代該江段含有魚類種數63種[15], 張敏瑩等[16]在2006年報道46種, 張曉可[17]在2016年調查發現僅有35種, 由于大量涉水工程實施、過度捕撈和水體污染等強烈的人為干擾因素[18], 安慶江段的漁業資源日趨衰退。本文通過魚類早期資源調查的手段來分析安慶江段的仔稚魚的種類組成、豐度時間變化以及與相關環境因子相關性研究, 以期為該江段漁業資源評估及保護政策的制定提供依據。
1 材料與方法
1.1 采樣時間和采樣地點
本次研究, 首先通過查閱長江中上游魚類早期資源調查的歷史文獻[19,20]及現場勘查, 然后于2016年4月17日至8月15日在長江下游安慶段選取一個斷面(N30°28′59.07″; E116°59′39.36″)進行魚苗采集, 該斷面江岸平直, 斷面約寬1380 m。每天上午8: 00—11: 00進行1次采樣, 單次拖拽時間根據魚苗高峰期實時調整。該斷面設置3個采樣點分別在斷面的南岸、中間和北岸, 南北2個采樣點距離岸邊100 m左右, 在每個采樣點的表層(1 m以內)進行魚苗的采集。采樣的網具為直徑80 cm, 網深2.5 m,網目40目, 孔徑0.25 mm的圓錐網, 網后連接圓柱形集苗器(長20 cm, 直徑10 cm)。過網口的水量采用德國JKY/Hydrobios/438115數字網口流量計測量(m3), 采用溫度計測量表層水溫(℃), 透明度通過塞氏透明度盤測量(cm), 采樣期間斷面的水位和水流量數據來自長江水文網(http://www.cjh.com.cn/)的安慶水文站和大通鎮水文站(由于大通水文站位于安徽省池州市距離調查斷面下游80 km且無大型支流匯入長江干流, 故可以反映該斷面的水流量情況,圖 1)。
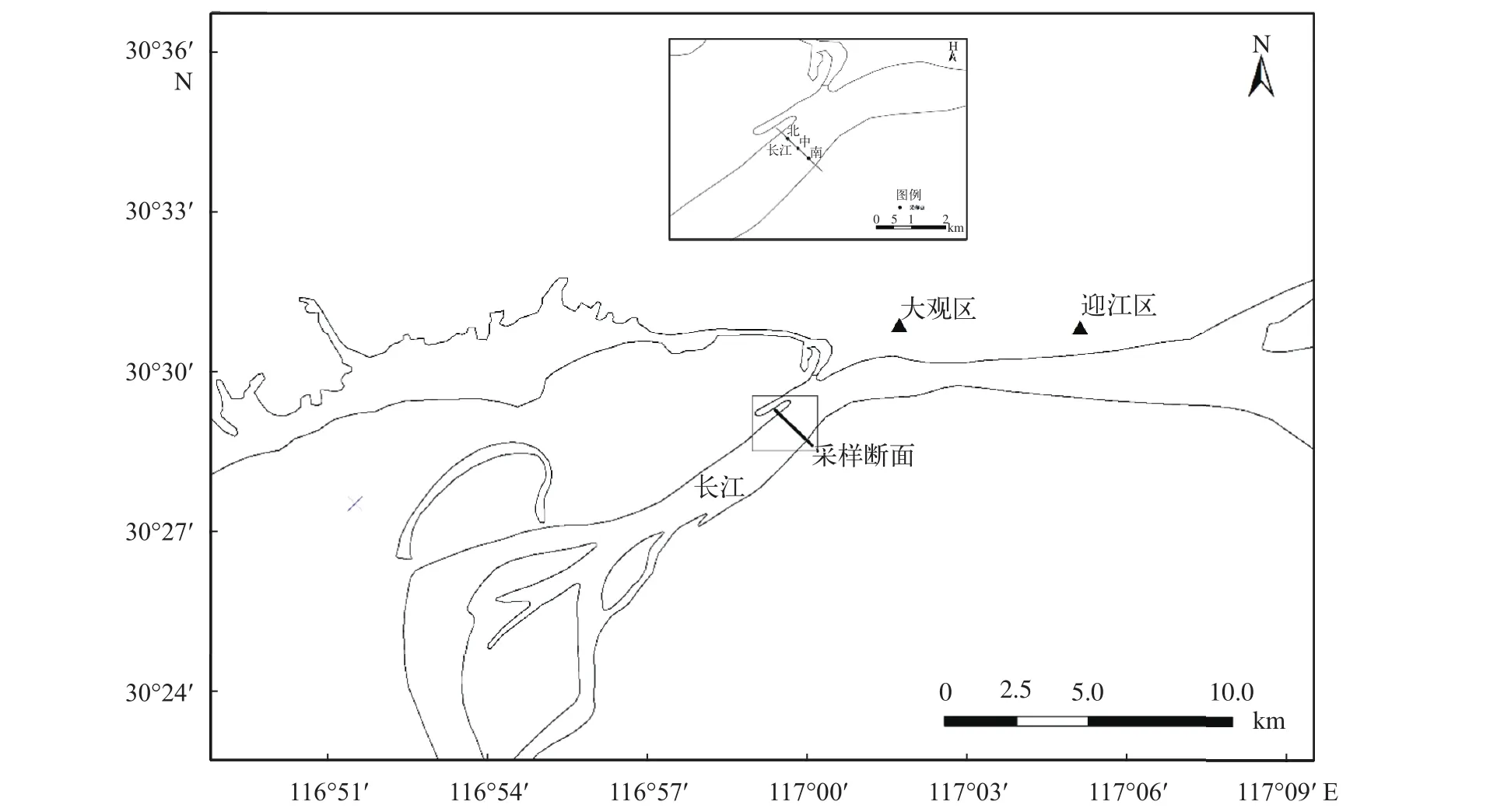
圖1 安慶江段魚類早期資源采樣點設置Fig. 1 The sampling sites on fisheries resources of early life history stages in Anqing section
1.2 樣品處理
在現場采集的魚苗冰水保存, 然后立即帶回實驗室, 參考曹文宣等[1]編著的《魚類早期資源》描述的各種魚類形態學特征, 通過形態手段方法在奧林巴斯解剖鏡(型號: SZX16)進行種類鑒定, 并分類計數, 對形態破損或無法分辨的仔稚魚進行編號保存于75%中性酒精中, 并通過分子生物學報進行種類鑒定, 由于該時期的體長較小, 因此對整個個體提取DNA, 然后經過PCR擴增COI基因序列, 最后在NCBI網站中與已經用成魚建立的長江魚類條形碼數據庫進行比對, 取遺傳相似度≥99%為對應種類[21], 若存在不同物種, 則舍棄該樣本。對已經鑒定完成的樣品, 適量選取形態較好的魚苗轉入5%的甲醛溶液, 侵泡1—2h, 最后通過奧林巴斯解剖鏡SZX16拍照軟件進行形態上的拍照保存。
1.3 數據分析
仔稚魚的豐度計算參考曹文宣等[1]和劉熠等[13]的計算方法, 以每100 m3水體含有仔稚魚數量(ind.),其計算公式為
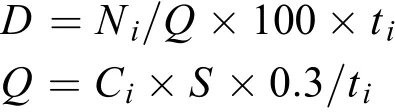
式中,D為采集期間的過網魚苗的平均豐度(ind./100m3);Ni為第i次采集仔稚魚的數量(ind.);Q為采集期間的過網水量(m3);S為圓錐網的網口面積(m2);ti為第i次采集時間(s);Ci為第i次采集時流量計的始末差值; 0.3為計算過網水量的公式系數。
生態優勢度: 選用Pinkas的相對重要性指數[22]描述出群落間的優勢種和重要種, 定義IRI>100為優勢種, 10<IRI<100為常見種,IRI<10為少見種[13]。計算公式為:
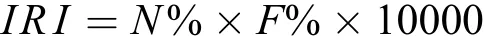
N%為某個物種占群落總數量的數量百分比,F%為某個物種被采集到的總天數占調查總天數的百分比。
1.4 冗余分析(Redundancy analysis, RDA)
將水流量、水位、水位、透明度4個理化因子與17種(IRI≥10)仔稚魚密度構成環境因子與物種矩陣, 首先對物種數據進行除趨勢對應分析(Detrended correspondence analysis, DCA)所得排序軸梯度長度(Length of gradient, LGA)來判斷哪種模型(線性或單峰)更為合適(LGA>4, 單峰模型; 3<LGA<4, 兩者皆可; LGA<3.線性模型), 本研究分析顯示各個排序軸均小于3, 表明仔稚魚密度變化與環境因子有較好的線性關系(適合RDA分析)。由于各物種和環境因子之間的數據存在很大的差異性, 離散程度高, 為了降低極端數據帶來的負面影響, 提高數據的正態性和方差齊性, 對物種數和環境因子數據均進行log(x+1)轉化[23], 并且通過蒙特卡洛檢驗(Monte-Carlo permutation test,P<0.05)來篩選環境因子[24]。
通過Excel 2016、SPSS22.0、Canoco 5.0和Origin9.1進行實驗數據的分析和圖形的繪制。
2 結果
2.1 仔稚魚數量及種類組成
物種組成本研究采集安慶江段仔稚魚共計104d, 捕獲92160尾, 實際鑒定90457尾, 共計49種,隸屬于7目11科(表 1)。鯉科魚類32種, 數量占比最大, 為90.16%; 其次是鰕虎魚科, 3種, 數量占比為5.71%; 其余9科, 總數占比不足5% (表 2)。
生態類型與產卵方式魚苗根據生態類型可分為3類: (1)江湖半洄游性魚類, 如鳊(Parabramis pekinensis)、鳡(Elopichthys bambusa)等8種,9533尾, 占比10.54%; (2)淡水定居性魚類, 如貝氏?(Hemiculter bleekeri)、?(Hemiculter leuciscu
lus)、銀鲴(Xenocypris argentea)等40種, 79217尾,占比87.57%; (3)溯河洄游性魚類, 如刀鱭(Coilia nasus) 1種, 1707尾, 占比1.89%。
從產卵方式可以分為3類: (1)產漂流性卵, 如貝氏?、鳊等20種79869尾, 占比88.30%; (2)產沉性卵, 如子陵吻鰕虎(Rhinogobius giurinus)、黃顙魚(Pelteobaggrus fulvidraco)、黃尾鲴(Xenocypris davidi)等25種8769尾, 占比9.69%; (3)產浮性卵, 如班鱖(Siniperca scherzeri)、鱖(Siniperca chuatsi)、刀鱭等4種, 1819尾, 占比2.01% (表 3)。
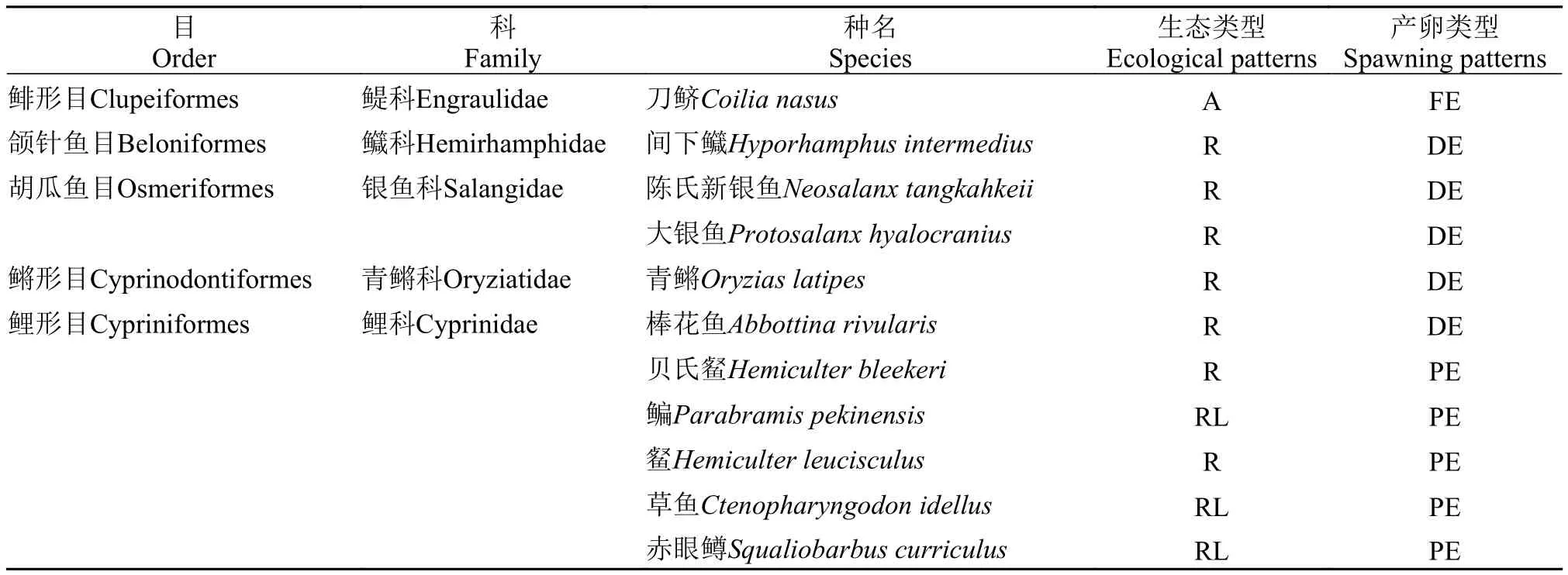
表1 2016年安慶江段4月17日—8月15日仔稚魚種類組成Tab. 1 Species composition of larval and juvenile fish in Anqing section from April 15th to August 17th, 2016

續表 1
2.2 仔稚魚豐度變化與優勢種組成
仔稚魚豐度變化特征2016年4月—8月期間, 仔稚魚豐度變化范圍37.48—1608.44 ind./100 m3,均值為407.56±353.25(Mean±SD), 峰值出現在6月15日。在調查期間內仔稚魚豐度出現4次明顯的高峰時期, 第一次在5月1日、后面三次集中出現在6月中下旬, 4次的高峰時期的仔稚魚豐度分別為1127.00、1608.44、1568.35 和1202.94 ind./100 m3。在6月24日之前仔稚魚的豐度變化是整體上呈現出上升趨勢, 6月26日—7月4日一直維持著相對較高的水平且并無明顯的漲幅, 但在7月4日之后仔稚魚豐度開始驟然下降, 7月5日—8月15日的仔稚魚平均豐度僅為139.08 ind./100 m3(圖 2)。
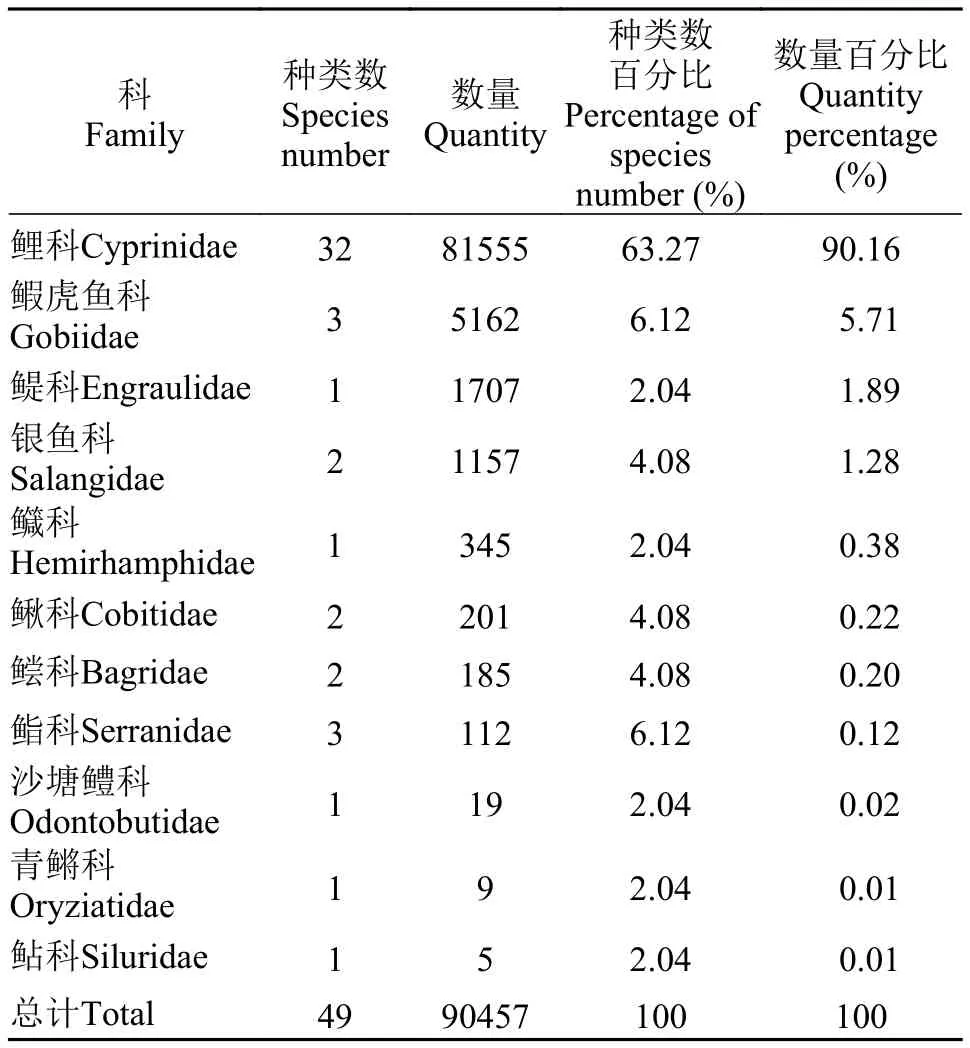
表2 各科所含的種類數和數量及其百分比Tab. 2 The species number and quantity contained in each family and their percentages
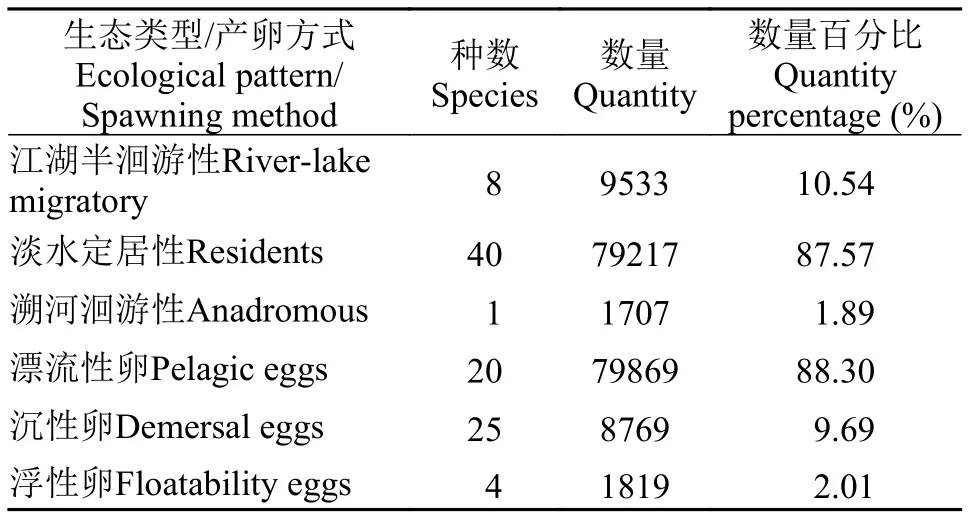
表3 仔稚魚生態類型/產卵方式的種類數及數量百分比Tab. 3 Species and quantity percentage of ecological type/spawning type of larvae and juveniles
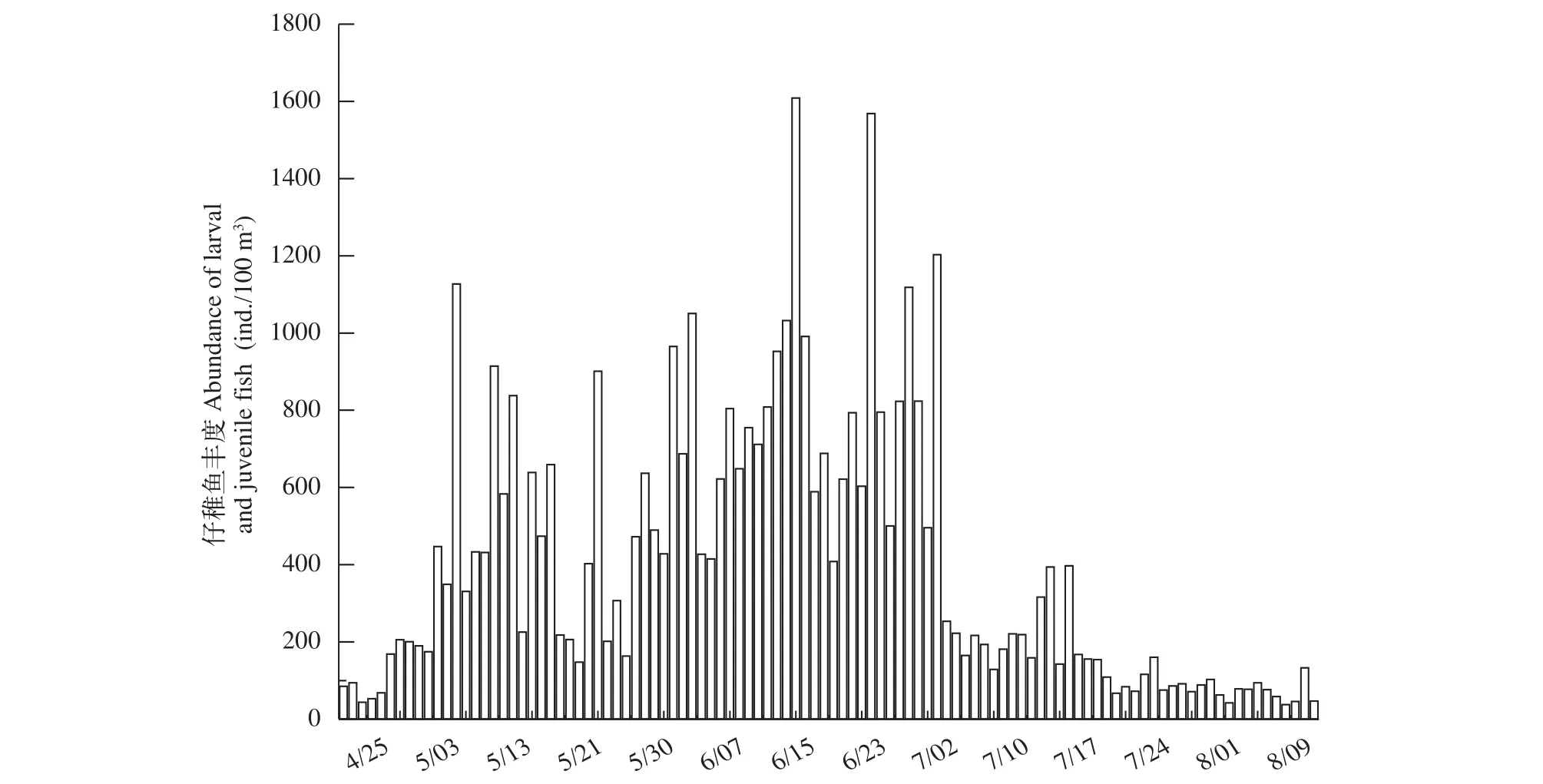
圖2 2016年4—8月安慶江段仔稚魚豐度變化Fig. 2 Abundance of larval and juvenile fish in Anqing section from April to August, 2016
仔稚魚優勢種組成通過公式IRI=N%×F%×10000計算得出, 相對重要性指數不低于100的共有10種仔稚魚, 按照其數值大小排列依次為貝氏?、?、銀鲴、子陵吻鰕虎、飄魚(Pseudolaubuca sinensis)、似鳊(Pseudobrama simoni)、寡鱗飄魚(Pseudolaubuca engraulis)、鳊、刀鱭、鰱(Hypophthalmichthys molitrix), 該10種優勢種數量占整個漁獲物的91.92%。其中貝氏?為第一優勢種, 數量占比約為44.53%, 在整個采樣期間, 每天均有出現; 第二優勢種為?, 數量占比約為14.21%; 四大家魚僅鰱魚成為優勢種且數量占比較小約為1.57% (表 4)。
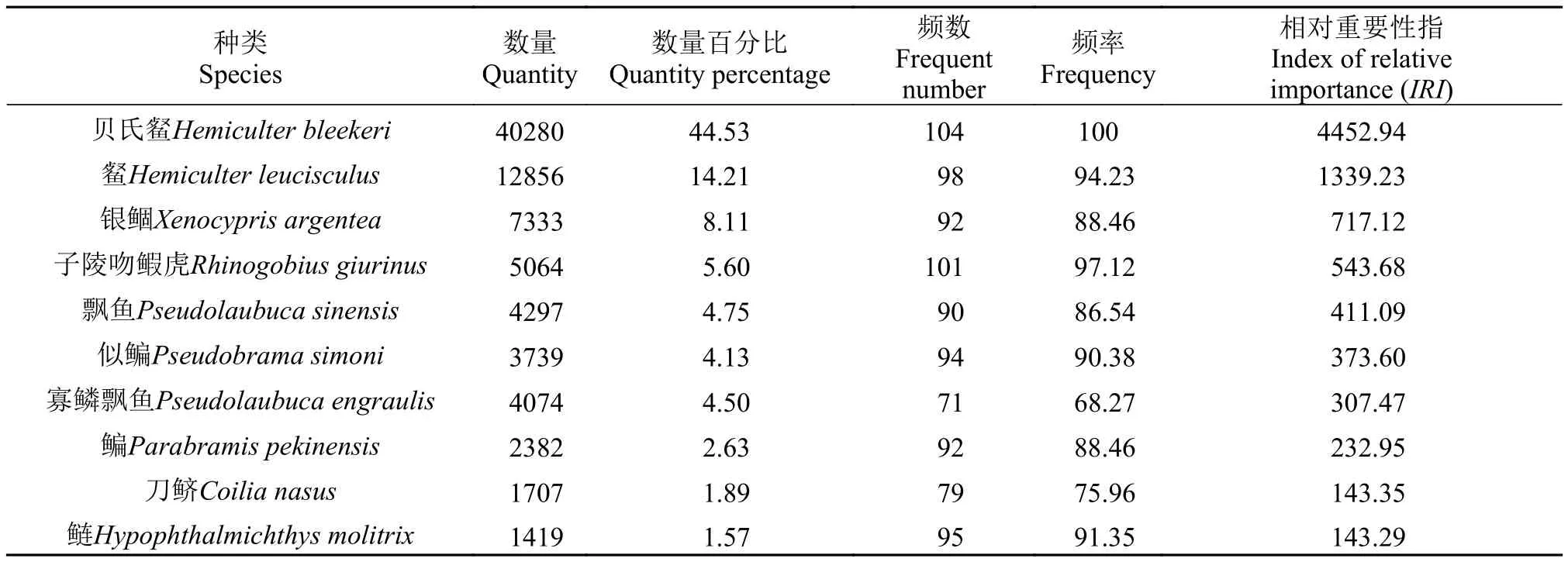
表4 安慶江段4—8月仔稚魚優勢種組成Tab. 4 Dominant species composition of larval and juveniles in Anqing section from April to August
優勢種豐度變化分析10種優勢種魚類有著不同的繁殖高峰期, 貝氏?為第一優勢種, 其豐度在整個監測期間都處于較高水平, 6月下旬之前呈上升趨勢, 之后開始急劇下降, 最大值出現在6月下旬, 為428.79 ind./100 m3; 銀鲴集中出現在5月上旬至6月下旬, 5月上旬豐度到最大值, 為74.97 ind./100 m3,其余時期豐度處于較低水平; 鰱豐度整體不高, 在上升至7月上旬時達到最大值為16.47 ind./100 m3; 刀鱭集中出現在6月下旬至7月下旬, 7月上旬豐度達到最大值, 為21.06 ind./100 m3; 寡鱗飄魚集中出現在6月下旬至7月下旬, 7月上旬豐度達到最大值, 為58.58 ind./100 m3; 其余鳊、?、飄魚、似鳊、子陵吻鰕虎的最大豐度均出現在6月上旬, 分別為31.84、124.46、43.82、43.45 和47.48 ind./100 m3(圖 3)。
2.3 仔稚魚豐度變化與環境因子的相關性分析
環境因子特征魚類的生長發育與各項水質指標息息相關, 在4月17日—8月15日期間所記錄的4個環境指標中水流量變化為34700—70700 m3/s,有2次明顯的上升時期分別在4月17日—4月25日和7月1日—7月15日, 在7月13日達到峰值, 之后開始快速下降; 水位變化為11.77—17.69 m, 其變化趨勢與水流量較為相似; 透明度變化為16—38 cm, 其變化趨勢在4月25日—5月10日和6月17日—7月14日波動性較大, 最小值出現在7月9日; 溫度變化為18—30℃, 一直處于緩慢上升趨勢(圖 4)。
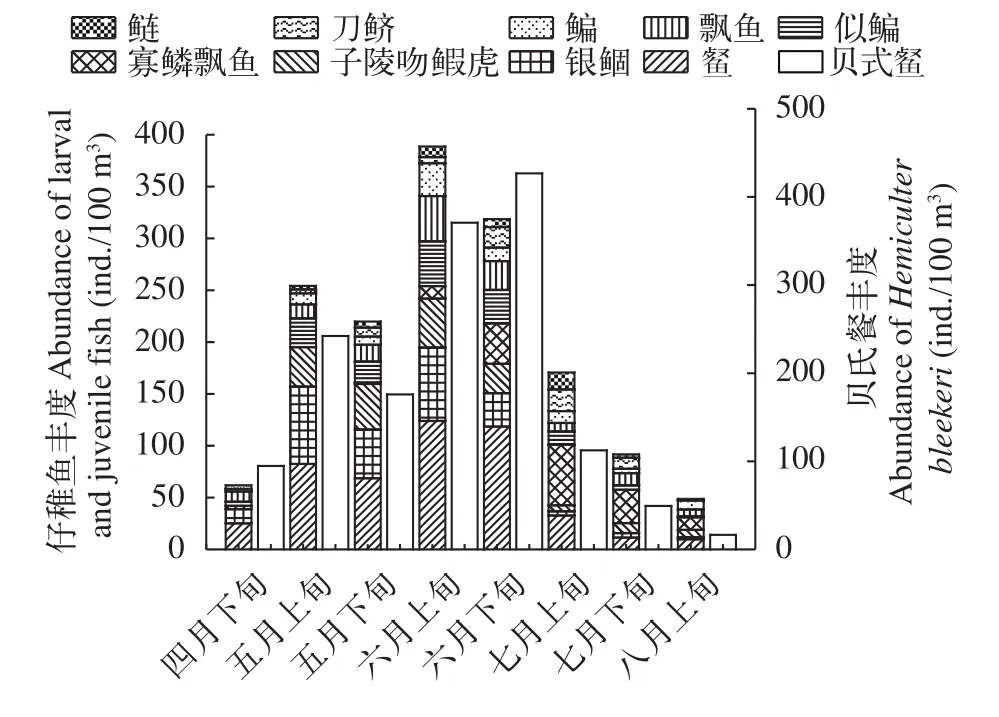
圖3 各月份優勢種魚類豐度Fig. 3 The abundance of dominant fish species in each month
仔稚魚密度變化與環境因子冗余分析(RDA)RDA結果顯示, 軸一、軸二、軸三的特征值分別為0.1673、0.0910和0.0136, 仔稚魚豐度特征在軸一、軸二、軸三解釋率分別為16.73%、9.09%和1.36%, 對仔稚魚豐度與環境因子關系累計解釋率達100%, 由此可知繪制的RDA分析圖能夠很好反映仔稚魚豐度變化和環境因子的關系。水流量對仔稚魚群落豐度變化貢獻率最大, 依次為透明度、水位, 并且解釋率分別為14.1%、6.0%和7.0%。透明度為主要的正相關因子(P<0.05), 如與貝氏?、?、陳氏新銀魚(Neosalanx tangkahkeii)、鳡、間下鱵(Hyporhamphus intermedius)、似鳊、飄魚、銀鲴、子陵吻鰕虎均呈正相關, 僅與寡鱗飄魚呈負相關。水流量和水位為主要的負相關因子(P<0.05), 如水流量和水位均與貝氏?、?、草魚(Ctenopharyngodon idellus)、陳氏新銀魚、鳡、黃尾鲴、銀鲴、似鳊、飄魚、子陵吻鰕虎呈負相關,均與刀鱭、寡鱗飄魚和翹嘴鲌(Culter alburnus)呈正相關 (圖 5)。
3 討論
3.1 安慶江段仔稚魚資源現狀
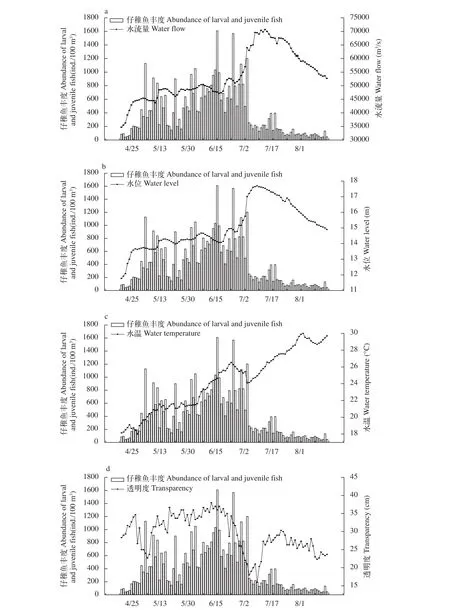
圖4 2016安慶江段仔稚魚豐度時間變化與環境因子的關系Fig. 4 The relationship between the abundance of larvae and juvenile fish and environmental factors in Anqing section in 2016
魚類的仔稚階段是整個生命周期內最重要時期, 是生命活動的開端, 仔稚魚群落的波動直接關系到成魚的群落結構[25], 基于安慶江段成魚資源的日益衰退, 從早期魚類調查的角度來了解該水域的魚類群落現狀。本次調查顯示, 仔稚魚種類數為49種, 與中上游江段的仔稚魚種類數相比[7,19], 安慶江段種類較為豐富。同2012—2013年任鵬[26]在安慶江段調查顯示的52種類群仔稚魚相比, 略微下降。在優勢種數方面, 為10種, 與中上游江段[7,19]及安慶江段在2012—2013年[26]優勢種數相比擁有一定優勢。安慶江段仔稚魚的較高水平豐度集中在5月和6月, 其中6月的仔稚魚豐度最高, 與其余各江段的研究結果相似[7,27]。貝氏?一直處于絕對優勢種的地位, 數量百分為44.57%, ?作為第二優勢種,這與長江中上游具有類似情況[7,28]。優勢種中的重要經濟性魚類在本次調查發現有3種, 為鳊、刀鱭及“四大家魚”僅有的鰱, 并且優勢種中多以小型魚類為主, 如貝氏?、?、似鳊、寡鱗飄魚、飄魚等。總之, 雖然安慶江段的仔稚魚階段魚類種類在長江流域內較為豐富, 但同樣面臨著資源衰退可能, 且魚類小型化和經濟魚類資源量較低的現象依然存在。
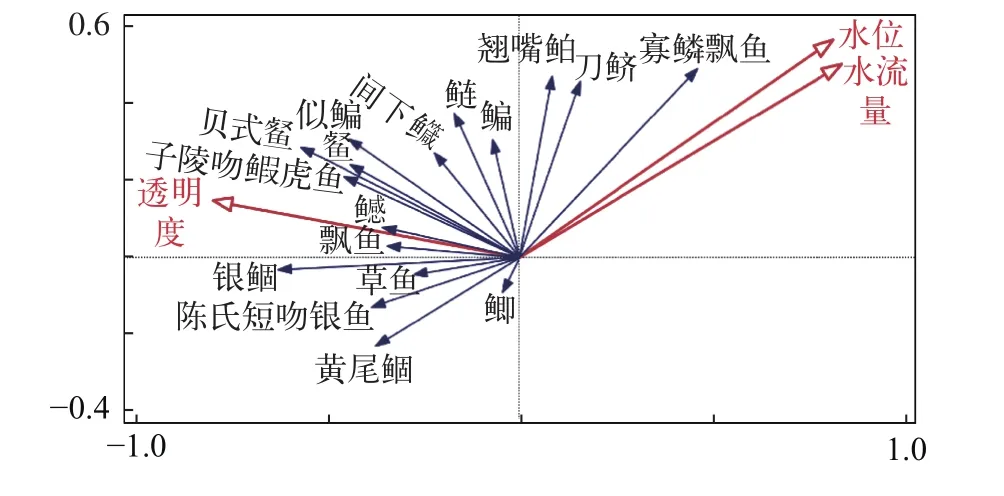
圖5 安慶江段仔稚魚物種-環境關系的RDA二維排序圖Fig. 5 RDA biplot of larvae and juvenile fish species and environmental variables in Anqing section
3.2 安慶江段仔稚魚生態學特征
安慶江段屬于相對穩定的分汊型河流, 擁有眾多的洲灘, 如白沙洲, 鵝毛洲, 江心洲等[29], 洲灘經過多年江水沖刷, 堆積了豐富的營養物質, 為淡水定居性魚類孕育提供充足的餌料, 加之自身的適應性較強, 僅需在局部水域內就可完成整個繁殖活動,因此淡水定居性的仔稚魚種類數為40種, 占比83.33%, 尾數占比87.57%。同時, 安慶江段擁有眾多的附屬湖泊[30], 如白蕩湖、石門湖、嬉子湖、菜子湖等形成典型的江湖復合生態系統[31], 該種生態系統為江湖半洄游性魚類的繁殖和索餌過程提供了優越的生境條件[32]。然而本次調查發現江湖半洄游性魚類種類數僅為8種, 尾數占比10.54%, 遠小于湖泊定居性魚類, 可能由于安慶市是長江下游重要的化工城市, 安慶江段及沿江湖泊環境污染日益嚴重[30], 同時漁業過度捕撈一直存在, 使得江湖半洄游性魚類的繁殖群體規模嚴重縮小。刀鱭作為唯一一個安慶江段仔稚魚群落中河海洄游性魚類,結果與任鵬[26]研究相同, 可能是由于安慶江段至河口距離較遠, 受潮汐作用非常小, 多數江海洄游性魚類已無法洄游至該江段, 刀鱭的數量占比也僅在1.89%。
卵期的下一個階段即為仔稚魚期, 魚卵的生態習性, 也一定程度上決定著仔稚魚群落結構。本次研究發現產漂流性卵魚類數量占比最大, 約占88.30%, 其次為產沉性卵的魚類, 但在種類數方面該兩種產卵類型的魚類較為接近。多位學者發現水流量對產漂流性魚類有很大的促進作用[33], 安慶江段在采集期間出現過水流量上漲較快的時期, 使得產漂流性卵魚類在數量上占據了絕對的優勢。產沉性卵的魚類雖在數量占比較小, 但種類含量較為豐富, 水流量的上漲必定引起水位的增加, 覆蓋了河道兩岸的植被, 為具有黏性的沉性卵提供附著物, 也為所有沉性魚卵提供了良好的避害空間和繁育場[31]。
3.3 環境因子對仔稚魚豐度的影響
仔稚魚的豐度變化受到內源因素和外源因素共同的作用[19]。在內源因素方面, 魚類繁殖周期長、發育時間短、攝食效率高可以大大提高成活率, 進而影響仔稚魚豐度的變化規律[8,19], 如優勢種中的貝氏?、?、似鳊在豐度大小上占有一定的優勢, 繁殖時期均在4—8月, 時間跨度大, 同時快速的世代更替率及攝食性廣的內源性因素使其能夠不斷補充資源量[8]。銀鲴的高豐度時期集中在6月下旬之前, 寡鱗飄魚的高豐度時期集中在6月下旬之后, 說明不同種魚類之間具有不同的繁殖策略,可有效的減少魚類之間攝食資源的競爭[34], 也是一種提高自身存活率的內源機制。
在外源因素方面, 本文通過對安慶江段仔稚魚豐度與環境因子之間的冗余分析, 顯示水流量、水位、和透明度是最為主要的影響因子。水流量表示江水在單位時間里通過一個斷面的水體體積, 是長江流域的重要水文指標, 一般認為水流量增大會引起水位的上漲, 當春夏季雨量增加, 匯入江水, 致使江水流量加大, 水位提升, 形成季節性洪水, 在洪水脈沖作用下使得江河泛濫平原系統進行營養物質的循環和能量的流動, 進而促進江水中的魚類種群繁衍生息[35]。本次雖結果表明水流量和水位對仔稚魚豐度變化有著較高的貢獻率和解釋率, 但對大多數種類是呈現負相關, 這與唐錫良[19]和郭國忠[27]得出結論存在差異。可能此次調查期間的水流量和水位的變化幅度、均值、最大值均顯著高于其他學者監測的水文數據, 水流量和水位的增加的雖可以促進仔稚魚的數量同步增長, 但當水流量和水位的漲幅速度相比仔稚魚過快時, 此外過高的水位使得兩岸淹沒, 河道拓寬, 仔稚魚獲得了更大的活動空間, 并且向新的岸邊聚集, 而采樣點并未變化,樣品的獲得量可能具有誤差, 因此呈現出不同的結果。透明度主要受水體中泥沙的含量影響, 一般認為當水流量和水位上升時, 透明度會下降, 本文RDA結果顯示透明度對大多數仔稚魚豐度的影響呈現正相關, 該種關系更可能為間接的作用。也有學者指出當透明度降低, 泥沙含量高, 能見度下降,會通過增加仔稚魚避害能力, 從而增加其豐度[36]。多數學者認為水溫也是影響仔稚魚豐度的重要影響因子之一[37,38], 然而本次研究并未得出此結論,可能由于調查周期跨度大, 7月之后水溫仍繼續上升與仔稚魚在此之后的急劇下降屬于自然的客觀規律, 因此致使在整體上的RDA分析并無相關性,但在7月之前水溫的上升, 仔稚魚豐度出現明顯的增加。
猜你喜歡
我愛學·笑話與口才(2025年3期)2025-02-24 00:00:00
華人時刊(2020年13期)2020-09-25 08:21:30
趣味(語文)(2020年3期)2020-07-27 01:42:46
意林·全彩Color(2019年8期)2019-09-23 02:12:36
作文與考試·初中版(2017年12期)2017-04-19 20:26:27
中國衛生(2015年2期)2015-11-12 13:13:54
中國火炬(2014年11期)2014-07-25 10:31:58
中國體育(2004年3期)2004-11-11 08:53:02
棋藝(2001年19期)2001-11-25 19:55:34
棋藝(2001年23期)2001-01-06 19:08:36
