中國東南部不同海拔亞熱帶森林中馬尾松徑向生長對氣候的響應
2019-12-06 09:00:30黎敬業黃建國梁寒雪蔣少偉周鵬郭霞麗李軍堂
熱帶亞熱帶植物學報 2019年6期
關鍵詞:生長
黎敬業, 黃建國, 梁寒雪, 蔣少偉, 周鵬, 郭霞麗, 李軍堂
中國東南部不同海拔亞熱帶森林中馬尾松徑向生長對氣候的響應
黎敬業1,2,3, 黃建國1,3*, 梁寒雪1,3, 蔣少偉1,2,3, 周鵬1,2,3, 郭霞麗1,2,3, 李軍堂4
(1. 中國科學院華南植物園, 中國科學院退化生態系統植被恢復與管理重點實驗室, 廣東省應用植物學重點實驗室, 廣州 510650; 2. 中國科學院大學, 北京 100049; 3. 中國科學院核心植物園, 廣州 510650; 4. 興山縣國有龍門河林場, 湖北 宜昌 443700)
為了解我國東南部亞熱帶森林不同海拔樹木生長對氣候響應的差異,建立了福建省武夷山脈東麓2個樣點的4個馬尾松()輪寬年表,對樹木徑向生長與氣候因子進行了bootstrapped相關分析和線性混合模型(LME)擬合。結果表明,在高海拔地區馬尾松徑向生長對氣候因子年際波動敏感性較強,主要表現為與生長季前冬季光溫條件以及生長季內7月降水的正相關,生長-氣候關系在不同樣點間表現出較強的一致性。線性混合模型可以較好地擬合高海拔樹木生長變化, 當使用前1年12月平均日最高溫、當年1月日照時長和當年7月降雨量3個氣候變量進行擬合時,模型解釋量達到0.5,其中前1年12月最高溫和當年1月日照時數在模型中起到主導作用,累積相對貢獻率約占80%,說明生長季前冬季的光熱條件是限制高海拔馬尾松徑向生長最主要的氣候因子。因此,我國亞熱帶地區高海拔的樹木徑向生長可能對未來氣候變化有更強的敏感性,相關森林管理政策的制定需要將此納入考慮;同時我國亞熱帶地區高海拔森林中的樹木有被用于樹輪氣候重建的潛力。
馬尾松;徑向生長;亞熱帶森林;海拔;氣候;線性混合模型;中國
隨著全球氣候變化加劇,世界范圍內的樹木生長普遍受到影響[1-3]。而由于氣候變化在不同區域的表現形式存在差異,且不同生境中的樹木生長存在不同的氣候限制因子,故樹木生長對氣候變化的響應也存在著多變的空間格局,探明這種生長-氣候關系的空間異質性對于理解和預測森林生態系統在氣候變化背景下的響應動態有著重要意義。目前該領域的研究主要沿兩個梯度展開,即緯度梯度和海拔梯度。在緯度梯度方面,總體表現為高緯度地區樹木生長受到生長季前期增溫的顯著促進[4-5], 同時受到生長季中后期高溫干旱的抑制[6-9]。與之相比,生長-氣候關系隨海拔梯度變化的方式更為多樣[10],有研究表明,歐洲堪薩斯地區和南美安第斯山脈的樹木生長與溫度關系表現為高海拔的正相關和低海拔的負相關[11-12],而波蘭塔特拉山地區的挪威松()徑向生長與溫度的關系卻表現為低海拔的正相關和高海拔的負相關[13]。這種海拔梯度上不均一的生長-氣候關系有可能導致山地森林生態系統在氣候變化中的響應模式表現出很高的空間異質性,進而可能導致森林群落結構產生垂直方向上的改變,對森林固碳潛力和物種多樣性的影響意義深遠。因此,我們迫切需要對此開展深入的研究,為相關地區森林管理和保育政策的制定提供參考。
在國內,大部分關于樹木生長-氣候關系海拔差異的研究集中在中部、西部和北部地區,東南地區未見相關報道[14-17],而中國的東南部又恰是多山地區,海拔異質性很高,南嶺山脈、武夷山脈、江南丘陵和東南沿海丘陵都集中分布于此。大量山地森林生態系統孕育出了豐富的物種多樣性,也是人口聚集的東南沿海城市重要的水源涵養區和生態屏障,多個國家級自然保護區和更多的省級自然保護區都劃設在這一地區。因此,為了填補樹木生長-氣候關系海拔差異研究在我國東南部的空白,我們開展了樹輪生態學試驗。馬尾松()作為在中國亞熱帶地區分布最廣,且具有重要的生態和經濟價值的針葉樹種[18-19],對其氣候響應特性進行研究有重要的理論和應用價值,又由于其樹輪中早晚材間界限清晰,易于進行交叉定年工作,因此被選做該研究的目標樹種。武夷山脈是我國東南部省份重要的生態屏障和水源涵養地,區內海拔落差大,氣候受東亞季風和西南季風的強烈影響,適合進行樹輪氣候和樹輪生態學試驗,故而被選作研究區域。
本研究希望回答以下科學問題: (1) 研究地區內樹木生長與氣候要素的關系是怎樣的?這一關系是否存在海拔差異?(2) 地區內同海拔相隔較遠的樣點間樹木生長-氣候關系是否存在相似性?這樣的相似性是否存在海拔差異?(3) 用氣候要素對樹木生長進行模型擬合,擬合優度是否存在海拔差異?模型中體現出的不同海拔樹木生長的主導氣候因子是否隨海拔變化?針對上述科學問題,我們提出以下假設: (1) 由于高海拔地區溫度較低,樹木徑向生長可能主要受到冬春季溫度的影響,表現為正相關,低海拔這一相關性不顯著;(2) 相隔較遠的樣點間樹木徑向生長與氣候的關系應該類似,且相似性在高海拔地區更強;(3) 用氣候因子的年際波動對徑向生長的年際波動進行擬合對高海拔樹輪年表的擬合效果要優于低海拔年表。
1 樣地概況
采樣區域(圖1)位于中國福建省境內的武夷山脈東麓(25.3°~26.6° N,116.8°~117.4° E),區內為亞熱帶季風氣候,年最高月均溫出現在7月,約23℃,年最低月均溫出現在1月,約4℃。該區年均降水量約1 800 mm,約65%的降水量出現在1-6月(圖2)。本試驗設置的2個采樣點WY1和WY2在武夷山脈東麓,與武夷山主脈呈平行分布,東北部的WY1樣點與西南部的WY2樣點相距約160 km。兩樣點內在高、低海拔各設置1樣方(20 m×20 m),高海拔樣方位于1 200~1 300 m,低海拔樣方位于700~ 800 m,海拔高度差大約500 m。所有樣方均在東南至西南方向的陽坡,坡度在20°以內,樣方內土壤和植被類型(次生或原生常綠針闊混交林)均較為一致, 以最大化減少由于立地條件引入的系統誤差。

圖1 采樣區域和樣點分布圖。▲: 采樣點; ●: 氣象站。

圖2 采樣區域氣象要素1987-2014年的月均值。Tmax: 日最高溫(0.1℃); T: 日均溫(0.1℃); Tmin: 日最低溫(0.1℃); SunH: 總日照時數(h); 柱子: 總降水量(mm)。
2 材料和方法
2.1 樹輪樣本處理
根據每個樣方內馬尾松的數量,隨機選取10~20棵長勢正常的馬尾松()作為目標樹,使用5.5 mm直徑生長錐在胸徑高度(約1.2~ 1.3 m)沿與坡向垂直90°方向取2根樣芯。樣芯隨即被保存在兩端封口的6 mm直徑吸管中帶回實驗室。樣芯在實驗室被固定在木槽中,經過干燥、打磨等處理得到樹輪清晰可辨的樹輪樣本。樹輪樣本經過交叉定年后,使用Lintab樹輪測量平臺和配套TSAP軟件(Frank Rinntech, Heidelberg, Germany)對輪寬進行測量(精確到0.01 mm)。測量后的輪寬序列使用COFECHA軟件[20]對定年質量進行檢測和輔助校正。最后,每個樣方內的所有輪寬序列使用ARSTAN軟件[21]進行去趨勢(使用50%序列長度的樣條函數對輪寬序列進行標準化,從而盡可能去除樹木本身年齡和其它低頻信號造成的生長速度變化趨勢,提高處理后輪寬年表內年際間氣候波動信息的信噪比)并整合成1個標準年表(Standard chro- nology),作為后續數據分析中的輪寬指數(RWI)使用。
2.2 氣候數據
本研究采用的溫度、降水量和日照時數使用中國國家氣候信息中心數據共享平臺(http://data.cma. cn)下載的中國地面氣候資料月值數據集,由于采樣點距離附近氣候站點遠近不一,海拔差異大,且各氣候站點在研究年份區間內有不同程度的數據缺失,因此本研究采用克里金插值法, 對已有站點的氣候數據進行空間插值, 得到研究樣點連貫和更為準確的氣候數據。此外,由于水分條件不僅受到降水的影響,也與土壤性質和溫度密切相關,為將上述因素納入考慮,自適應帕爾默干旱指數(scPDSI, CRU self-calibrating PDSI 1901-2016 0.5° Global 3.25, http://climexp.knmi.nl)也被用于相關分析中(scPDSI數值沒有量綱,其數值與所表征的濕潤程度正相關)。
2.3 數據分析
首先,為了展現各樣方內馬尾松生長對氣候條件年際間波動的響應模式,并對比其海拔差異, 采用R軟件中的bootRes函數包計算bootstrapped相關分析[22](重采樣次數=1 000),包括各樣方年表與各月值氣候要素(月均日最高溫Tmax、月均日最低溫Tmin、月總降水量PCP、月總日照時數SunH和月值自適應帕爾默干旱指數scPDSI)的相關系數。本研究中,用于計算bootstrapped相關系數。考慮到生長季前的氣候條件有可能影響到生長季內的樹木生長[4,23-25],本研究使用的氣候數據覆蓋了自生長季前一年10月至生長季當年12月的月份區間。
其次,為了評估氣候條件對不同海拔馬尾松徑向生長速率年際波動的解釋能力,采取線性混合模型分別對高低海拔年表分組(每組2個樣方,高海拔為WY1-H和WY2-H,低海拔為WY1-L和WY2-L)進行擬合。相比普通多元回歸,線性混合模型可以處理變量內各觀測值之間不獨立,不服從正態分布,或者響應變量觀測值內的不同分組與對應的自變量線性關系不一致的數據。在本研究中,同一海拔分組內2個年表內的輪寬指數共同組成一個響應變量,其內部分屬于不同年表的觀測值與對應氣候數據的線性關系(斜率)會存在差異,此時如果使用普通的線性模型進行處理,將會導致模型解釋量偏低,低估氣候變量對樹木生長的解釋能力[26-27]。線性混合模型的結構采用公式W=0+β+(0+μ)X+ε計算。
本研究構建的模型中,固定效應為氣候因子,而隨機效應為樣方(年表)差異。W和X分別代表標準年表中樣方和年份的輪寬指數以及對應樣方和年份的氣候數據;0和β分別代表固定效應和隨機效應的截距;0和μ分別代表固定效應和隨機效應的斜率;ε代表無法被固定效應和隨機效應解釋的隨機誤差。本研究采用R軟件的函數包[26]進行線性混合模型的計算和分析。為了決定最終模型中應該包含哪些氣候要素,我們先進行了自變量只包含單一氣候要素的模型分析,然后選取解釋量最高的若干變量組合納入最終模型內。最后,為了比較不同氣候要素對研究地區內不同海拔內樹木生長影響的主次差異,我們對上最優模型中各變量的相對貢獻率進行了分解估計[28]。
3 結果和分析
3.1 年表基本統計特征
本研究建立了4個標準年表(高海拔WY1-H, WY2-H; 低海拔WY1-L, WY2-L)。從各年表的歷史折線圖看,兩高海拔年表間的同步性較好,存在較多重合的特征年,而兩低海拔年表的同步性較差, 特征年基本不重合(圖3)。對年表的公共區間(1987- 2014)分析,高海拔的年表相對低海拔表現出更高的平均敏感度(MS),說明高海拔樣點內的馬尾松徑向生長速率(樹輪寬度)年際波動中所含高頻信號較多。此外,高海拔樣方內各目標樹輪寬序列的主成分1 (PC1)所解釋的總體變異比例(平均58%)也顯著高于低海拔樣方(平均35%)(表1),說明高海拔樣方內不同馬尾松個體之間徑向生長速度年際波動的同步性要顯著高于低海拔樣方。根據上述現象可以推論,研究區域內高海拔森林內馬尾松徑向生長速率可能對氣候要素的年際間波動有更強的敏感性。對各年表1987-2014年的公共區間進行分析,EPS為0.84~0.89, 樣本量偏少可能是EPS值偏低的原因, 大致符合樹輪學年表建立質量要求(EPS>0.85)[29],是較為可信的年表數據,后續分析都將基于這一年份區間進行(表1)。
3.2 Bootstrapped相關分析
首先,對單年表與單氣候要素進行bootstrapped相關分析(圖4),顯著相關性大部分出現在高海拔年表和氣候要素的組合中,且在不同樣點間呈現出良好的一致性。這驗證了在研究區域內,高海拔馬尾松徑向生長對氣候因子的敏感度強于低海拔樣點, 且不同樣點間馬尾松徑向生長在生長季內具有較為統一的氣候性限制因子種類以及時空分布格局, 也側面體現出研究區域內影響馬尾松生長的生態因子在高海拔地區具有較低的空間異質性。
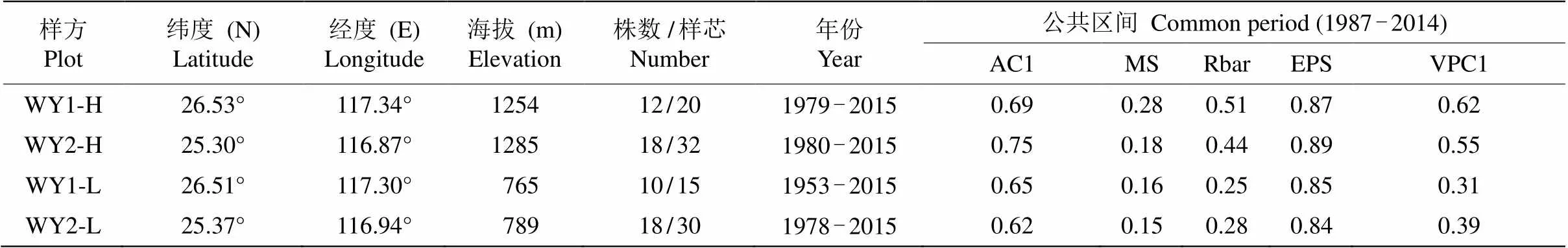
表1 樣方信息和各樣方STD年表統計量
AC1: 一階自相關; MS: 平均敏感度; Rbar: 序列間平均相關系數; VPC1: 全體序列主成分1解釋量。
AC1: 1st order autocorrelation; MS: Mean sensitivity; Rbar: All series correlation; EPS: Expressed population signal; VPC1: Variance in PC1 of all series.
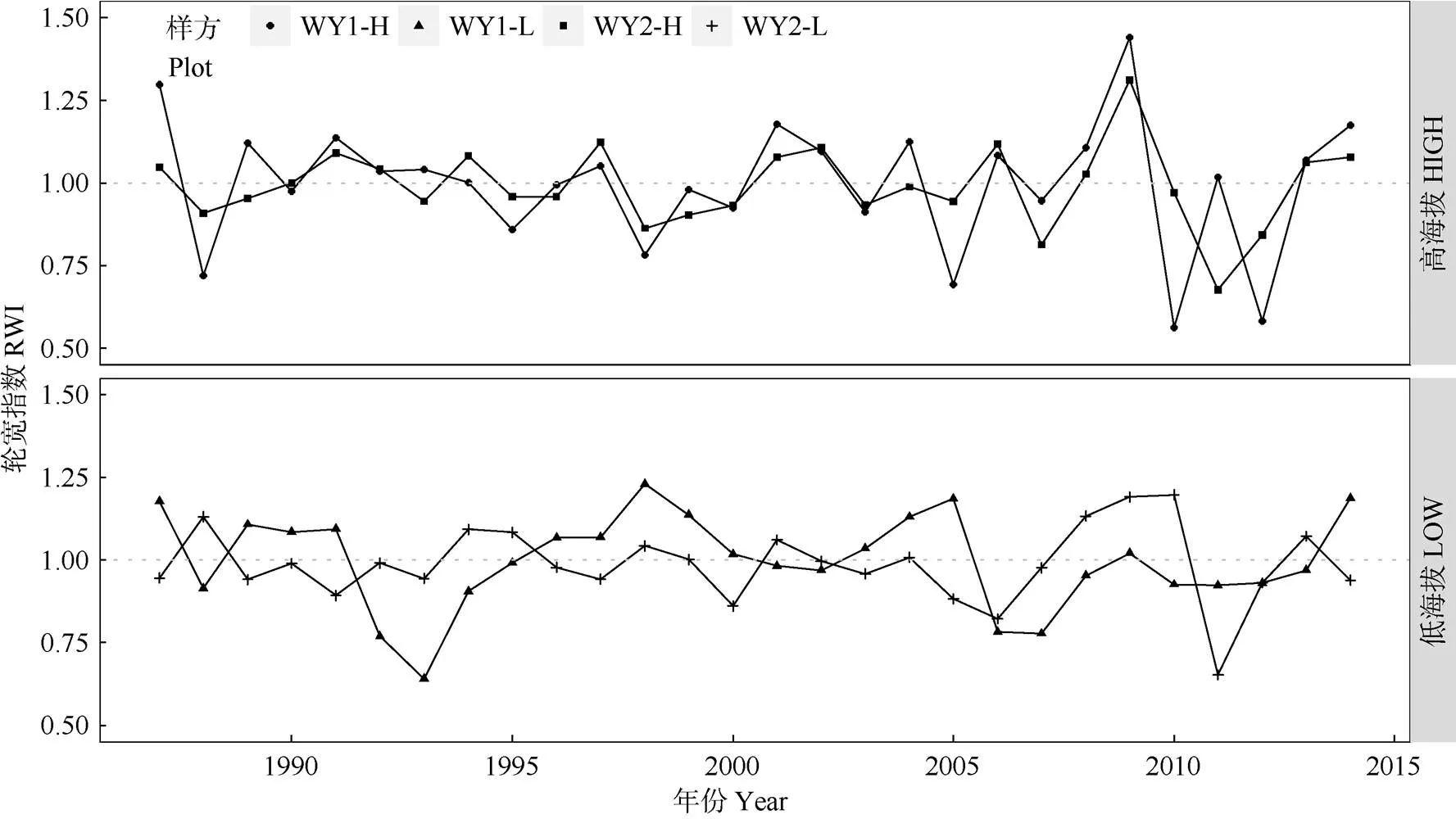
圖3 高低海拔樣點的輪寬指數
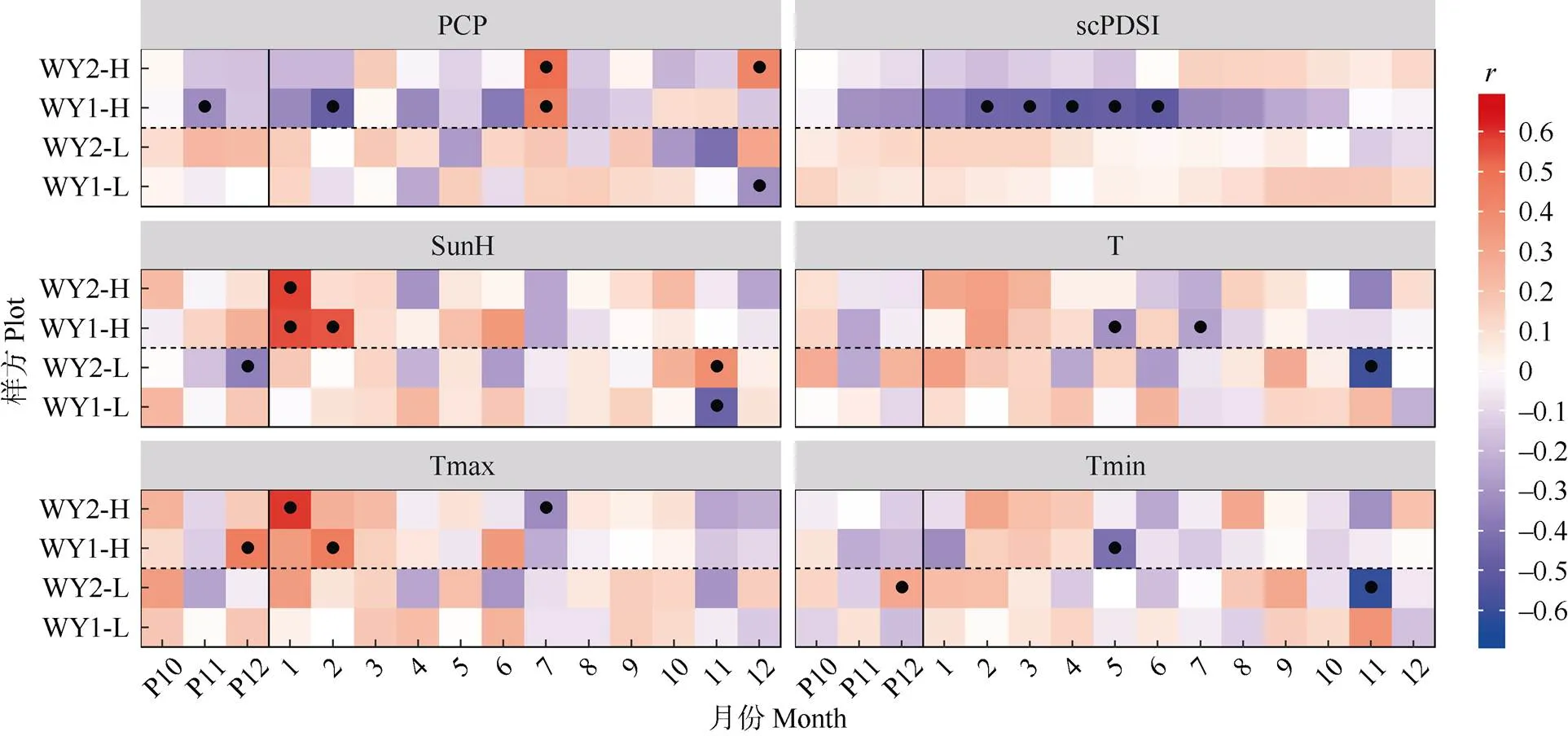
圖4 標準年表與氣象要素的bootstrapped相關系數。●: P<0.05。P10~P12: 前1年的10-12月。以下圖表同。
具體地說,高海拔馬尾松年表與生長季前1年12月至當年1-2月的溫度和日照時長呈現顯著的正相關(<0.05),說明生長季前冬季的光熱條件對馬尾松生長有著重要的影響。然而,此前有報道認為樹木形成層活動起始主要由日最低溫控制[30-33],而馬尾松形成層開始活動的臨界溫度約為11℃[34]。在本研究區域內,12-2月的平均日最低溫都遠低于10℃,考慮到高海拔地區相對于低海拔地區由于對流層大氣垂直溫度直減率導致的溫降,高海拔樣方內的冬季溫度更是遠遠無法滿足馬尾松形成層活動的要求。近年在本研究地區西南方向廣東省低海拔地區開展的微樹芯研究也證明,當地馬尾松形成層活動在冬季進入半休眠狀態,并不分裂產生木質部細胞[32]。綜上可以推斷,研究地區內的馬尾松徑向生長在冬季(12-2月)是停止的。因此,本研究中高海拔馬尾松年表與冬季溫度/光照呈現出的顯著正相關更可能是由于較高的冬季溫度和光照時長對休眠期馬尾松葉片光合作用產生了促進作用。有研究表明,生長季前期樹木光合作用的起始溫度閾值要遠低于木質部活動的起始溫度閾值[35-37],樟子松()的形成層活動溫度閾值大約為5℃~ 10℃, 而其光合作用在0℃以上就可以進行[38-39]。據此推測本研究地區內冬季溫度很可能可以支持馬尾松通過光合作用固定能量和碳,供給此后的生長季內消耗,那么在此期間的溫度和日照水平的提升顯然對樹木生長是有利的。
此外,高海拔年表總體上在6月以前表現出對降水量和帕爾默干旱指數呈負相關,而在7月突然轉變為顯著的正相關。這說明,上半年內高海拔區域馬尾松生長得到的水分供應是充足的。由圖2可見,研究地區內年降水量的近65%都分布在1-6月。在此階段,過多降水會減少光照,降低樹木的凈光合速率,且有可能導致土壤水分含量過高,造成根系缺氧,抑制樹木生長。而由圖2可見,研究地區內降水量從7月開始驟減超過50%,與此同時,氣溫在7月達到年中的最高值,馬尾松生長較易面臨水分虧缺和高溫脅迫,從而表現出對7月降水量的正相關關系。
最后值得注意的是,低海拔年表除在生長季前1年12月以及當年11月之外,其與氣候要素的相關性均不顯著,且即使是在上述月份,不同樣點間的生長-氣候關系也無明顯的一致性。這說明與高海拔地區相比,低海拔森林內的氣候條件總體較為適合馬尾松的生長,因此氣候條件的年際間波動對低海拔樹輪寬度影響不大。
3.3 線性混合模型分析
當使用單一氣候因子對單一年表分組(高海拔兩年表/低海拔兩年表)進行擬合的情況下,對高海拔年表輪寬指數的年際波動解釋量最高的氣候因子是生長季當年1月的日照時長(2=0.30),其次是7月的降水量(2=0.20,<0.01)。對低海拔年表輪寬指數年際波動解釋量最高的參數是生長季前一年10月的月均日最高溫(2=0.06)和當年9月的月均最低溫(2=0.04),但>0.05, 均不顯著(表2)。當將上述解釋量較高的氣候要素納入同一線性混合模型(LME)中并將變量個數限定為3個時,最優模型對高海拔年表的解釋量達到了0.50,而對低海拔年表的最高解釋量只有0.13 (圖5)。這說明高海拔馬尾松的徑向生長年際變率中包含了大量的氣候信息,與之相比低海拔馬尾松的徑向生長年際變率中的氣候信息則很少。對模型內不同變量的相對貢獻率分析結果表明(圖6),低海拔氣候-年表混合模型內3個變量的相對貢獻率則沒有顯著差別。而在高海拔模型中,對馬尾松徑向生長解釋量最高的氣候要素是1月日照,相對貢獻率超過了60%,而如果將前一年12月的月均日最高溫一并考慮,則上述2個氣候要素對模型的相對貢獻率大約達到了80%。據此結合前述分析可以推斷,研究區域內高海拔馬尾松徑向生長速率年際波動的主要驅動因子是生長季前冬季的光熱條件,暖冬和長日照可以極大促進馬尾松的生長。
4 結論
全球氣候變化背景下,認識森林生態系統對氣候變化響應模式的空間分布格局對更好地制定相關森林管理和生態保護政策有著重要意義。本研究首次對中國東南部亞熱帶森林中樹木徑向生長與氣候條件年際波動關系的海拔差異進行了研究, 結果表明,武夷山脈東麓高海拔地區的馬尾松徑向生長對氣候要素的年際波動有著較強的敏感性,主要受到生長季前冬季光熱條件的影響,這與此前國內各地區的許多研究結論吻合[14-16,25],說明我國高海拔地區樹木生長與氣候關系在水平方向上異質性較低。與之相比,低海拔地區馬尾松生長對氣候條件年際波動的響應弱,空間差異大。本研究結果表明,武夷山脈東麓高海拔森林的馬尾松生長可能會對全球變化響應更敏感,應該在未來的森林管理中給予更多關注。此外,亞熱帶高海拔森林內生長的樹木對氣候可能有較高的敏感度,具有樹輪氣候學研究的潛在價值。

表2 單氣象要素混合模型擬合結果

圖5 對高低海拔年表進行線性混合效應模型擬合的預測值和觀測值散點圖
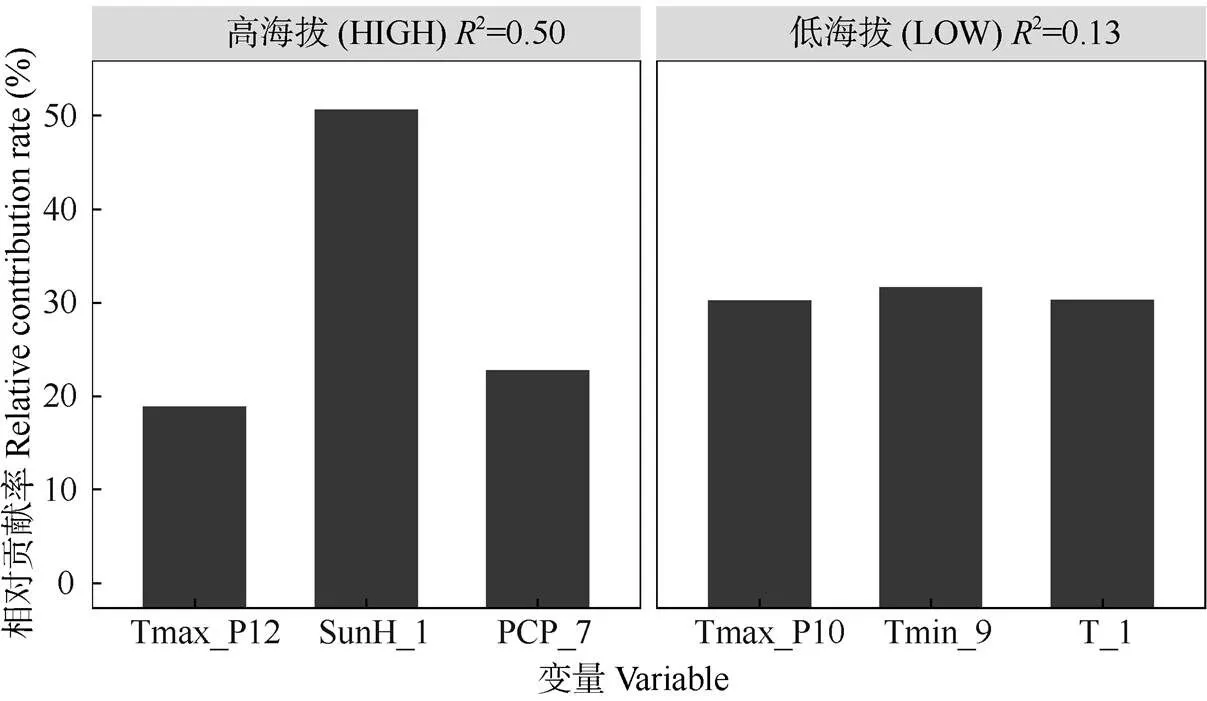
圖6 線性混合效應模型LME內變量的相對貢獻率
[1] YIN Y H, MA D Y, WU S H. Climate change risk to forests in China associated with warming [J]. Sci Rep, 2018, 8: 493. doi: 10.1038/s 41598-017-18798-6.
[2] PEROS M C, GAJEWSKI K, VIAU A E. Continental-scale tree popu- lation response to rapid climate change, competition and disturbance [J]. Glob Ecol Biogeogr, 2008, 17(5): 658-669. doi: 10.1111/j.1466- 8238.2008.00406.x.
[3] KHANDURI V P, SHARMA C M, SINGH S P. The effects of climate change on plant phenology [J]. Environmentalist, 2008, 28(2): 143-147. doi: 10.1007/s10669-007-9153-1.
[4] HUANG J G, TARDIF J C, BERGERON Y, et al. Radial growth response of four dominant boreal tree species to climate along a latitudinal gradient in the eastern Canadian boreal forest [J]. Glob Change Biol, 2010, 16 (2): 711-731. doi: 10.1111/j.1365-2486.2009.01990.x.
[5] JIANG X Y, HUANG J G, STADT K J, et al. Spatial climate- dependent growth response of boreal mixed wood forest in western Canada [J]. Glob Planet Change, 2016, 139: 141-150. doi: 10.1016/j. gloplacha. 2016.02.002.
[6] dos SANTOS V A H F, FERREIRA M J, RODRIGUES J V F C, et al. Causes of reduced leaf-level photosynthesis during strong El Ni?o drought in a Central Amazon forest [J]. Glob Change Biol, 2018, 24(9): 4266-4279. doi: 10.1111/gcb.14293.
[7] SLIK J W F. El Ni?o droughts and their effects on tree species compo- sition and diversity in tropical rain forests [J]. Oecologia, 2004, 141(1): 114-120. doi: 10.1007/s00442-004-1635-y.
[8] WILLIAMSON G B, LAURANCE W F, OLIVEIRA A A, et al. Ama- zonian tree mortality during the 1997 El Ni?o drought [J]. Conserv Biol, 2000, 14(5): 1538-1542. doi: 10.1046/j.1523-1739.2000.99298.x.
[9] NAKAGAWA M, TANAKA K, NAKASHIZUKA T, et al. Impact of severe drought associated with the 1997-1998 El Ni?o in a tropical forest in Sarawak [J]. J Trop Ecol, 2000, 16(3): 355-367. doi: 10.1017/ S0266467400001450.
[10] FANG K Y, GOU X H, CHEN F H, et al. The advance of dendro- ecology [J]. J Glaciol Geocryol, 2008, 30(5): 825-834. 方克艷, 勾曉華, 陳發虎, 等. 樹輪生態學研究進展 [J]. 冰川凍土, 2008, 30(5): 825-834.
[11] MARTIN-BENITO D, PEDERSON N, K?SE N, et al. Pervasive effects of drought on tree growth across a wide climatic gradient in the temperate forests of the Caucasus [J]. Glob Ecol Biogeogr, 2018, 27 (11): 1314-1325. doi: 10.1111/geb.12799.
[12] CHRISTIE D A, LARA A, BARICHIVICH J, et al. El Ni?o-Southern Oscillation signal in the world’s highest-elevation tree-ring chrono- logies from the Altiplano, central Andes [J]. Palaeogeogr Palaeocl Palaeoecol, 2009, 281(3/4): 309-319. doi: 10.1016/j.palaeo.2007.11.013.
[13] SAVVA Y, OLEKSYN J, REICH P B, et al. Interannual growth response of Norway spruce to climate along an altitudinal gradient in the Tatra Mountains, Poland [J]. Trees, 2006, 20(6): 735-746. doi: 10.1007/s00 468-006-0088-9.
[14] ZHANG H, SHAO X M, ZHANG Y. Research progress on the response of radial growth to climatic factors at different altitudes [J]. J Earth Environ, 2012, 3(3): 845-854. 張慧, 邵雪梅, 張永. 不同海拔高度樹木徑向生長對氣候要素響應的研究進展 [J]. 地球環境學報, 2012, 3(3): 845-854.
[15] KANG Y X, LIU J H, DAI S F, et al. Characteristics of ring-width chronologies ofand their responses to climate change at different elevations in Taibai Mountain [J]. J NW Agric For Univ (Nat Sci), 2010, 38(12): 141-147. 康永祥, 劉婧輝, 代拴發, 等. 太白山不同海拔太白紅杉年輪生長對氣候變化的響應 [J]. 西北農林科技大學學報(自然科學版), 2010, 38(12): 141-147.
[16] LEI J P, XIAO W F, HUANG Z L, et al. Responses of ring width ofto the climate change at different elevations in Zigui County, Three-Gorge Reservoir area [J]. Sci Silv Sin, 2009, 45(2): 33- 39. 雷靜品, 肖文發, 黃志霖, 等. 三峽庫區秭歸縣不同海拔馬尾松徑向生長對氣候的響應 [J]. 林業科學, 2009, 45(2): 33-39.
[17] LYU L, DENG X, ZHANG Q B. Elevation pattern in growth cohe- rency on the southeastern Tibetan Plateau [J]. PLoS One, 2016, 11(9): e0163201. doi: 10.1371/journal.pone.0163201.
[18] ZHOU Z X. Masson Pine in China [M]. Beijing: China Forestry Press, 2001. 周政賢. 中國馬尾松 [M]. 北京: 中國林業出版社, 2001.
[19] HOU H Y. Vegetation of China with reference to its geographical distribution [J]. Ann Miss Bot Gard, 1983, 70(3): 509-548. doi: 10. 2307/2992085.
[20] HOLMES R L, ADAMS R K, FRITTS H C. Tree-ring chronologies of western North America: California, eastern Oregon and northern Great Basin, with procedures used in the chronology development work, including users manuals for computer programs COFECHA and ARSTAN [R]. Tucson: Laboratory of Tree-Ring Research, University of Arizona, Tucson. Chronol Ser No VI, 1986.
[21] COOK E R, HOLMES R L. Users manual for program ARSTAN [R]. Tucson: Laboratory of Tree-Ring Research, University of Arizona, Tucson, USA, 1986.
[22] ZANG C. R: The bootRes package for response and correlation function analysis Dendrochronologia [CP]. Franco: Franco Biondi Dendroclimatic Calibration, 2012. doi:10.1016/j.dendro.2012.08.001.
[23] LI D W, FANG K Y, LI Y J, et al. Climate, intrinsic water-use efficiency and tree growth over the past 150 years in humid subtropical China [J]. PLoS One, 2017, 12(2): e0172045. doi: 10.1371/journal. pone.0172045.
[24] CHEN F, YUAN Y J, YU S L, et al. Influence of climate warming and resin collection on the growth of Masson pine () in a subtropical forest, southern China [J]. Trees, 2015, 29(5): 1423-1430. doi: 10.1007/s00468-015-1222-3.
[25] LI L L, SHI J F, HOU X Y, et al. High altitudeHayata growth response to climate in Jiulongshan and Guniujiang, southeastern China [J]. Chin J Appl Ecol, 2014, 25(7): 1849-1856. 李玲玲, 史江峰, 侯鑫源, 等. 中國東南高海拔黃山松生長對氣候的響應——以浙江省九龍山和安徽省牯牛降為例 [J]. 應用生態學報, 2014, 25(7): 1849-1856.
[26] BATES D M, M?CHLER M, BOLKER B M, et al. Fitting linear mixed-effects models using lme4 [J]. J Statis Software, 2015, 67(1): 1-48. doi:10.18637/jss.v067.i01.
[27] WINTER B. Linear models and linear mixed effects models in R with linguistic applications [M/OL]. arXiv:1308.5499, 2013.
[28] Gr?mping U. Relative importance for linear regression in R: The Package relaimpo [J]. J Stat Software, 2006, 17(1): 1-27.
[29] FRITTS H C, SHATZ D J. Selecting and characterizing tree-ring chro- nologies for dendroclimatic analysis [J]. Tree-Ring Bull, 1975, 35: 31-40.
[30] ENSMINGER I, SCHMIDT L, LLOYD J. Soil temperature and intermittent frost modulate the rate of recovery of photosynthesis in Scots pine under simulated spring conditions [J]. New Phytol, 2008, 177(2): 428-442. doi: 10.1111/j.1469-8137.2007.02273.x.
[31] ROSSI S, ANFODILLO T, ?UFAR K, et al. Pattern of xylem phenology in conifers of cold ecosystems at the northern Hemisphere [J]. Glob Change Biol, 2016, 22(11): 3804-3813. doi: 10.1111/gcb. 13317.
[32] HUANG J G, GUO X L, ROSSI S. Intra-annual wood formation of subtropical Chinese red pine shows better growth in dry season than wet season [J]. Tree Physiol, 2018, 38(8): 1225-1236. doi: 10.1093/ treephys/tpy046.
[33] PELLERIN M, DELESTRADE A, MATHIEU G, et al. Spring tree phenology in the Alps: Effects of air temperature, altitude and local topography [J]. Eur J For Res, 2012, 131(6): 1957-1965. doi: 10.1007/ s10342-012-0646-1.
[34] ZHANG D P. A preliminary study of the relationships between the activity of stem cambium forming xylem and rainfall and temperature in[J]. J Fujian Coll For, 1994, 14(3): 215-219. 張大鵬. 馬尾松莖木質部產生與水熱關系的初報[J]. 福建林學院學報, 1994, 14(3): 215-219.
[35] TRANQUILLINI W, HAVRANEK W M, ECKER P. Effects of atmo- spheric humidity and acclimation temperature on the temperature response of photosynthesis in youngMill. [J]. Tree Physiol, 1986, 1(1): 37-45. doi: 10.1093/treephys/1.1.37.
[36] TANJA S, BERNINGER F, VESALA T, et al. Air temperature triggers the recovery of evergreen boreal forest photosynthesis in spring [J]. Glob Change Biol, 2003, 9(10): 1410-1426. doi: 10.1046/j.1365-2486. 2003.00597.x.
[37] OLEKSYN J, ZYTKOWIAK R, KAROLEWSKI P, et al. Genetic and environmental control of seasonal carbohydrate dynamics in trees of diversepopulations [J]. Tree Physiol, 2000, 20(12): 837-847. doi: 10.1093/treephys/20.12.837.
[38] ROSSI S, DESLAURIERS A, GRI?AR J, et al. Critical temperatures for xylogenesis in conifers of cold climates [J]. Glob Ecol Biogeogr, 2008, 17(6): 696-707. doi: 10.1111/j.1466-8238.2008.00417.x.
[39] WU S H, JANSSON P E, KOLARI P. The role of air and soil tempe- rature in the seasonality of photosynthesis and transpiration in a boreal Scots pine ecosystem [J]. Agric For Meteorol, 2012, 156: 85-103. doi: 10.1016/j.agrformet.2012.01.006.
Elevational Heterogeneity in Radial Growth-climate Association ofin Southeastern China
LI Jing-ye1,2,3, HUANG Jian-guo1,3*, LIANG Han-xue1,3, JIANG shao-wei1,2,3, Zhou Peng1,2,3, GUO Xia-li1,2,3, LI Jun-tang4
(1. Key Laboratory of Vegetation Restoration and Management of Degraded Ecosystems,Key Laboratory of Applied Botany, South China Botanical Garden, Chinese Academy of Sciences, Guangzhou 510650, China; 2. University of Chinese Academy of Sciences, Beijing 100049, China; 3. Core Botanical Gardens, Chinese Academy of Sciences,Guangzhou 510650, China; 4. State-owned Longmenghe Forestry Station of Xingshan County, Yichang 443700, Hubei, China)
To achieve a better understanding about the elevation heterogeneity in tree growth-climate association in southeastern China, four tree-ring width chronologies offrom eastern foot of Wuyi Mountains in Fujian were established, and the relationship between radial growth and climate factors along elevation were investigated by bootstrapped correlation analysis and linear mixed effect model (LME). The results showed that radial growth of trees at higher elevation had higher sensitivity to climate and higher consistency between sites, mainly reflected as the positive correlations with the winter-spring temperature/sunshine conditions and precipitation in July. Moreover, the linear mixed effect model using three monthly climate factors, such as monthly mean daily maximum temperature in last December, monthly total sunshine hours in current January, and monthly total precipitation in current July, can explain 50% of total variance of radial growth at higher elevation, and the both fronts play a leading role in the model with cumulative relative contribution rate of 80%. So, it was suggested that the pre-growing season temperature/sunshine condition are the main factor regulating tree radial growth at high elevation, and that these trees might have higher sensitivity to future climate changes and forest management policies should take this into account. Moreover, the results showed that the trees at high elevation in subtropical forest had the potential to be used for tree ring-climate reconstruction.
; Radial growth; Subtropical forest; Elevation; Climate, LME model; China
10.11926/jtsb.4038
2019-01-05
2019-02-18
國家自然科學基金項目(41861124001, 31570584, 41661144007); 中國科學院國際合作重點項目(GJHZ1752); 中國科學院百人計劃項目; 廣東自然科學基金項目(2016A030313152)資助
This work was supported by the National Natural Science Foundation of China (Grant No. 41861124001, 31570584, 41661144007); the International Collaborative Key Project of the Chinese Academy of Sciences (Grant No. GJHZ1752); the 100 Talents Program of the Chinese Academy of Sciences, and the Natural Science Foundation in Guangdong (Grant No. 2016A030313152).
黎敬業,碩士研究生,主要從事樹木年輪學和森林生態學研究。E-mail: jingye_li@scbg.ac.cn
Corresponding author. E-mail: huangjg@scbg.ac.cn
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
