IL28RA真核表達載體的構建和對HaCaT細胞遷移能力的影響
2019-12-06 07:20:38尹雪莉張勝權張學軍
安徽醫科大學學報 2019年11期
尹雪莉,張勝權,張學軍
銀屑病(psoriasis,Pso)確切的病因和發病機制至今仍未完全闡明,是皮膚科領域亟待解決的難題之一[1-5]。全基因組關聯分析研究(genome-wide association study,GWAS)成為發現復雜疾病易感基因最常用的方法之一。GWAS發現大量Pso易感基因,其中具有免疫功能的白介素28受體ɑ(IL-28 receptor α,IL28RA)基因為中國漢族人Pso和系統性紅斑狼瘡共有易感基因,也是歐洲人群Pso的易感基因[6-7]。IL28RA在正常皮膚組織呈中度表達,在原代角質形成細胞和永生HaCaT角質形成細胞中高表達[8-9]。既然IL28RA是Pso易感基因,因此,IL28RA基因可能在Pso發生發展中扮演很重要的角色。該研究通過構建pCMV6-IL28RA重組體并轉染HaCaT細胞,建立IL28RA過表達細胞模型進而觀察IL28RA對HaCaT細胞遷移能力的影響。
1 材料與方法
1.1 材料
1.1.1細胞、質粒和細菌 pCMV6質粒(cat. no. PS100001)購自美國OriGene公司;E.coliDH5α感受態細菌由本實驗室保存;HaCaT細胞購自中科院上海細胞庫。
1.1.2主要試劑 瓊脂糖凝膠回收試劑盒購自美國Promega 公司;Lipofectamine 3000 Transfection Reagent購自美國Invitrogen 公司;限制性內切核酸酶HindⅢ、XhoⅠ、T4 連接酶、Trizol 試劑、逆轉錄試劑盒、DNA Marker、SYBR Premix Ex Taq試劑盒購自日本Takara公司;PCR引物及RT-qPCR引物由上海生工生物公司合成;G418購自美國Sigma公司;DMEM 培養基、胰酶和胎牛血清購自美國Gibco公司。
1.2 方法
1.2.1引物設計和HepG2細胞培養及總RNA提取 根據NCBI Genbank 序列提供的人IL28RA mRNA 序列利用primier 5.0 軟件設計引物,上游引物:AGAATGTGACGCTGCTCTCC;下游引物:GCCGGCTCCACTTCAAAAAG。HepG2細胞培養及總RNA提取:復蘇HepG2細胞,將細胞置于37 ℃、5% CO2培養箱中用含10% 胎牛血清和1%的青霉素、鏈霉素雙抗培養基培養,待其融合80%,棄培養基用2 ml PBS洗3次后加入1 ml Trizol試劑,放置于冰上裂解,提取細胞總RNA。
1.2.2總RNA逆轉錄成cDNA和PCR擴增IL28RA 利用逆轉錄試劑盒將上步總RNA逆轉錄成cDNA。該方法分2步:① 逆轉錄加入0.5 μl Oligo dTPrimer,0.5 μl dNTP Mixture,4 μl RNA,無RNase 水4 μl,70 ℃ 15 min 并迅速放置于冰上;② 上述變性反應液中加入5×PrimeScript Buffer 2 μl,RNase Inhibitor 0.25 μl,PrimeScript RTase 0.5 μl,無RNase 水2.15 μl。點動離心,30 ℃、10 min,42 ℃、60 min,70 ℃、15 min。待反應結束后將其放置在-20 ℃冰箱中備用。PCR擴增IL28RA:依次加入2 μl cDNA,10×PCR Buffer 2.5 μl,0.5 μl上游引物,0.5 μl下游引物,MgCl21.5 μl,2.5 μl dNTP Mixture,TaqDNA Polymerase 1 μl,14.5 μl去離子水。94 ℃、5 min,94 ℃、30 s,40 ℃、30 s,72 ℃、2 min,進行35 個循環,72 ℃、5 min。將PCR產物進行瓊脂糖凝膠電泳并利用瓊脂糖凝膠回收試劑盒回收PCR產物。
1.2.3真核表達載體構建 分別用限制性內切酶HindⅢ、XhoⅠ將IL28RA的PCR片段和質粒pCMV6進行雙酶切,純化回收、連接。瓊脂糖凝膠電泳檢測酶切實驗結果并進行膠回收。回收的酶切產物按質粒pCMV6和IL28RA PCR產物1 ∶8的比例加入T4連接酶體系,在15 ℃條件下連接過夜。將連接產物轉化至E.coliDH5α感受態細菌中,并涂布在固體LB 平板(25 μg/ml卡那霉素)上生長,觀察平板菌落并挑取單克隆菌落接種于 LB 液體培養基中進行37 ℃振蕩過夜,用基因組DNA提取試劑盒提取質粒pCMV6-IL28RA,并酶切,電泳鑒定和采取1.5 ml細菌培養物送至上海生工生物公司測序。
1.2.4重組質粒轉染HaCaT細胞 待HaCaT細胞生長良好時,將其種入24孔板,每孔1×105個細胞。待其融合80%時先換成無抗生素無血清的DMEM 并利用LipofectamineTM3000 Transfection Reagent 分別將對照pCMV6 和重組IL28RA 質粒轉染HaCaT細胞,6 h 后換成完全DMEM 培養基。48 h 后加入終濃度為400 ng/ml 的G418 進行篩選,每2 d 換1次培養基,4周后可觀察到空白對照組細胞全部死亡,而轉染對照質粒組、過表達組細胞瓶中仍有細胞存在,說明穩定轉染成功。細胞培養瓶中維持400 ng/ml G418 培養轉染成功的細胞。
1.2.5RT-qPCR法檢測IL28RA mRNA相對表達水平 將HaCaT未轉染細胞、轉染對照質粒pCMV6 和過表達質粒 pCMV6-IL28RA的細胞分別接種24孔板(細胞接種密度為1×105),當細胞融合70%~80%時,用Trizol試劑提取每種細胞總RNA測定總RNA濃度,根據RT-qPCR說明書進行檢測,甘油醛磷酸脫氫酶(GAPDH)(上游引物:AGATCATCAGCAATGCCTCCTG,下游引物:ATGGCATGGACTGTGGTCATG)作為內參對照。反應體系為20 μl:2×SYBR Mix 10 μl,DEPC H2O 8 μl,cDNA 1 μl,上下游引物分別各0.5 μl。于7500 qPCR 系統:95 ℃預變性5 min、95 ℃變性 15 s、60 ℃退火20 s、72 ℃ 2 min,共40個循環。采用2-ΔΔCt進行兩組間IL28RA mRNA相對表達量計算。
1.2.6傷口愈合法檢測IL28RA對HaCaT細胞遷移能力影響 將HaCaT未轉染細胞,轉染對照質粒pCMV6細胞和過表達pCMV6-IL28RA的細胞分別種于12孔培養板中以5000細胞/ml鋪板兩塊。孵育細胞24 h,使用10 μl無菌移液管尖進行劃痕,產生匯合單層傷口,形成傷口后分別在0 h和48 h拍攝照片。

2 結果
2.1 IL28RA基因編碼序列的獲得從HepG2細胞中提取RNA進行逆轉錄,然后利用引物擴增出 IL28RA的編碼序列,全長1 563 bp,見圖1A。
2.2 IL28RA真核表達載體的構建及鑒定獲取IL28RA基因全長后,經酶切、連接、轉化。挑取單克隆菌落進行PCR及擴大培養后進行酶切并測序。酶切質粒得到5 000 bp左右pCMV6條帶及1 563 bp左右的IL28RA 條帶,見圖 1B。經NCBI blast 比對證明質粒測序結果正確,見圖1C。
2.3 RT-qPCR法分析轉染后IL28RA mRNA在HaCaT中表達情況及分析其對HaCaT細胞遷移能力影響將未轉染HaCaT細胞、轉染對照質粒pCMV6細胞和過表達pCMV6-IL28RA的細胞接種12孔板,至90%融合時,收集細胞并提取細胞RNA,結果顯示與對照組pCMV6相比,過表達組IL28RA mRNA相對表達水平(0.751 7±0.050 7)高于pCMV6對照組(4.442 0±0.621 1),差異有統計學意義(t=4.032,P<0.05)。細胞傷口愈合實驗表明,過表達IL28RA組(68.543 3±3.946 1)48 h細胞遷移未愈合面積百分比大于pCMV6組(42.786 7±2.049 7)和HaCaT組(50.913 3±2.413 6),差異有統計學意義(F=79.01,P<0.01),同時也表明IL28RA基因過表達使得HaCaT細胞遷移減慢。見圖2。
3 討論
Pso是一種以表皮角質形成細胞過度增殖為特征的慢性炎癥性皮膚病,影響世界總人口數2%左右[10]。GWAS發現IL28RA是中國漢族人Pso易感基因,但該基因在Pso中發病機制尚不明確[7]。IL28RA基因的蛋白質產物屬于Ⅱ類細胞因子受體家族[11]。人類IL28RA在膀胱癌、白血病、乳腺癌、頭頸癌和肺癌組織等中表達,在人類的免疫系統呈中度表達[8]。Meta分析IL28RA基因在癌癥的預后價值,研究[12]顯示IL28RA在癌癥中表達越低,患者預后越差。IL-29、IL-28A、IL-28B結合到該受體上,能誘導構象變化促使其與另一個受體IL10R2形成受體配體復合物,激活激酶/信號轉導與轉錄因子信號通路,發揮干擾素λ抗病毒、抗增殖功效[9]。Dumoutier et al[13]研究顯示IL-29抗增殖及抗病毒作用取決于IL28RA絡氨酸343和517兩個位點,當受體兩位點處絡氨酸突變為苯丙氨酸,IL-29抗增殖及抗病毒作用消失。因此,IL28RA在IL-29抗病毒和抗增殖中起著決定性作用。由此可見,IL28RA可能對細胞的功能有重要的調控作用(如增殖、分化、凋亡等),還可能參與機體的免疫活動調節。

圖1 IL28RA 基因的擴增及重組質粒構建與鑒定
A:IL28RA基因 PCR擴增圖;B:IL28RA基因重組質粒酶切鑒定圖;C:重組質粒部分測序結果;M:DNA Marker;1:IL28RA基因全長 PCR 產物;2:雙酶切IL28RA 重組質粒;3:雙酶切 pCMV6質粒
本研究旨在通過構建IL28RA基因真核表達載體,轉染使其在人永生皮膚角質形成細胞HaCaT中過表達,觀察過表達IL28RA基因對HaCaT細胞遷移能力影響。首先利用基因重組技術將IL28RA基因編碼區插入質粒pCMV6中,并用RT-qPCR檢測轉染后IL28RA基因在HaCaT細胞中的mRNA相對表達量,結果顯示過表達IL28RA成功轉染HaCaT細胞;隨后用細胞傷口愈合實驗檢測IL28RA基因過表達對細胞遷移的影響,結果發現過表達IL28RA細胞,48 h后細胞遷移未愈合面積百分比大于pCMV6組和HaCaT組即與對照細胞相比,過表達IL28RA能抑制HaCaT細胞遷移。本研究中IL28RA作為細胞膜上受體,需要相應的配體刺激(如IL-29)進而發揮其功能。然而在傷口愈合實驗中,加入終濃度為100 ng/ml IL-29處理細胞,同時增加IL-29濃度和作用時間(圖2),但是IL-29加入并未改變IL28RA對傷口愈合面積的影響,其中具體機制還需要進一步研究。本研究為進一步探討IL28RA生物學功能及其在Pso中發病機制研究奠定了基礎。
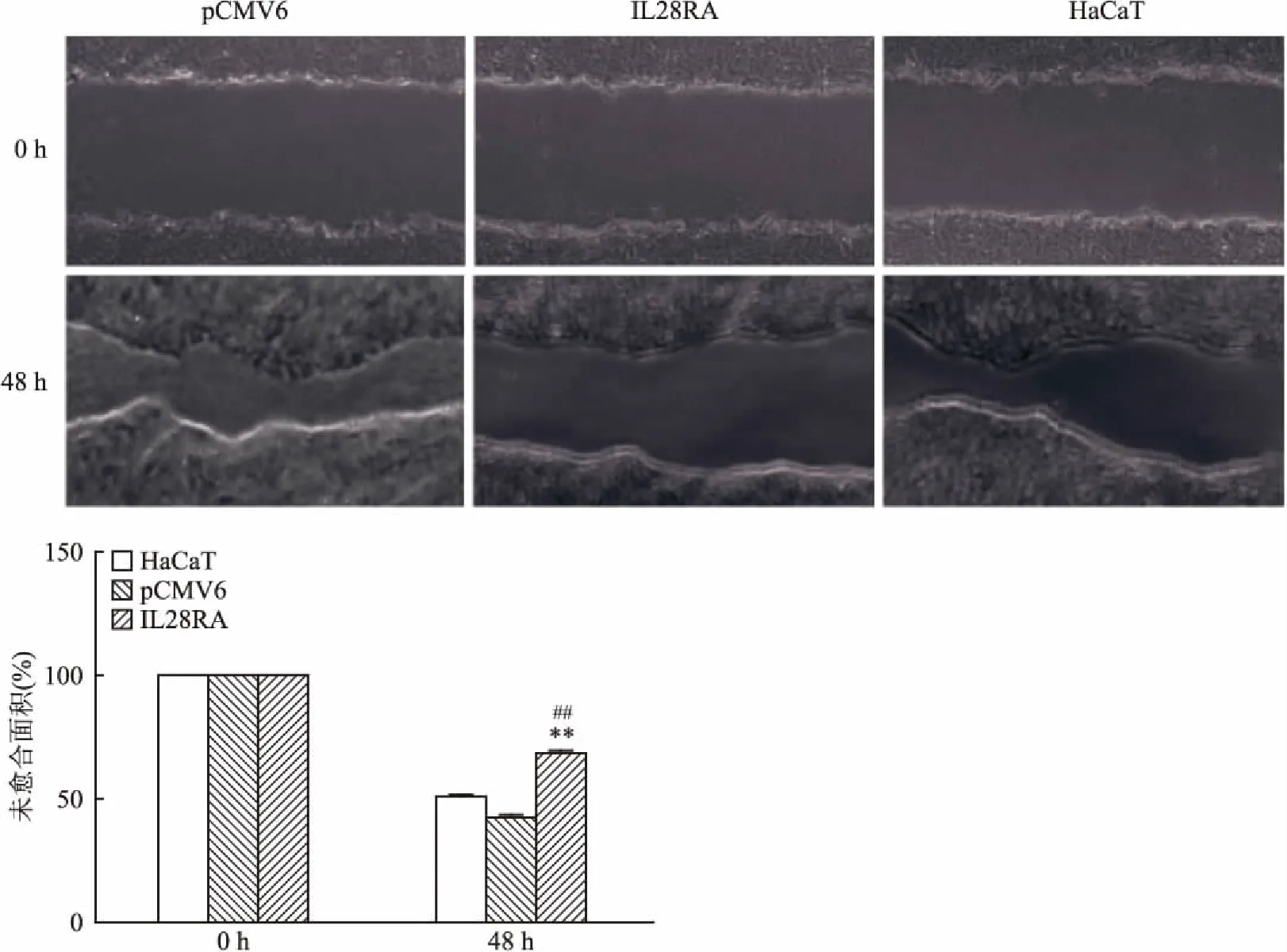
圖2 傷口愈合實驗檢測IL28RA基因過表達對HaCaT細胞遷移能力的影響
與pCMV6組比較:**P<0.01;與未轉染HaCaT組比較:##P<0.01
