不同來源食源性致病菌脂多糖誘導巨噬細胞炎癥相關因子表達研究
2019-12-09 07:34:22南京市食品藥品監(jiān)督檢驗院揚州大學食品科學與工程學院
食品安全導刊 2019年27期
□ 蔣 卉 南京市食品藥品監(jiān)督檢驗院 萬 凱 揚州大學食品科學與工程學院
革蘭氏陰性食源性致病菌作為最常見的病原菌,已成為嚴重威脅到食品安全和公眾健康。雖然高溫高壓可以殺死細菌,但細菌內(nèi)毒素(脂多糖,LPS)不會隨著細菌的死亡而消失或被破壞,細菌死亡后釋放的LPS仍可能威脅人類健康。LPS由內(nèi)至外由3個部分共價連接:疏水的類脂A、核心寡糖和親水的O-抗原。LPS引起炎癥反應,可導致多種感染或致命的膿毒性休克綜合癥[1-3]。研究表明,脂多糖可誘導巨噬細胞炎癥反應[4-5],促進炎癥相關因子表達。但是不同種類革蘭氏陰性菌的LPS結(jié)構(gòu)不同,其免疫活性也有很大差別[6-8]。本文通過考察大腸桿菌O26:B6、腸炎沙門氏菌、銅綠假單胞菌10來源的LPS誘導Raw264.7細胞炎癥相關因子表達的情況,對不同來源食源性致病菌LPS的免疫活性進行比較研究。
1 儀器與試劑
1.1 主要實驗儀器
生物安全柜(美國Thermo公司);3111型二氧化碳培養(yǎng)箱(美國Thermo公 司 );CFX96 Optics Module型擴增儀(美國BIO-RAD公司);UV5Nano超微量分光光度計(瑞士METTLER TOLEDO公司);Allegra X-30R離心機(美國BEXKMAN 公 司 );SpectraMaxμ M3多功能酶標儀(美國Molecular Devices公司);Axio Vert.A1倒置式顯微鏡(德國Carl Zeiss公司)。
1.2 主要實驗試劑
Raw264.7小鼠單核巨噬細胞白血病細胞購自中國科學院細胞庫;DMEM 培 養(yǎng) 基(C11995500BT)、胎牛血清(10099141C)購自美國Gibco公司;大腸桿菌O26:B6來源LPS(L2654)、腸炎沙門氏菌來源LPS(L7770)、銅綠假單胞菌10來源LPS(L9143)購自美國Sigma公司;Prime Script? RT reagent Kit with gDNA Eraser(RR047A)、TB Green ? Premix Dimer Eraser?(RR091A)購自Takara公司;總RNA提取試劑(B511321)購自生工生物工程(上海)股份有限公司;CCK-8試劑(C0037)、一氧化氮檢測試劑盒(S0021)購自碧云天生物技術有限公司;IL-1β、IL-6、TNF-α、IL-1α、iNOS和β-actin基因引物由南京金斯瑞生物工程有限公司合成。
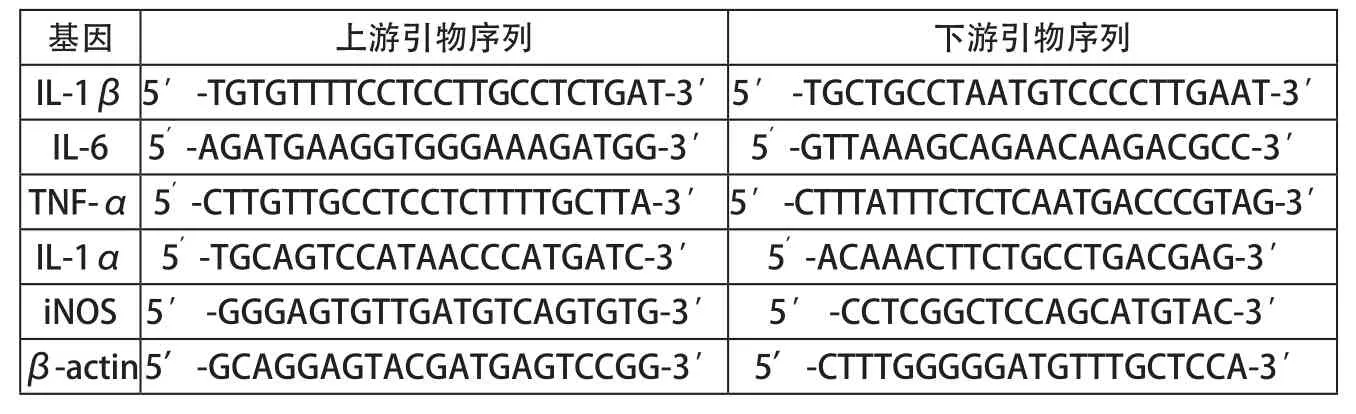
表1 目的基因及內(nèi)參的引物序列表
2 方法
2.1 細胞培養(yǎng)
細胞培養(yǎng)液:90% DMEM,10%(v/v)胎牛血清。Raw264.7細胞于37 ℃、5% CO2條件下培養(yǎng)。
2.2 細胞活性實驗
將細胞接種于96孔板,每孔5 000個細胞。分別給予0(對照組)、0.001、0.01、0.1、1、10、100、1 000 ng/mL與10 000 ng/mL的大腸桿菌O26:B6、腸炎沙門氏菌、銅綠假單胞菌10來源LPS,于37 ℃、5% CO2的條件下處理24 h。提前4 h加入CCK-8溶液,使用酶標儀在450 nm處測量吸光度。實驗平行測定3次。
2.3 細胞中IL-1β、IL-6、TNF-α、IL-1α和iNOS mRNA的檢測
采用RT-qPCR法檢測細胞中的炎癥相關因子IL-1β、IL-6、TNF-α、IL-1α和iNOS mRNA的表達。相關基因引物序列見表1。
將細胞分為對照組(不加LPS)、大腸桿菌O26:B6來源LPS組(LPS終濃度0.1、1、10 μg/mL)、腸炎沙門氏菌來源LPS組(LPS終濃度0.1、1、10 μg/mL)、銅綠假單胞菌10來源LPS組(LPS終濃度0.1、1、10 μg/mL),每組均設置3個復孔,于37 ℃、5% CO2條件下處理6 h后倒掉孔板中的培養(yǎng)液液,收集孔板底部的細胞,用于提取總RNA。細胞總RNA按照UNlQ-10柱式Trizol總RNA抽提試劑盒說明操作,用UV5Nano超微量分光光度計測定總RNA純度和濃度。將符合A260/A280=1.8-2.0的RNA樣品進行逆轉(zhuǎn)錄,逆轉(zhuǎn)錄反應按照逆轉(zhuǎn)錄試劑盒說明操作。將合成得到的cDNA作為模板,進行PCR反應。PCR反應的儀器設置參數(shù):Stage 1:預變性95 ℃、30 s循環(huán)1次;Stage 2:PCR 反應 95 ℃、5 s,55 ℃、30 s,72 ℃、30 s循環(huán)40次;Dissociation Stage:95 ℃、15 s,60 ℃、30 s,95 ℃、15 s循環(huán)1次。反應結(jié)束后確認RT-qPCR的擴增曲線和溶解曲線,根據(jù)公式計算出目標基因的相對表達水平,計算公式[9]參考式(1)。

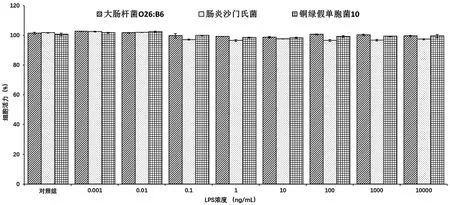
圖1 不同來源LPS刺激下Raw264.7細胞的存活率圖
2.4 NO的檢測
將細胞接種于96孔板,分別給予0(對照組)、0.001、0.01、0.1、1、10、100、1 000、10 000 ng/mL 的大腸桿菌O26:B6、腸炎沙門氏菌、銅綠假單胞菌10來源LPS于37 ℃、5%CO2條件下處理6 h、24 h,每組均設置3個復孔。采用NO試劑盒檢測細胞上清液中NO穩(wěn)定氧化代謝產(chǎn)物亞硝酸鹽()水平。吸取50 μL上清液于96孔板中,按照一氧化氮檢測試劑盒說明書加入相應試劑,使用酶標儀在540 m處測量各孔吸光度,根據(jù)標準曲線計算出NO的含量。
2.5 統(tǒng)計學方法
采用SPSS統(tǒng)計軟件進行分析,兩組間比較采用獨立樣本t檢驗,p<0.05為差異有統(tǒng)計學意義。
3 實驗結(jié)果
3.1 LPS對Raw264.7細胞存活力的影響
經(jīng) 0.001、0.01、0.1、1、10、100、1 000、10 000 ng/mL的 大 腸桿菌O26:B6、腸炎沙門氏菌、銅綠假單胞菌10來源LPS刺激24 h后,Raw264.7細胞的生長并未受到影響,各組與空白對照組相比均無統(tǒng)計學差異(見圖1)。因此,所選LPS濃度可用于后續(xù)實驗。
3.2 不同來源食源性致病菌LPS誘導Raw264.7細胞表達IL-1β、IL-6、TNF-α、IL-1α、iNOS mRNA的情況
如圖2所示,與對照組相比,經(jīng)各濃度大腸桿菌O26:B6、腸炎沙門氏菌來源LPS刺激6 h后炎癥因子IL-1β、TNF-α、IL-1α、iNOS mRNA的表達量均有顯著升高(p<0.05,n=3),IL-6 mRNA的表達量有一定程度的升高,但無統(tǒng)計學差異(1 μg/mL和10 μg/mL腸炎沙門氏菌來源LPS處理組除外)。與對照組相比,銅綠假單胞菌10來源LPS刺激6 h后僅有10 μg/mL LPS處理組的iNOS mRNA的表達量有顯著升高(p<0.05,n=3),其余組各種炎癥因子稍有升高,但均無統(tǒng)計學差異。
刺激Raw264.7細胞6 h后,大腸桿菌O26:B6來源LPS組與腸炎沙門氏菌來源LPS組相比,各種炎癥因子mRNA的表達量均無統(tǒng)計學差異。而銅綠假單胞菌10來源LPS組的各種炎癥因子mRNA的表達量(0.1 μg/mL LPS處理組的IL-6 mRNA的表達量除外)均顯著低于大腸桿菌O26:B6來源LPS組、腸炎沙門氏菌來源LPS組(p<0.05,n=3)。
不同來源食源性致病菌LPS誘導Raw264.7細胞表達IL-1β、IL-6、TNF-α、IL-1α和iNOS mRNA的量有所差異。銅綠假單胞菌10來源LPS誘導Raw264.7細胞表達炎癥相關因子的能力明顯低于大腸桿菌O26:B6來源LPS、腸炎沙門氏菌來源LPS。大腸桿菌O26:B6來源LPS與腸炎沙門氏菌來源LPS誘導Raw264.7細胞表達炎癥相關因子的能力無統(tǒng)計學差異。
3.3 不同來源食源性致病菌LPS誘導Raw264.7細胞釋放NO的情況
采用Griess法檢測了分別在大腸桿菌O26:B6、腸炎沙門氏菌、銅綠假單胞菌10來源LPS刺激6 h、24 h下Raw264.7細胞釋放NO的情況。研究結(jié)果如圖3所示,在相同的LPS濃度下,大腸桿菌O26:B6來源LPS與腸炎沙門氏菌來源LPS誘導Raw264.7細胞產(chǎn)生的NO量無統(tǒng)計學差異;大腸桿菌O26:B6、腸炎沙門氏菌來源LPS誘導Raw264.7細胞產(chǎn)生的NO量明顯多于銅綠假單胞菌10來源的LPS(p<0.05,n=3)。隨著刺激時間的延長,Raw264.7細胞NO釋放量增加。刺激6 h時,不同來源LPS誘導Raw264.7細胞釋放NO具有明顯的劑量依賴性(見圖3a)。值得注意的是,刺激24 h時,各濃度大腸桿菌O26:B6、腸炎沙門氏菌來源LPS誘導后Raw264.7細胞的NO釋放量均呈現(xiàn)較高水平,而銅綠假單胞菌10來源LPS誘導Raw264.7細胞釋放NO依然具有明顯的劑量依賴性(見圖3b)。這說明,長時間的LPS刺激可使Raw264.7細胞NO的釋放量穩(wěn)定,且不再與LPS濃度相關。

圖2 不同來源LPS誘導Raw264.7細胞表達炎癥相關因子的情況圖
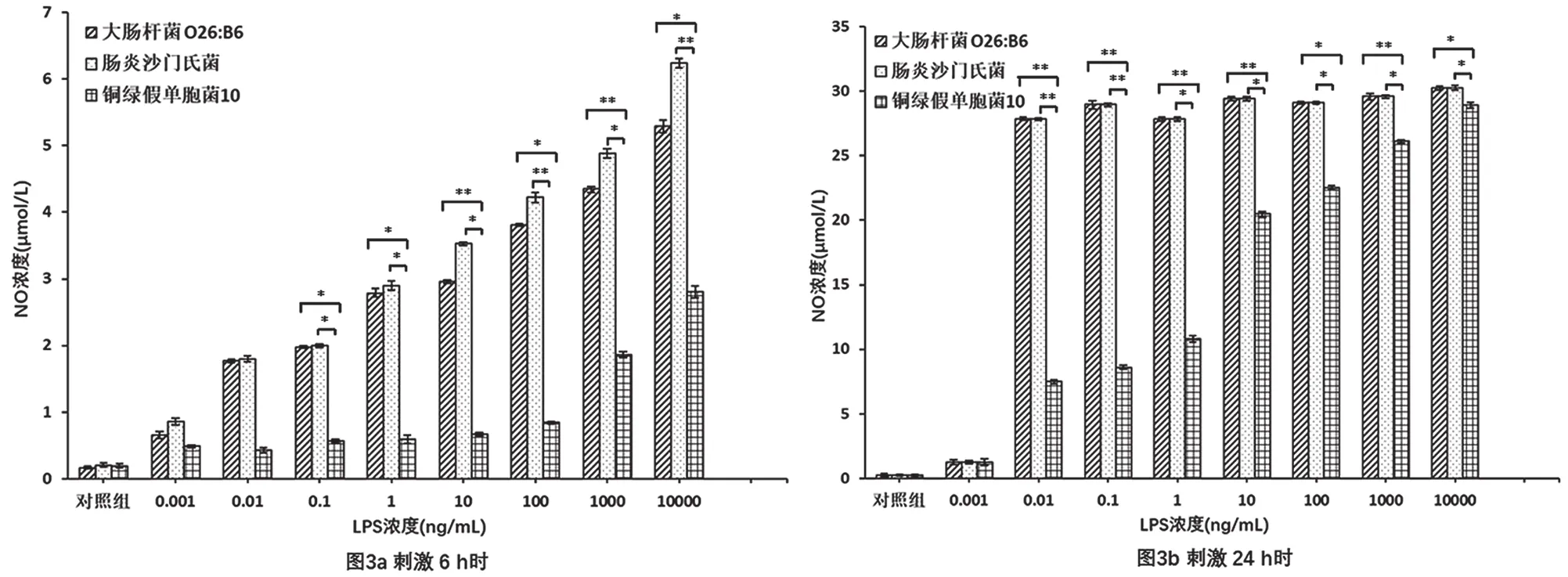
圖3 不同來源LPS誘導Raw264.7細胞釋放NO的情況圖
4 討論
LPS為革蘭氏陰性菌的主要成分,是介導系統(tǒng)性炎癥反應綜合癥的主要啟動因子[10]。巨噬細胞可分為經(jīng)典的M1型、M2型巨噬細胞,還有腫瘤相關巨噬細胞、CD169+巨噬細胞、TCR+巨噬細胞[11]。當細胞接受LPS刺激時,LPS通過巨噬細胞表面的Toll樣受體4(TLR4)來活化核轉(zhuǎn)錄因子KappaB(NF-κB),發(fā)生經(jīng)典的M1型巨噬細胞激活,從而誘導大量的炎癥因子和細胞因子的表達[12],包括TNT-α、IL-6和IL-1α等產(chǎn)生炎癥[13-15],并能夠高表達誘生型一氧化氮酶(iNOS)。
本文選擇Raw264.7巨噬細胞(M1)為細胞模型,初步研究了不同來源食源性致病菌LPS誘導巨噬細胞炎癥相關因子IL-1β、IL-6、TNF-α、IL-1α、iNOS mRNA的表達及NO的釋放情況。結(jié)果表明,銅綠假單胞菌10來源LPS誘導Raw264.7細胞表達炎癥相關因子的能力及NO的釋放明顯低于大腸桿菌O26:B6來源LPS、腸炎沙門氏菌來源LPS。這與革蘭氏陰性菌LPS的結(jié)構(gòu),主要與類脂A的結(jié)構(gòu)有關。類脂A酰化程度對LPS與巨噬細胞中MD-2和TLR4的結(jié)合具有很大的影響[16-17],進而影響LPS的免疫活性。雖然所有革蘭氏陰性菌細胞壁均有LPS,但是脂多糖的結(jié)構(gòu)是不同的。據(jù)文獻報道,大部分腸桿菌科細菌的LPS均含有六個酰基鏈[6],如大腸桿菌、沙門氏菌等。而銅綠假單胞菌10來源的LPS含有五個酰基鏈[7-8]。六酰化(含有六個酰基鏈)類脂A表現(xiàn)出的免疫活性最強[6]。與之相比,五酰化類脂A活性要低100倍,而四酰化類脂A無活性[7-8]。故銅綠假單胞菌10來源的LPS是一種較弱的TLR4受體激動劑,其免疫活性較弱。來自腸桿菌科的食源性致病菌LPS可誘導巨噬細胞炎癥相關因子mRNA的高表達,具有較強的免疫活性。
