尼羅羅非魚淋巴毒素α基因克隆與表達分析
2019-12-17 05:20:18鄒子鴻湯菊芬魯義善簡紀常
廣東海洋大學學報 2019年6期
鄒子鴻,鄭 琦,黃 瑜,湯菊芬,王 蓓,魯義善,簡紀常,蔡 佳
(廣東海洋大學水產(chǎn)學院,廣東省水產(chǎn)經(jīng)濟動物病原生物學及流行病學重點實驗室 暨水產(chǎn)經(jīng)濟動物病害控制廣東省普通高等學校重點實驗室,廣東 湛江 524088)
尼羅羅非魚淋巴毒素α基因克隆與表達分析
鄒子鴻,鄭 琦,黃 瑜,湯菊芬,王 蓓,魯義善,簡紀常,蔡 佳
(廣東海洋大學水產(chǎn)學院,廣東省水產(chǎn)經(jīng)濟動物病原生物學及流行病學重點實驗室 暨水產(chǎn)經(jīng)濟動物病害控制廣東省普通高等學校重點實驗室,廣東 湛江 524088)
【目的】克隆尼羅羅非魚(Oreochromis niloticus)淋巴毒素α 基因(lymphotoxin alpha, LTα),分析該基因的組織分布,探討該基因在抗菌抗病毒免疫過程中的重要作用。【方法】通過RACE 技術(shù)獲得尼羅羅非魚淋巴毒素α 基因(On-LTα)的cDNA 全長,通過生物信息學分析其序列結(jié)構(gòu)特征;利用實時熒光定量PCR 檢測基因的組織分布特征,以及健康尼羅羅非魚感染無乳鏈球菌后該基因的表達變化,并進一步分離出尼羅羅非魚非特異性毒性細胞(NCC),檢測脂多糖(LPS)和聚肌胞苷酸(Poly I:C)刺激后,On-LTα 在NCC 中的表達變化。【結(jié)果】On-LTα 基因序列全長為2 699 bp ,包含705 bp 開放閱讀框(ORF),編碼234 個氨基酸(GenBank 登錄號:MK770358)。該蛋白屬于跨膜蛋白,具有腫瘤壞死因子(TNF)家族保守結(jié)構(gòu)域。實時熒光定量結(jié)果表明,On-LTα在健康尼羅羅非魚11 種組織中均有表達,在脾臟中的表達量最高;無乳鏈球菌刺激后,該基因在脾臟中的表達量顯著升高;LPS 與Poly I:C 刺激后,On-LTα 在NCC 中表達量也顯著上調(diào)。【結(jié)論】On-LTα 參與了尼羅羅非魚抗菌抗病毒的免疫應(yīng)答過程。
尼羅羅非魚;淋巴毒素α 基因;基因克隆;基因表達;非特異性細胞毒性細胞(NCC)
腫瘤壞死因子(Tumor necrosis factor, TNF)超家族由19 個成員組成,TNF 通過與其受體(TNF receptor, TNFR)相互作用可調(diào)節(jié)炎癥和非特異性免疫反應(yīng),在自身免疫性疾病、抵抗細菌和病毒感染中發(fā)揮重要作用[1-2]。淋巴毒素α(LTα)又稱TNF-β,是TNF 超家族中重要成員之一,有膜結(jié)合和游離兩種存在形式,在T 細胞、B 細胞和自然殺傷(Natural killer,NK)細胞等淋巴細胞中均有表達,在免疫調(diào)節(jié)中發(fā)揮不同的作用[3-5]。在抗感染免疫中,LTα通過調(diào)控NF-κB 信號通路與該通路中調(diào)節(jié)蛋白的活性參與炎癥、基質(zhì)降解和細胞凋亡過程[6],還可調(diào)節(jié)樹突狀細胞激活的適應(yīng)性免疫過程[7],以及誘導白介素(IL-6)的表達[8]。LTα 基因缺失會減弱CD8+T 細胞的應(yīng)答,從而不能抑制小鼠中樞神經(jīng)系統(tǒng)中病毒的感染[9]。LTα 在小鼠的NK 細胞前體中表達,并可促進NK 細胞的發(fā)育[10-11]。LTα 缺失會影響NK 細胞的發(fā)育和募集,導致腫瘤生長和轉(zhuǎn)移增強[12]。然而目前LTα 的研究主要集中在哺乳動物中,魚類TNF-β(LTα)基因的報道尚十分匱乏。
尼羅羅非魚(Oreochromis niloticus)是我國重要的淡水經(jīng)濟魚類之一,隨著養(yǎng)殖規(guī)模的擴大以及養(yǎng)殖密度的增加,病害頻發(fā),嚴重影響了羅非魚養(yǎng)殖業(yè)的發(fā)展[13]。腫瘤壞死因子在抵抗細菌和病毒感染中發(fā)揮重要作用,淋巴毒素α 基因是腫瘤壞死因子家族的一員。為了解該基因在尼羅羅非魚抗菌抗病毒過程中的作用,本研究克隆尼羅羅非魚淋巴毒素α 基因(On-LTα),并進行生物信息學分析,檢測On-LTα 在健康魚體中的組織分布及其在無乳鏈球菌、脂多糖(LPS)和聚肌胞苷酸(PolyI:C)刺激后的表達變化,為深入了解該基因在尼羅羅非魚抗感染免疫中的作用提供基礎(chǔ)。
1 材料與方法
1.1 材料
實驗用尼羅羅非魚(50 ± 3)g,購自廣東省高州市某羅非魚養(yǎng)殖場,于24 h 循環(huán)水系統(tǒng)中暫養(yǎng)一周后用于實驗,系統(tǒng)水溫為(28 ± 1)℃。
SMARTer?RACE 試劑盒、pMD18-T 載體、Ex Taq 酶購自TaKaRa 公司。LPS 購自碧云天公司。PolyI:C 購自Apexbio 公司。DH5α 購自武漢轉(zhuǎn)導生物實驗室。TransZol Up Plus RNA Kit、實時熒光定量試劑盒(TransStart Green qPCR SuperMix)購自北京全式金公司。Percoll 分離液購自GE Healthcare公司。無乳鏈球菌(Streptococcus agalactiae)由廣東省水產(chǎn)經(jīng)濟動物病原生物學及流行病學重點實驗室保存。
1.2 方法
1.2.1 總RNA 的提取及cDNA 的合成 取上述暫養(yǎng)活力較好且健康的羅非魚5 尾,剖取腦、心臟、胸腺、肝臟、頭腎、脾臟、腸、腎、鰓、肌肉和皮膚用于組織分布檢測。使用TransZol Up Plus RNA Kit 提取總RNA,參照反轉(zhuǎn)錄試劑盒說明書進行反轉(zhuǎn)錄,獲得 cDNA 一鏈用于后續(xù)實驗。使用SMARTer? RACE 試劑盒合成脾臟cDNA 第一鏈,用于3′ 及5′ RACE。
1.2.2 On-LTα 基因全長cDNA 克隆 通過NCBI數(shù)據(jù)庫預(yù)測得到On-LTα 基因序列,利用PCR 技術(shù)從羅非魚cDNA 中擴增出On-LTα 的開放閱讀框(Open reading frame,ORF),然后設(shè)計全長引物5′RACE 和3′RACE(表1),通過RACE 技術(shù)得On-LTα 基因的cDNA 全長。
1.2.3 On-LTα 基因生物信息學分析 基因序列的比對、ORF 的查找以及同源性分析在NCBI 網(wǎng)站上進行;利用ExPASy (http://web.expasy.org/protparam/)在線分析理化性質(zhì);通過 SignalP 4.1 Server (http:// www.cbs.dtu.dk/services/SignalP/)預(yù)測信號肽;利用SMART(http://smart.embl-heidelberg.de/)在線分析結(jié)構(gòu)域;通過 PSITE(http://linux1.softberry.com/ berry.phtml?topic=psite&group=programs&subgroup=proloc)預(yù)測氨基酸功能位點;二級結(jié)構(gòu)和三級結(jié)構(gòu)的模擬則運用 SOPMA 和SWISS-MOLDEL 軟件完成;使用DNAMAN5.0 軟件進行多序列比對;經(jīng)過ClustalX 比對后的序列利用MEGA5.2 軟件構(gòu)建系統(tǒng)進化樹。
1.2.4 無乳鏈球菌人工感染 將制備好的1 mL 菌液接種于50 mL 的BHI 液體培養(yǎng)基中,置于28 ℃、180 r/min 的搖床中擴培24 h。擴培后用磷酸緩沖液(PBS)將菌液調(diào)整為1×109CFU/mL 后待用。隨機取健康尼羅羅非魚10 尾,分為實驗組和對照組,實驗組每尾注射100 μL 菌液,對照組則注射等量的生理鹽水,注射0、3、6、12、24、48、72 h 后,分別從兩組魚(每組5 尾)的脾臟中提取RNA,反轉(zhuǎn)錄成cDNA 后保存,方法同1.2.1。
1.2.5 尼羅羅非魚頭腎非特異性細胞毒性細胞(NCC)的分離 方法如鄭琦等[14]。取2 g 尼羅羅非魚的頭腎組織,充分破碎后,使用RPMI-1640 培養(yǎng)基配制成細胞懸液,添加至兩層不同濃度Percoll分離液液面,置于水平離心機中以500 g、4 ℃條件離心30 min。
1.2.6 LPS 和Poly(I:C)刺激NCC 將NCC 接種至24 孔細胞培養(yǎng)板中,細胞量約1×105個/孔,分別加入終質(zhì)量濃度為1 mg/mL 的LPS 和5 mg/mL 的Poly(I:C)進行刺激,依次收取刺激0、4、8、12、24 h 后的細胞,提取RNA,反轉(zhuǎn)錄后作為定量模板。
1.2.7 On-LTα 基因表達分析 根據(jù)On-LTα 基因全長序列,用Primer Premier 5.0 設(shè)計引物On-LTα-RT-F、On-LTα-RT-R , 內(nèi)參引物為 On-LTα-actin-F 和On-LTα-actin-R (表1)。反應(yīng)體系為10 μL,其中模板0.5 μL,引物(10 μmol/L)各 0.5 μL,2×SYBR Green Mix (Roche) 5 μL,雙蒸水0.5 μL,每個樣品設(shè)置5個復孔,反應(yīng)條件為:95 ℃,10 s;95 ℃,5 s;58 ℃,15 s;72℃,20 s;循環(huán)35 次。實驗結(jié)果依照 2-ΔΔCt法計算[15],用SPSS 20 進行統(tǒng)計學分析。

表1 引物名稱和序列 Table 1 Primer sequences
2 結(jié)果與分析
2.1 On-LTα 基因cDNA 全長序列及氨基酸序列分析
On-LTα(GenBank 登錄號:MK770358)cDNA序列全長為2 699 bp,ORF 包含705 bp,編碼234個氨基酸(圖1)。理論分子質(zhì)量為21.77 ku,等電點為6.49。該蛋白平均親水值(GRAVY)為-0.194,屬親水性蛋白。SMART 在線分析可知,該蛋白含有TNF 典型結(jié)構(gòu)域(圖2)。TMHMM Server 分析表明,該蛋白在第38 和60 位氨基酸之間存在一個跨膜結(jié)構(gòu)域。通過SignalP 5.0 在線分析可知,該蛋白無信號肽。亞細胞定位預(yù)測主要分布于細胞膜。通過 PSITE 預(yù)測可知,該蛋白存在N-糖基化位點2 個、蛋白激酶C 磷酸化位點6 個、酪蛋白激酶 II 磷酸化位點5 個、酪氨酸激酶磷酸化位點1 個。
2.2 On-LTα 蛋白空間結(jié)構(gòu)預(yù)測
預(yù)測的On-LTα 蛋白空間結(jié)構(gòu)見圖3。由圖3可知,On-LTα 蛋白的234 個氨基酸殘基中,包含33.33%的α 螺旋(藍色h)、20.09%的延伸鏈(紅色e)、11.11%的β 轉(zhuǎn)角(綠色t)以及35.47%的無規(guī)則卷曲(黃色c),其中α 螺旋和無規(guī)則卷曲是該蛋白最大量的結(jié)構(gòu)元件。通過構(gòu)建的蛋白三維立體結(jié)構(gòu)模型(圖4)發(fā)現(xiàn),該蛋白與人類的同源物具有一定保守性。
2.3 同源性比較及進化分析
利用ClustalX 軟件將尼羅羅非魚淋巴毒素α 與底鳉(Fundulus heteroclitus)、斑馬魚(Danio rerio)、人類(Homo sapiens)、南極鱈(Notothenia coriiceps)和大黃魚(Larimichthys crocea)進行多序列比對,結(jié)果顯示,該基因編碼的蛋白在以上物種中相似度不高(圖5)。結(jié)合NCBI 分析發(fā)現(xiàn),尼羅羅非魚On-LTα氨基酸序列與底鳉的相似度最高(50.69%),與南極鱈的相似度(47.35%),與大黃魚的相似度(40.52%),與人類的相似度(27.93%),與斑馬魚的相似度最低(27.27%)。

圖1 尼羅羅非魚On-LTα 蛋白核苷酸及推導的氨基酸序列 Fig.1 Nucleotides and deduced amino acid sequences of protein On-Ltα in Nile Tilapia
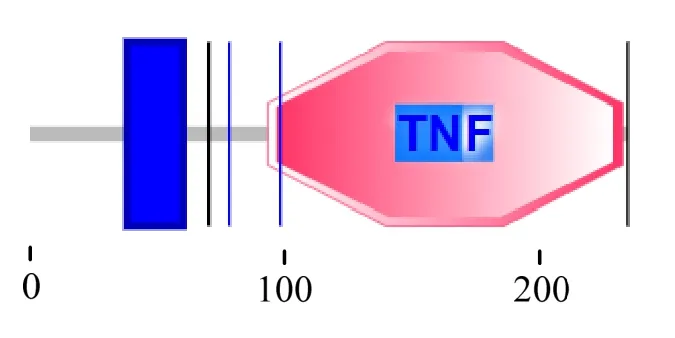
圖2 尼羅羅非魚On-LTα 基因結(jié)構(gòu)域示意圖 Fig.2 Schematic of Nile Tilapia On-LTα

圖3 尼羅羅非魚On-LTα 蛋白二級結(jié)構(gòu)預(yù)測 Fig.3 Predicted secondary structure of Nile Tilapia On-LTα
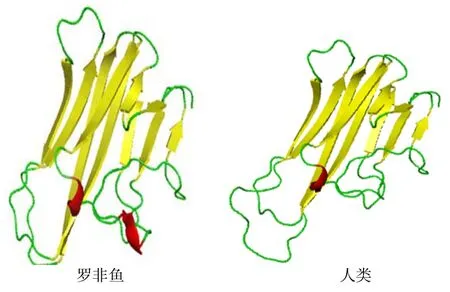
圖4 尼羅羅非魚On-LTα 蛋白與人類On-LTα 蛋白三維結(jié)構(gòu) Fig.4 The three-dimensional structures of On-LTα in Nile Tilapia (Lift) and Homo sapiens (Right)

圖5 尼羅羅非魚LTα 與其他物種LTα 氨基酸序列比對 Fig.5 Multiple alignments of LTα amino acid sequence of Nile Tilapia with other species
對尼羅羅非魚(O.niloticus)、底鳉(F.heteroclitus)、褐家鼠(Rattus norvegicus)、牛(Bos taurus)、金絲猴(Rhinopithecus roxellana)、大棕蝙蝠(Eptesicus fuscus)、西部錦龜(Chrysemys picta bellii)、人(H.sapiens)和大黃魚(Larimichthys crocea)等物種的LTα 基因所編碼氨基酸進行類聚分析,結(jié)果見圖6。圖6 可見,尼羅羅非魚的LTα 與底鳉分類地位最近,列舉物種中魚類全部聚為一支。
2.4 尼羅羅非魚On-LTα 基因的組織分布
組織分布結(jié)果顯示,On-LTα 基因在尼羅羅非魚11 種組織中均有表達,在脾臟中的表達量最高,其次是心臟、肌肉、腸,而肝臟中的表達量最低(圖7)。
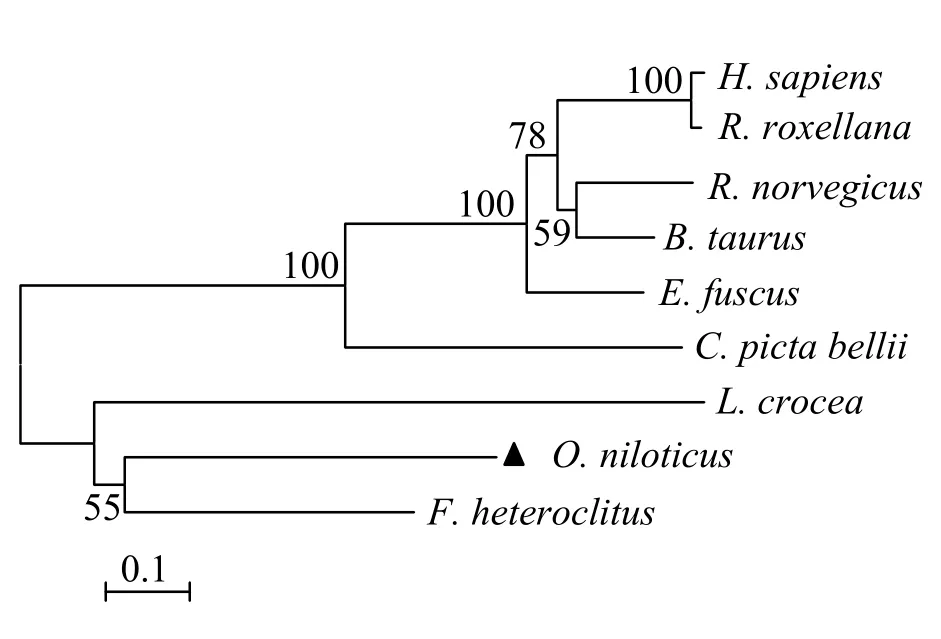
圖6 N-J 法構(gòu)建On-LTα 氨基酸系統(tǒng)進化樹 Fig.6 Phylogenetic tree of On-LTα by neighbour-joining method
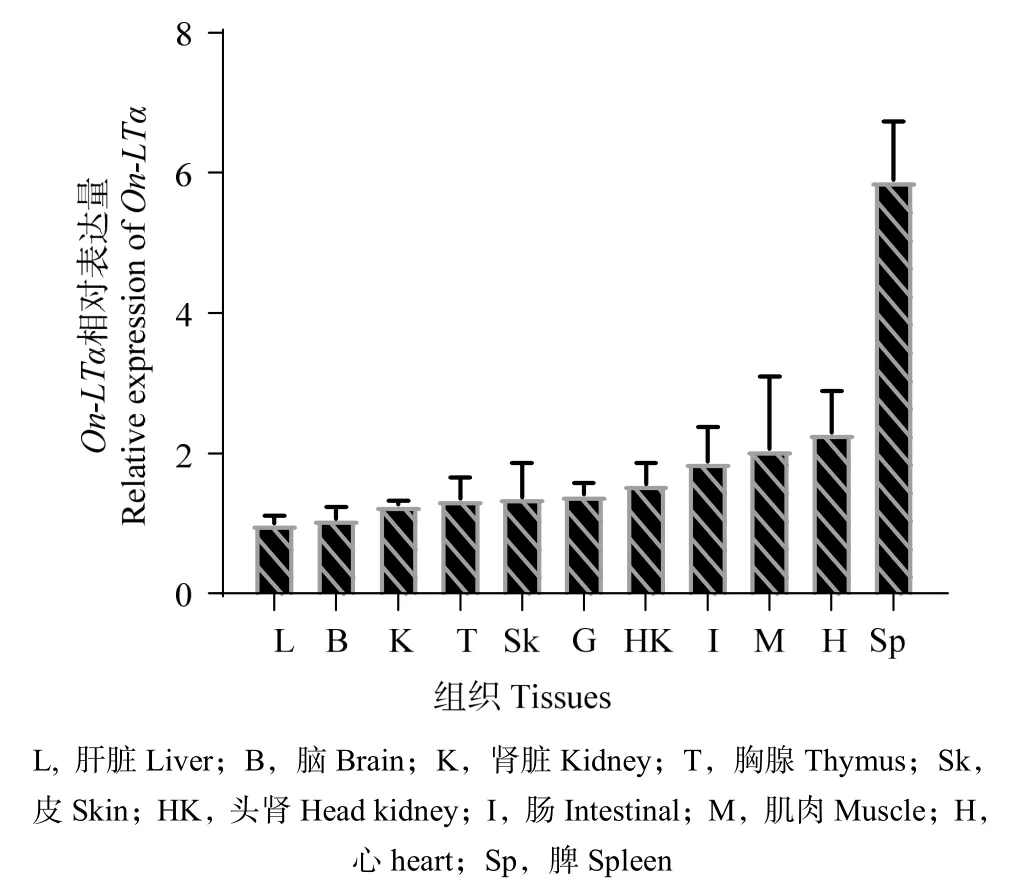
圖7 尼羅羅非魚On-LTα 在不同組織中的表達 Fig.7 The expression of On-LTα of Nile Tilapia in different tissues
2.5 無乳鏈球菌刺激后尼羅羅非魚脾臟中On-LTα表達變化
無乳鏈球菌刺激后,尼羅羅非魚的脾臟On-LTα表達量在3、6、12、48 h 時顯著性(P < 0.05)上 調(diào),其中在3 h 時達到最大值(圖8)。
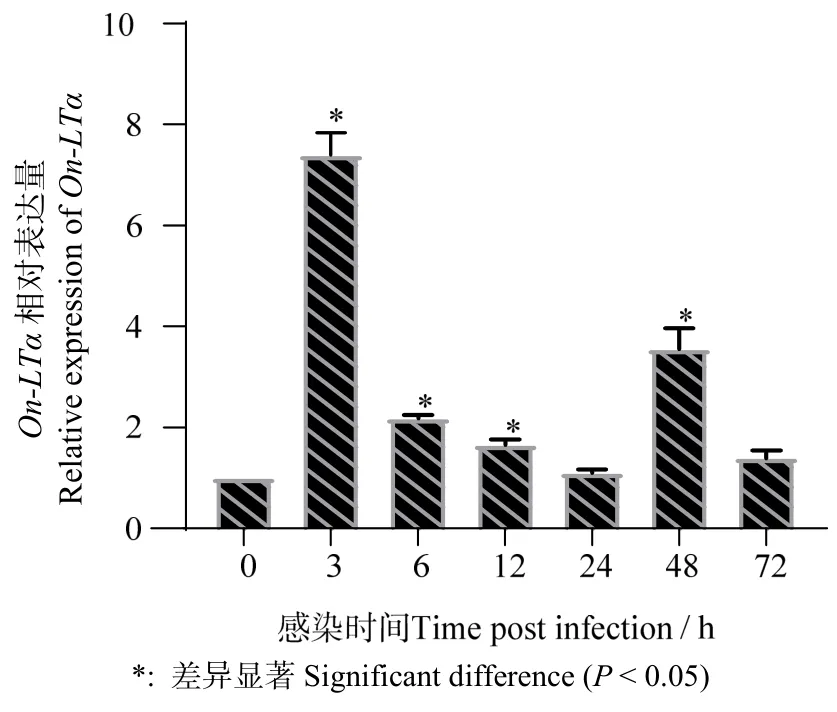
圖8 無乳鏈球菌感染尼羅羅非魚后On-LTα 在脾臟中的表達 Fig.8 Temporal expression of On-LTα in spleen of Nile Tilapia after infected by Streptococcus agalactiae
2.6 LPS、Poly I:C 分別刺激NCC 后On-LTα 的表達
圖9 表明,尼羅羅非魚NCC 在LPS 和PolyI:C刺激后,NCC 中的On-LTα 均在12 h 時表達量明顯上調(diào)。
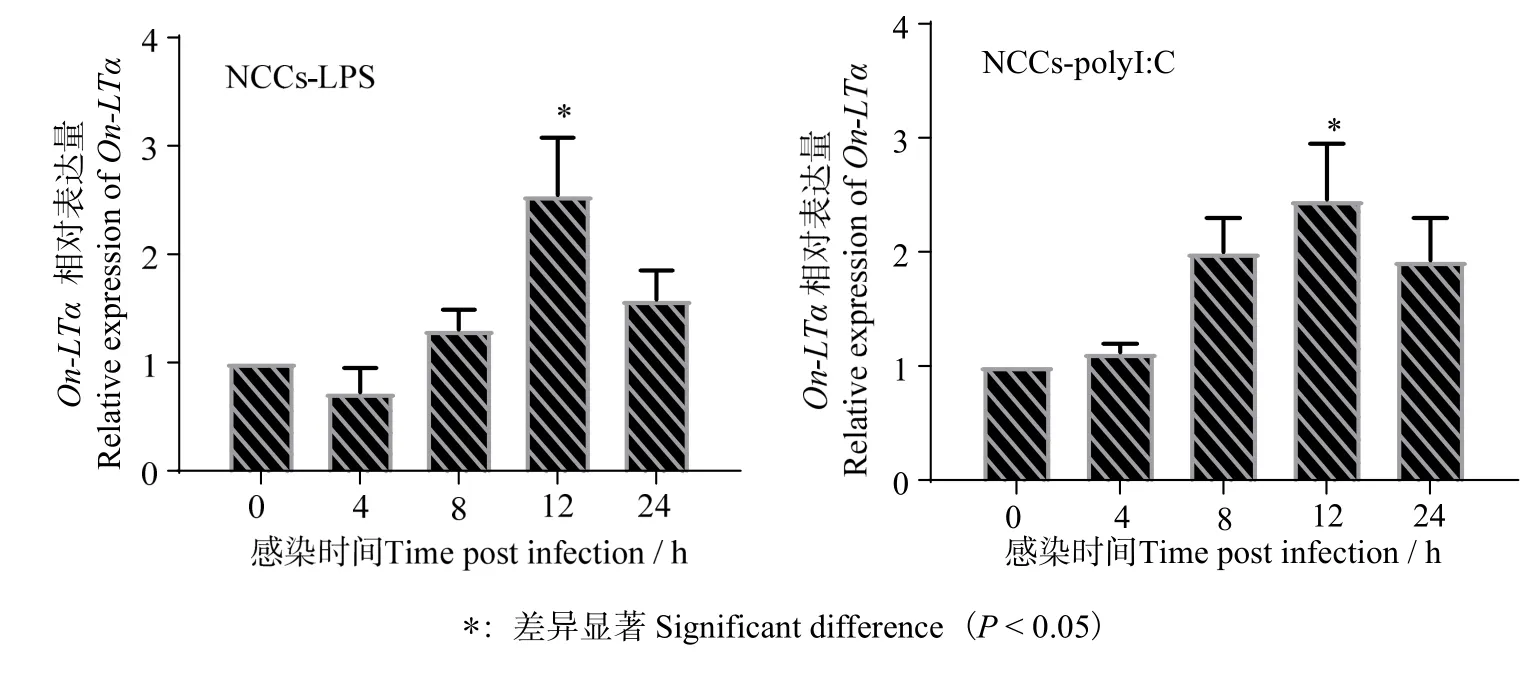
圖9 LPS 和PolyI:C 刺激后On-LTα 基因在NCC 中的表達 Fig.9 Expression of On-LTα in NCC of Tilapia after infected by LPS and PolyI:C
3 討論
本研究克隆得尼羅羅非魚淋巴毒素α 基因cDNA 全長,并推導出編碼On-LTα 的氨基酸序列。分析顯示,On-LTα 序列與其他幾種魚類之間的同源性較低(僅為27% ~ 50%),僅部分氨基酸和功能位點保守,但該基因結(jié)構(gòu)上均有一個特征性的跨膜結(jié)構(gòu)域和典型的TNF 家族保守結(jié)構(gòu)域。而TNF 超家族配體和受體的結(jié)構(gòu)及作用模式相對保守,其同源結(jié)構(gòu)域在不同物種間的保守性也較低(約20% ~ 30%)[16]。
On-LTα 在健康尼羅羅非魚的多種組織中均有表達,且在脾臟的表達量最高。魚類脾臟是二級淋巴器官,On-LTα 對于調(diào)控二級淋巴器官的發(fā)育以及脾臟中T、B 細胞區(qū)域中基質(zhì)細胞的發(fā)育有重要作用[17-18],On-LTα 基因敲除后的小鼠體內(nèi)大部分淋巴結(jié)消失,脾臟結(jié)構(gòu)嚴重紊亂[19-23]。本研究中,On-LTα
無乳鏈球菌是一種廣泛分布于自然界的條件致病菌[24],主要是羅非魚的細菌性病原體。研究表明,無乳鏈球菌滅活疫苗可誘導脾臟腫瘤壞死因子的表達量升高[25]。本研究中,健康尼羅羅非魚感染無乳鏈球菌后,脾臟中On-LTα mRNA 表達量在3 h時顯著上調(diào),說明On-LTα參與了尼羅羅非魚脾臟中的抗菌免疫過程。
NCC 在進化上是自然殺傷細胞(NK)的前體,功能與NK 細胞相似,主要來源于血液和淋巴器官,也是魚類防御病原微生物侵入的第一道防線[26-27],該類細胞激活后可表達腫瘤壞死因子α,通過TNF-α與膜表面受體結(jié)合而發(fā)揮毒性作用,在免疫監(jiān)視、增強魚體非特異性免疫,提高抗病能力方面起重要作用[28]。On-LTα(TNFβ)與TNFα基因密切相關(guān)聯(lián),它們的序列相似(30%),可通過相同的受體發(fā)揮作用[29],且同一種細胞類型可同時表達TNFα和TNFβ,TNFβ還可調(diào)節(jié)TNFα的表達[2]。有研究表明,長牡蠣(Crassostrea gigas)血淋巴細胞中一個有典型TNF 結(jié)構(gòu)域的分子(命名為CgTNF-1),其mRNA表達量在LPS 處理后12 h 顯著增加[16]。本研究中NCC 在經(jīng)LPS 處理后12 h,On-LTα mRNA 表達量也顯著增加,與文獻[16]一致,表明具有TNF 典型結(jié)構(gòu)域的On-LTα可響應(yīng)LPS 處理引起的免疫應(yīng)答。經(jīng)PolyI:C 刺激后12 h,NCC 內(nèi)mRNA 表達量也顯著上調(diào)。由此可見,On-LTα與NCC 的激活存在聯(lián)系,具體機制需進一步研究證實。
本研究成功克隆尼羅羅非魚中的On-LTα基因,并分析在無乳鏈球菌刺激下該基因在脾臟中的表達,以及尼羅羅非魚NCC 經(jīng)LPS 和Poly I:C 刺激后該基因在體內(nèi)或體外的表達。結(jié)果表明,LTα基因參與了尼羅羅非魚抗菌抗病毒的免疫過程。
[1]馮速, 邱樊, 張壯壯, 等.腫瘤壞死因子家族及其相關(guān)藥物的研究進展[J].藥學進展,2016,40(4):284-292.
[2]孫亞薇, 白志超, 朱秋雨, 等.腫瘤壞死因子β及其相關(guān)疾病的研究進展[J].中國醫(yī)藥生物技術(shù),2017,12(4):365-368.
[3]FHU C W, GRAHAM A M, YAP C T, et al.Reed-Sternberg cell-derived lymphotoxin- activates endothelial cells to enhance T-cell recruitment in classical Hodgkin lymphoma[J].Blood,2014,124(19):2973-2982.
[4]RUDDLE N H.Lymphotoxin and TNF: How it all began—a tribute to the travelers[J].Cytokine & Growth Factor Reviews,2014,25(2):83-89.
[5]WARE C F.Network communications: lymphotoxins,LIGHT, and TNF[J].Annual Review of Immunology,2005,23(1):787-819.
[6]BUHRMANN C, SHAYAN P, AGGARWAL B B, et al.Evidence that TNF-β (lymphotoxin α) can activate the inflammatory environment in human chondrocytes[J].Arthritis Research&Therapy,2013,15(6):R202.
[7]GOMMERMAN J L, BROWNING J L,WARE C F.The Lymphotoxin Network: orchestrating a type I interferon response to optimize adaptive immunity[J].Cytokine Growth Factor Rev,2014,25(2):139-145.
[8]LAUENBORG B, CHRISTENSEN L, RALFKIAER U,et al.Malignant T cells express lymphotoxin α and drive endothelial activation in cutaneous T cell lymphoma[J].Oncotarget,2015,6(17):15235-49.
[9]KUMARAGURU U, DAVIS I A, DESHPANDE S, et al.Lymphotoxin alpha-/-mice develop functionally impaired CD8+ T cell responses and fail to contain virus infection of the central nervous system[J].Journal of Immunology,2001,166(2):1066-1074.
[10]KIM T J, UPADHYAY V, KUMAR V, et al.Innate lymphoid cells facilitate NK cell development through a lymphotoxin-mediated stromal microenvironment[J].Journal of Experimental Medicine, 2014, 211(7):1421-1431.
[11]IIZUKA K, CHAPLIN D D, WANG Y, et al.Requirement for membrane lymphotoxin in Natural Killer Cell development[J].Proceedings of the National Academy of Sciences of the United States of America,1999,96(11):6336-6340.
[12]ITO D, BACK T A, WILTROUT R, et al.Mice with a targeted mutation in lymphotoxin-alpha exhibit enhanced tumor growth and metastasis: impaired NK cell development and recruitment.[J].Journal of Immunology,1999,163(5):2809-2815.
[13]黃錦爐.羅非魚無乳鏈球菌病病原學、病理學及cpsE基因的原核表達研究[D].四川:四川農(nóng)業(yè)大學,2012.
[14]鄭琦, 陳一錦, 黃瑜, 等.尼羅羅非魚非特異性細胞毒性細胞(NCC)分離條件的優(yōu)化[J].廣東海洋大學學報,2017,37(3):123-126.
[15]LIVAK K J, SCHMITTGEN T D.Analysis of relative gene expression data using real-time quantitative PCR and the 2 (-Delta Delta C (T)) Method[J].Methods,2001,25(4):402-408.
[16]孫穎.長牡蠣腫瘤壞死因子(CgTNF-1)免疫調(diào)節(jié)機制的初步研究[D].青島: 中國科學院研究生院(海洋研究所),2014.
[17]UPADHYAY V, FU Y X.Lymphotoxin signalling in immune homeostasis and the control of microorganisms[J].Nature Reviews Immunology, 2013,13(4):270-279.
[18]NGO V N, KORNER H, GUNN M D, et al.Lymphotoxin α/β and tumor necrosis factor are required for stromal cell expression of homing chemokines in b and t cell areas of the spleen[J].Journal of Experimental Medicine,1999,189(2):403-412.
[19]BANKS T A, ROUSE B T, KERLEY M K, et al.Lymphotoxin-alpha-deficient mice.Effects on secondary lymphoid organ development and humoral immune responsiveness[J].Journal of Immunology,1995,155(4):1685-93.
[20]ALIMZHANOV M B, KUPRASH D V,KOSCOVILBOIS M H,et al.Abnormal development of secondary lymphoid tissues in lymphotoxin β-deficient mice[J].Proceedings of the National Academy of Sciences of the United States of America,1997,94(17):9302-9307.
[21]MACKAY F, MAJEAU G R, LAWTON P, et al.Lymphotoxin but not tumor necrosis factor functions to maintain splenic architecture and humoral responsiveness in adult mice[J].European Journal of Immunology,1997,27(8):2033-2042.
[22]RENNERT P D, BROWNING J L, MEBIUS R, et al.Surface lymphotoxin α/β complex is required for the development of peripheral lymphoid organs[J].Journal of Experimental Medicine,1996,184(5):1999-2006.
[23]KONI P A, SACCA R, LAWTON P,et al.Distinct roles in lymphoid organogenesis for lymphotoxins alpha and beta revealed in lymphotoxin beta-deficient mice[J].Immunity,1997,6(4):491-500.
[24]劉志剛, 可小麗, 盧邁新, 等.羅非魚無乳鏈球菌強毒株基因組表達文庫的構(gòu)建及鑒定[J].南方水產(chǎn)科學,2015,11(6):34-40.
[25]付天增.無乳鏈球菌和海豚鏈球菌滅活疫苗對羅非魚的免疫效果[D].濟南: 山東農(nóng)業(yè)大學,2017.
[26]魏世娜.紅笛鯛NCCRP-1 基因的克隆和表達分析[D].湛江: 廣東海洋大學,2010.
[27]魏世娜, 簡紀常, 吳灶和, 等.魚類非特異性細胞毒性細胞(NCC)的研究進展[J].中國水產(chǎn)科學, 2010,17(2):374-380.
[28]甘楨.尼羅羅非魚T 細胞激活相關(guān)基因的功能研究[D].湛江: 廣東海洋大學,2015.
[29]AGGARWAL B B,KOHR W J,HASS P E,et al.Human tumor necrosis factor.Production, purification, and characterization.[J].Journal of Biological Chemistry,1985,260(4):2345-54.
Cloning and Expression Analysis of Gene Lymphotoxin α in Nile Tilapia (Oreochromis niloticus)
ZOU Zi-hong, ZHENG Qi, HUANG Yu, TANG Ju-fen, WANG Bei, LU Yi-shan, JIAN Ji-chang, CAI Jia
(Fisheries College of Guangdong Ocean University, Guangdong Provincial Key Laboratory of Pathogenic Biology and Epidemiology for Aquatic Economic Animals & Key Laboratory of Control for Diseases of Aquatic Economic Animals of Guangdong Higher Education Institutes, Zhanjiang 524088, China)
【Objective】 To clone and analyze the lymphotoxin alpha gene (LTα), analyze the transcript distribution in different tissues, and to explore the important role of lymphotoxin alpha (LTα) in the antibacterial and antiviral immunity of Oreochromis niloticus.【Method】The full-length cDNA of lymphotoxin α from Nile Tilapia (On-LTα)was obtained by RACE.The sequence of On-LTα was analyzed by bioinformatics method.The tissue distribution and expression profiles of On-LTα following Streptococcus lactobacillus immunization were investigated by Real-time quantitative PCR.Furthermore, On-LTα transcript levels in the isolated non-specific toxic cells (NCC) after lipopolysaccharide (LPS) and Poly I:C stimulation were also examined.【Results】On-LTα gene sequence is 2 699 bp including a 705 bp open reading frame (ORF) which encodes a polypeptide of 234 amino acids.(GenBank accession number: MK770358).On-LTα protein belonged to the transmembrane protein and possessed a conserved domain of the tumor necrosis factor (TNF) family.Quantification PCR found that On-LTα was constitutively expressed in all 11 tissues tested.In the healthy Nile tilapia, the highest expression being detected in the spleen.The relative expression levels of On-LTα in the spleen were significantly increased during S.agalactia infection.The isolated NCCs cultured in vitro could also induce the significant upregulation of On-LTα under LPS and Poly I:C stimulation.【Conclusion】On-LTα might be involved in the antimicrobial and antiviral immune responses of tilapia.
Oreochromis niloticus; gene lymphotoxin alpha; gene clone; gene expression; Non-specific cytotoxic cell (NCC)
Q954.4
A
1673-9159(2019)06-0001-08
10.3969/j.issn.1673-9159.2019.06.001
鄒子鴻,鄭琦,黃瑜,等.尼羅羅非魚淋巴毒素α基因克隆與表達分析[J].廣東海洋大學學報,2019,39(6):1-8.
2019-06-28
國家自然科學基金(31302226, 31572651);廣東海洋大學優(yōu)秀碩士學位論文培育項目(201833)
鄒子鴻(1994―),男,碩士研究生,研究方向為水生動物免疫學。E-mail: 1220140057@qq.com
蔡佳,男,副教授,博士,研究方向為水生動物免疫學。E-mail: matrix924@foxmail.com
(劉慶穎、劉胐)
