基于三維冠層模型的玉米光合作用和光能利用模擬
2019-12-19 01:42:12顧生浩王勇健溫維亮盧憲菊于澤濤郭新宇
農業工程學報 2019年20期
關鍵詞:模型
顧生浩,王勇健,溫維亮,盧憲菊,于澤濤,郭新宇
基于三維冠層模型的玉米光合作用和光能利用模擬
顧生浩,王勇健,溫維亮,盧憲菊,于澤濤,郭新宇※
(1. 北京農業信息技術研究中心,北京 100097;2. 國家農業信息化工程技術研究中心,北京 100097;3. 數字植物北京市重點實驗室,北京 100097)
光線分布和葉片光合特征在冠層內部具有極強的時空異質性,基于三維冠層模型的玉米光合模型是精確評估品種高光效的重要手段。該研究將作物三維冠層模型、光線分布模型、光合模型與光能利用模型相耦合,建立了玉米冠層光合生產模型3DMaizeCaP,設置3個不同株型的玉米品種(矮單268、京科968和鄭單958),2種不同光照條件(晴天和陰天),通過大田試驗與模型模擬研究揭示了玉米冠層光合速率和光能利用效率對品種和環境的響應。結果表明,矮單268、京科968和鄭單958的葉片最大光合速率和暗呼吸速率均隨節位下降呈線性降低的垂直分布規律,各品種中矮單268的最大光合速率最大,而暗呼吸速率最小;冠層凈光合速率日變化趨勢明顯,矮單268在陰天和晴天下的冠層最大凈光合速率(以CO2計)為21.6和26.2mol/(m2·s),均顯著(<0.05)高于京科968(20.8和24.9mol/(m2·s))和鄭單958(19.6和24.4mol/(m2·s));矮單268的日CO2凈同化量在陰天和晴天下均顯著(<0.05)高于鄭單958,增幅分別高達14.8%和12.4%,各品種間株型雖有顯著差異(<0.05),但冠層日累積光截獲并無顯著差異(>0.05);單葉尺度上,各葉片中第16節位的單葉日凈同化量達到最大;矮單268的光能利用效率最大,在陰天和晴天下分別為3.22和3.03 g/MJ,比京科968分別高4.5%和5.6%,比鄭單958分別高7.7%和7.8%;初始光量子效率對玉米冠層光能利用效率的敏感性顯著高于最大光合速率(<0.05)。從提高玉米冠層光能利用效率考慮,建議設計株型緊湊、葉片光合性能強的玉米品種。研究可為定量研究玉米冠層光合速率提供估算方法,也可為高光效品種選育提供評價依據和鑒定技術。
光合作用;作物;模型;冠層光分布;三維點云;植物功能結構模型;光響應曲線;光能利用效率
0 引 言
在全球氣候變化和人口不斷增加的背景下,提高單位面積糧食產量,在有限的耕地資源上實現糧食增產、穩產對于保障糧食安全具有重要意義。作物的產量潛力是單位面積土地接收的光能(photosynthetic absorbed radiation, PAR)、葉冠層對光能的截獲效率、光能利用效率(radiation use efficiency, RUE)和經濟系數的乘積[1]。大量的理論和試驗研究表明,作為決定作物產量潛力的重要因素,冠層RUE目前仍存在較大的提升空間,通過提高作物冠層RUE可實現產量大幅提升[2-4]。RUE是日生物量積累與冠層PAR日累積截獲量之比[5],被APSIM[6]、DSSAT[7]和WOFOST[8]等眾多作物模型用于表征冠層光合效率,并在作物生產力模擬上取得顯著成果[9]。雖然RUE的計算方法簡便、實用,但同時又面臨經驗性較強、機理過度泛化、分辨率較低的問題,這有可能為模型精確模擬帶來高度不確定性[10]。基于冠層光合作用模型計算逐日RUE具有較高的穩定性和分辨率[11],有助于理解葉片光合生理和冠層光能利用之間的聯系,國內外學者圍繞冠層光合作用模型開展了大量的研究工作,并且取得了顯著的成果[12-15]。
目前,葉片光響應曲線到作物冠層光合生產的尺度提升是構建冠層光合作用模型的主要方法,模型構建過程中的關鍵問題之一是如何處理葉片光分布和葉片光合生理特征在冠層內部的空間異質性,采用PAR和光響應曲線特征參數在時間或空間上的均值來測算冠層光合速率容易出現高估的問題[16-18]。根據光分布算法的異同可將作物冠層光合作用模型分為大葉模型(big-leaf model)[19]、多層模型(multilayer model)[20]、陽葉陰葉模型(sun-shade)[21]和植物功能結構模型(functional-structural plant model)[22]。大葉模型將冠層簡化為平展的葉片,多層模型考慮到PAR和光合生理特征的垂直分布將冠層劃分為若干層次,陽葉陰葉模型進一步將冠層中的葉片分為陽葉(sunlit leaves)和陰葉(shaded leaves)。Lindquist等[10]基于大葉模型模擬出生長速率與逐日PAR累積截獲量、累積生物量與生長季PAR累積截獲量,并采用線性回歸方法估算了玉米在適宜種植條件下的RUE。于強等[12]將玉米冠層按葉面積指數劃分成若干層次,并與冠層光分布模型、單葉光合模型相耦合,模擬研究了玉米冠層光合速率對株型和環境的響應。但冠層PAR垂直分布模型用于模擬夏玉米封壟前PAR的垂直分布時存在精度不高的問題[23]。Wu等[24]在區分冠層陽葉和陰葉的基礎上,完成了C4作物葉片光合生理生化模型到作物冠層光合作用模型的尺度提升,并模擬了RUE對環境的響應。盡管陽葉、陰葉模型已經在模擬精度、計算效率、機理性和穩定性方面得到顯著改善,但仍無法考慮株型指標(如葉片大小、形態和葉傾角)和種植模式(如寬窄行、間套作)對消光系數的影響,這在一定程度上限制了冠層光合作用模型的應用[25]。
植物功能結構模型以器官為建模的基本單元,包含更豐富的株型信息,可結合光線追蹤算法計算器官尺度上的光分布狀況[26]。雖然Louarn等[27]利用玉米三維模型ADEL-Maize計算了冠層累積PAR,并結合試驗數據分析了低溫下光能利用效率和光截獲對玉米減產的相對貢獻,但該研究僅將植物功能結構模型用于測算冠層PAR截獲量,缺乏對冠層光合速率、干物質生產的模擬研究。目前,基于植物功能結構模型的冠層光合速率和光能利用效率研究多以水稻[15]和小麥[28]為主,針對玉米開展的研究工作較少。因此,本研究利用株型差異顯著的矮單268(AD268)、京科968(JK968)和鄭單958(ZD958)為研究對象,分析葉片光響應曲線特征參數的空間分布規律,通過葉片光響應曲線的尺度提升建立了一個高精度的冠層光合生產模型3DMaizeCaP,探討了品種和環境對玉米冠層光合速率和光能利用效率的影響。本研究可為定量研究玉米冠層光合速率提供估算方法,也可為高光效品種選育提供評價依據和鑒定方法。
1 材料與方法
1.1 試驗材料與設計
本試驗以AD268、JK968和ZD958為供試材料,于2018年在北京農林科學院院內試驗田(39°56'N,116°16'E)開展。試驗采用隨機區組設計,3個品種,3次重復,共設9個小區。種植密度為6株/m2,等行距種植,行距為60 cm。每個小區長10 m,寬9 m。6月4日澆水后穴播,5葉期定苗,每穴保留1株。試驗地土壤為砂壤土,播種前耕翻15 cm,耕作層田間持水量為32%。耕層土壤有機質質量分數為27.2 g/kg,總氮為1.34 g/kg,速效磷為37.6 mg/kg,速效鉀為91 mg/kg,pH值為7.6。
1.2 玉米光響應特征參數
在抽雄期(播后第65天),每個小區選取1株具有代表性的玉米。取樣前在所選植株莖基部標記指北線,以確定田間方位角。將所選植株以直徑30 cm,深25 cm帶土挖出后放置在花盆中并運回實驗室進行葉片光響應曲線的測定和植株三維數字化。取樣后立即為盆中植株澆水,以保證植株不會因失水造成形態變化。采用英國PP SYSTEM公司生產的CIRAS-2型全自動便攜式光合作用測定系統測定葉片凈光合速率。分別選取上、中、下3個部位的葉片進行光響應曲線的測定,測量時先以LED強光源適應后測量,光強PPFD依次設定為1 600、1 400、1 200、1 000、800、700、600、500、400、300、250、200、150、100、50、0mol/(m2·s),葉室內CO2摩爾濃度控制在380mol/mol。由于葉子飄和于強提出的直角雙曲線修正模型能夠準確、合理地通過曲線擬合得出光響應曲線特征參數[29],研究選取該模型來計算光響應曲線的初始量子效率()和飽和光強下的最大光合速率(max,mol/(m2·s),以CO2計)。
1.3 玉米冠層結構三維重建與冠層光分布計算
利用FastScan獲取玉米株型三維數字化數據,基于三維數字化的玉米株型參數提取方法提取玉米株型參數[30],并將葉片模板添加至玉米器官三維模板資源庫[31]。在此基礎上,結合基于拔節期冠層頂部圖像所提取的冠層內各植株生長位置和植株方位平面信息,利用基于分布函數的玉米群體三維模型構建方法[32],重建包含9株(3行×3列)的玉米冠層結構三維模型。為了避免計算冠層光分布過程中的群體邊際效應,在所重建目標冠層周圍復制中心目標群體,形成81株(9行×9列)的玉米冠層三維模型,并在該冠層模型周圍構建玉米株高75%的虛擬墻,保證晴天和陰天情況下直射光及散射光分布的計算精度。采用基于Z-Buffer的光線投射方法和基于多分辨率細分半球的方法分別計算冠層內各時刻的直射光和散射光分布,并在時間尺度上對光截獲結果積分,得到當天的冠層光截獲總量[33-34]。
1.4 玉米冠層光合作用模型
基于不同節位葉片光響應曲線的特征參數和某一時刻的光截獲強度,使用直角雙曲線模型(式1)[29]來計算第節位上的葉片在時刻的凈光合速率(A,t,mol/(m2·s)),按逐日(式2)進行積分得到不同葉片的日CO2凈同化量(A,DAY,mmol/d),然后將單株所有葉片累加后乘以種植密度,再分別按照逐小時(式3)和逐日(式4)進行積分得到整個冠層凈光合速率的每小時(c,t,mol/(m2·h))和全天CO2凈同化量(c,DAY,mmol/(m2·d))。研究針對不同株型對冠層光合速率的影響,并未考慮葉片氮含量、葉片溫濕度和同化物等因素對光合作用的反饋。
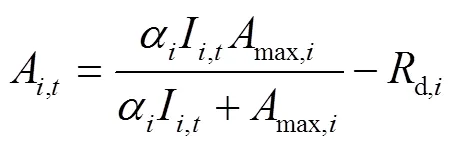
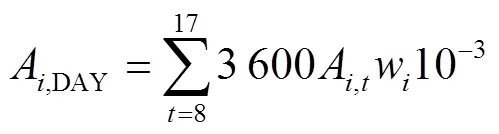
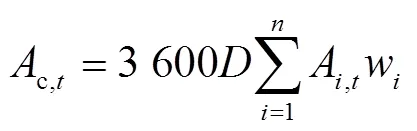
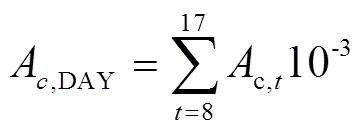
式中A,t是第片葉在時刻的凈光合速率(以CO2計),mol/(m2·s);α是第片葉的初始量子效率,無量綱;I是第片葉在時刻所截獲的光合有效輻射強度,mol/(m2·s);max,i是第片葉的最大凈光合速率,mol/(m2·s);是植株最大葉片數;d,i是第片葉的暗呼吸速率,mol/(m2·s);A,DAY是第片葉的日CO2凈同化量(mmol/d);c,t是第小時玉米冠層CO2凈同化量,mol/(m2·h);w是第片葉的葉片面積,m2;是種植密度,株/m2。c,DAY是玉米冠層全天的CO2凈同化量,mmol/(m2·d)。
1.5 玉米冠層光能利用效率
采用Wu等[24]的方法計算玉米冠層光能利用效率(RUEDAY, g/MJ),其公式如下:
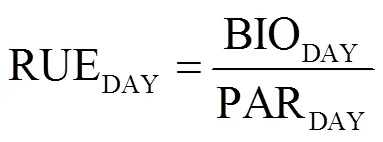
式中BIODAY為日內生物量積累,g/(m2·d);PARDAY為冠層PAR日累積截獲量,MJ/(m2·d)。
為了考慮葉片暗呼吸和植株其他器官的維持呼吸,將葉片總光合速率求和所得冠層總同化量乘以轉化系數即為冠層日內生物量積累[11]:
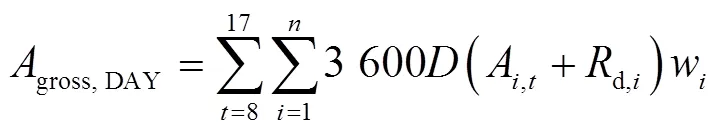
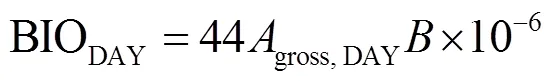
式中gross,DAY為玉米冠層日總同化量,mol/(m2·d);為單位質量的CO2所能產生的生物量,取0.41[11]。
冠層PAR日累積截獲量的計算公式如下
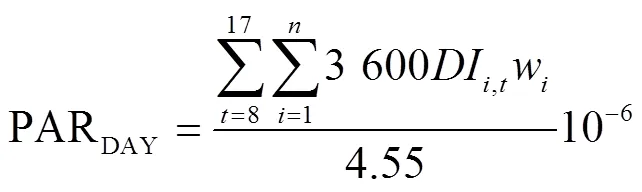
式中PARDAY,4.55為自然光條件下可見光輻照度(W/m2)與光量子通量密度(mol/(m2·s))間的轉換系數(mol/J)[20]。
1.6 模擬情景
將上述玉米冠層光分布模塊的輸出項,即每個葉片PAR截獲強度的日變化,作為冠層光合模塊的輸入項,通過單葉尺度光響應曲線到冠層光合生產模擬的尺度提升,建立玉米冠層光合生產模型3DMaizeCaP。分別選取AD268、JK968和ZD958的三維株型作為品種輸入參數,晴天和陰天下冠層頂部光量子通量密度的日變化為環境輸入參數,利用3DMaizeCaP模型,探究玉米冠層光合速率和光能利用效率對株型差異和環境變化的響應。
1.7 敏感性分析
采用OAT方法進行模型參數敏感性分析,即每次只改變1個參數,根據參數變化對模擬結果的影響程度[35],用相對敏感度(RS)定量光響應曲線中的初始量子效率和最大光合速率對冠層RUE的敏感性大小,其計算公式如下
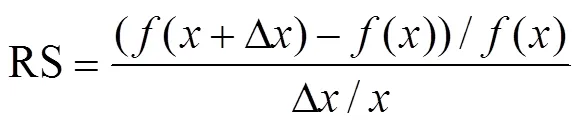
式中為RUE模型參數中的某一項參數,包括初始量子效率α和最大光合速率max,i;?為改參數的改變量;()和(+?)分別表示參數改變前后的RUE模擬值。
1.8 模型編譯和數據處理
模型編譯、數據處理和結果可視化均在RStudio開發環境中使用R語言完成。利用最小二乘法進行光響應曲線擬合,采用LSD法進行單因素方差分析。
2 結果與分析
2.1 玉米株型特征參數
抽雄期JK968的株高最高,達296.3 cm,但其葉片數和穗節位分別為14和12,均顯著(<0.05)低于AD268和ZD958,其葉傾角為65.8°,亦顯著(<0.05)低于AD268和ZD958(圖1)。

注:圖中株型指標為平均值±標準差(n=3),植株三維點云重繪自Wang et al.[36]
2.2 葉片光響應曲線特征參數
由表1可知,在玉米抽雄期,各品種葉片暗呼吸速率和最大光合速率均隨著葉片節位的升高而增大。AD268的上部葉和中部葉的最大光合速率顯著(<0.05)高于JK968和ZD958,分別達到35.86和36.24mol/(m2·s)。AD268葉片光合速率的平均值為28.98mol CO2/(m2·s),較ZD958和JK968相比,分別高10.5%和19.1%(<0.05)。初始量子效率隨葉片節位變化不明顯,AD268、JK968的初始量子效率均值為0.046,ZD958為0.043。
2.3 冠層凈光合速率日變化和葉片CO2日凈同化量模擬
不同品種在陰天和晴天下的冠層凈光合速率均呈先增大,至11:00-12:00達到最大,而后降低的日變化趨勢(圖2)。陰天狀況下,AD268冠層凈光合速率的最大值和平均值分別為21.6和13.6mol/(m2·s),均顯著(<0.05)高于JK968(20.8,12.7mol/(m2·s))和ZD958(19.6,11.9mol/(m2·s)),且全天各時刻的冠層凈光合速率的大小依次為AD268>JK968>ZD958。晴朗狀況下,AD268冠層凈光合速率的最大值和平均值分別為26.2和18.9mol/(m2·s),均顯著(<0.05)高于JK968(24.9,17.1mol/(m2·s))和ZD958(24.4,16.8mol/(m2·s)),且在11:00以前,冠層凈光合速率依次為AD268>ZD958>JK968,14:00以后呈AD268>JK968> ZD958(圖2)。
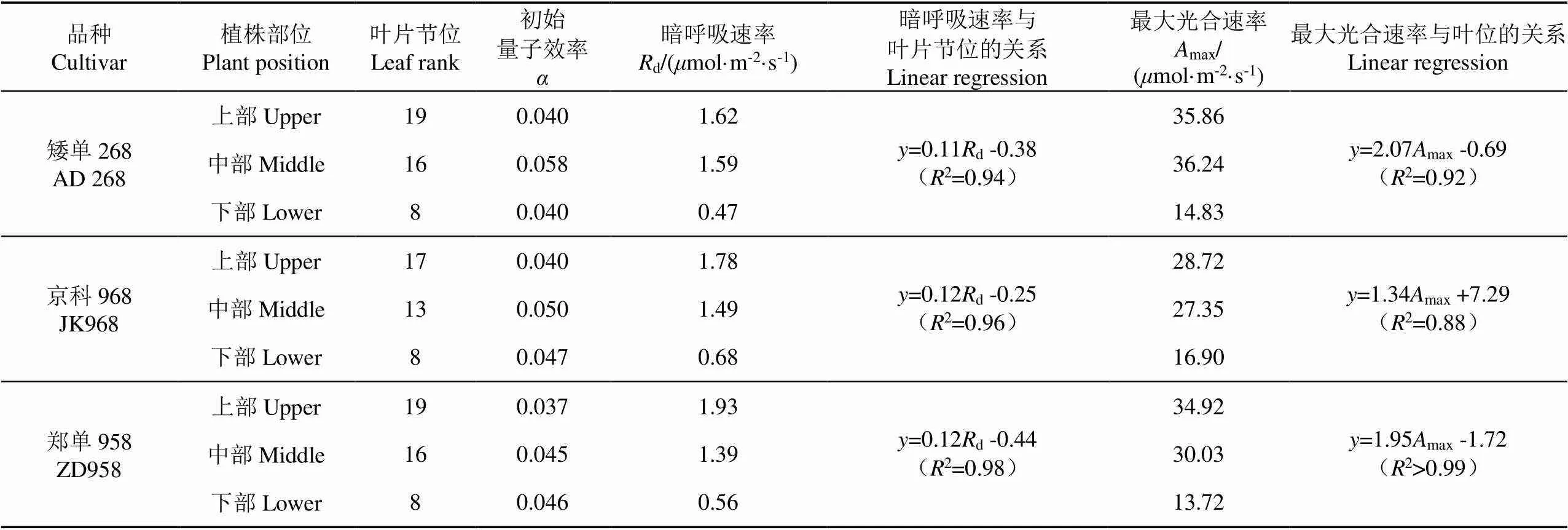
表1 玉米葉片光響應曲線特征參數隨品種和葉位的變化
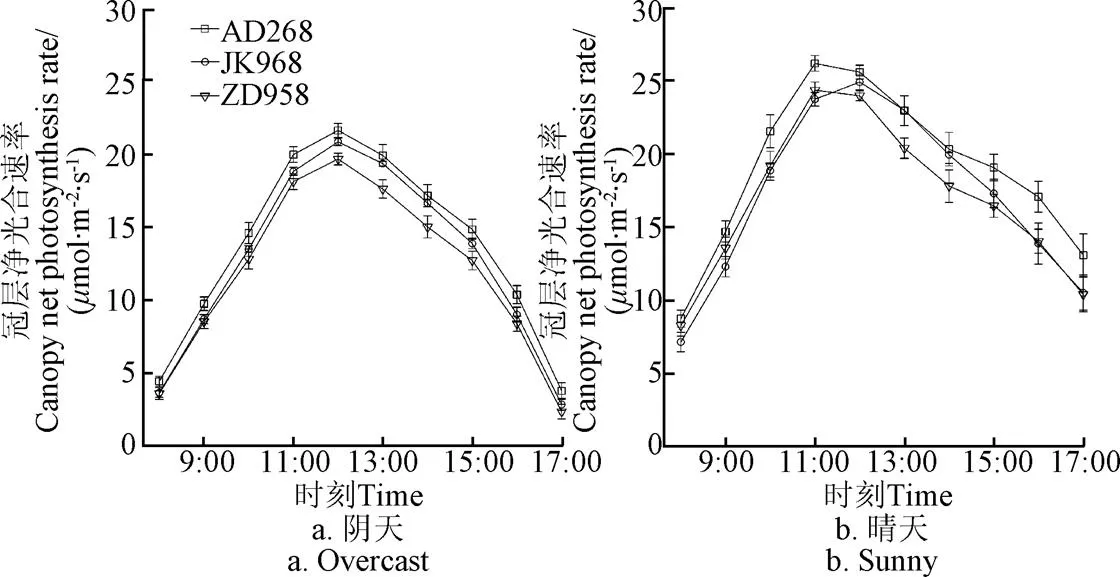
圖2 不同玉米品種在陰天和晴天下冠層凈光合速率的日變化
AD268的CO2日凈同化量在陰天和晴天下分別為491.21和681.25 mmol/(m2·d),比ZD958顯著(<0.05)增加14.8%和12.4%(表2)。各品種單葉CO2日凈同化量隨節位的上升呈先增加后降低的趨勢。JK968、AD268和ZD958的單葉CO2日凈同化量最大值在陰天狀況下依次為10.4、9.1和8.2 mmol/d,而在晴天狀況下依次為15.4、14.0和12.7 mmol/d,除JK968在晴天下的最大值出現第14節位外,其余均出現在第16節位(圖3)。
2.4 冠層PAR截獲量與光能利用效率模擬
陰天狀況下AD268的日生物量積累最大(13.26 g/(m2·d)),與ZD958相比顯著提升6.8 %(<0.05),而晴天狀況下各品種間日生物量積累無顯著差異(表2)。各品種冠層PAR日累積截獲量無顯著差異。各品種冠層光能利用效率差異顯著(<0.05),其中AD268的光能利用效率在陰天和晴天下均為最大,分別為3.22g/MJ和3.03g/MJ,比JK968分別高4.5%和5.6%,比ZD958分別高7.7%和7.8%(表2)。
2.5 初始量子效率和最大光合速率對冠層RUE的影響
從各品種來看,AD268對初始量子效率的變化最為敏感(敏感度為0.834 7),而JK968對最大光合速率的變化最為敏感(敏感度為0.206 8)。玉米冠層光能利用效率對初始光量子效率的敏感度為0.797 8~0.834 7,而對最大光合速率的敏感度為0.169 4~0.206 8,冠層光能利用效率對初始光量子效率的敏感性顯著高于葉片最大光合速率(<0.05)(圖4)。這主要是初始光量子效率的提高能夠顯著增加中下部弱光環境下葉片光合速率所致。
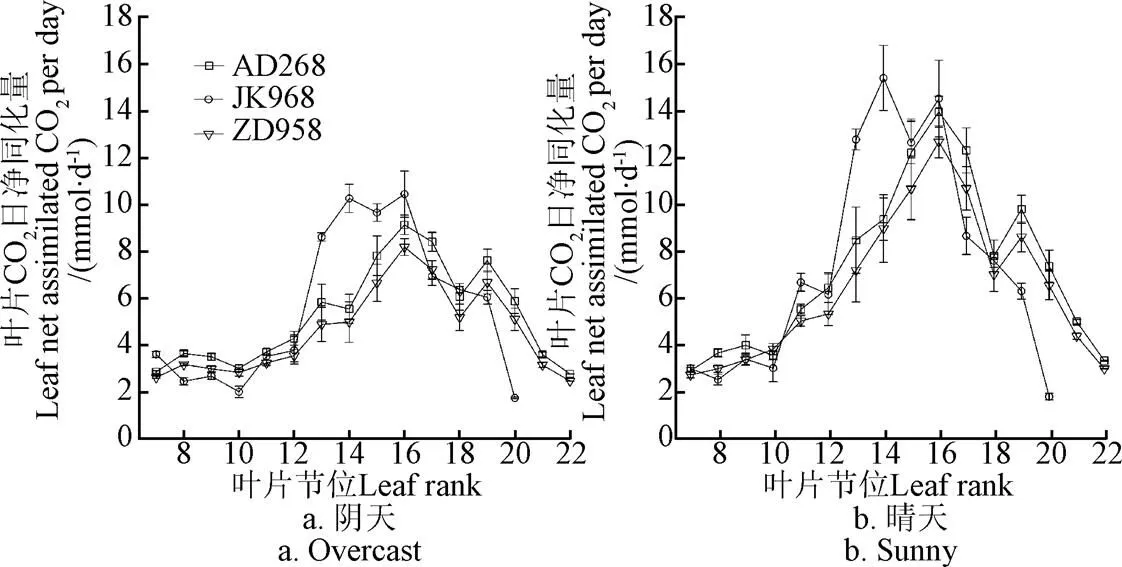
圖3 不同玉米品種在陰天和晴天下各節位葉片CO2日凈同化量
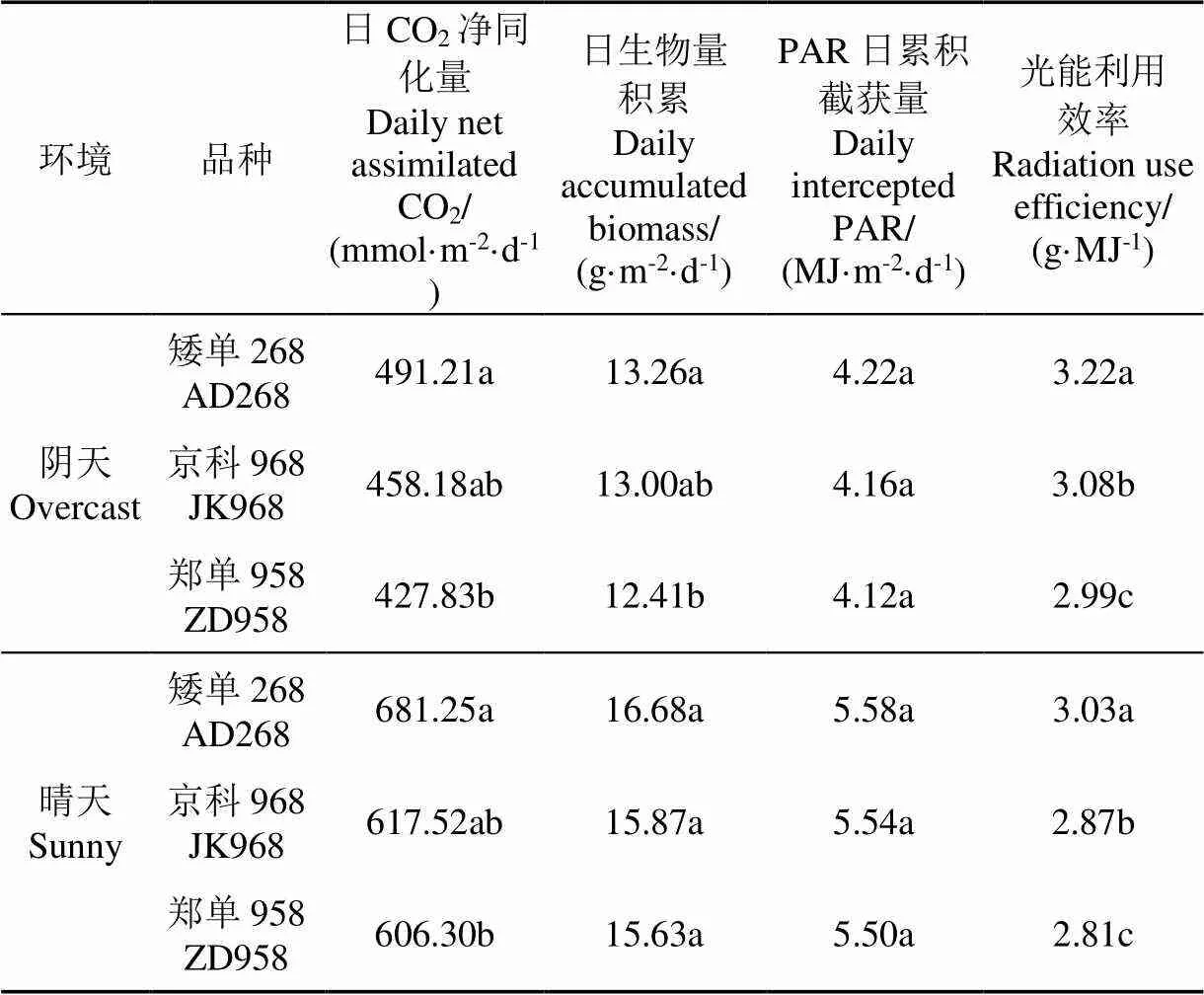
表2 不同玉米品種在陰天和晴天下冠層CO2日凈同化量、日生物量積累、PAR日累積截獲量和光能利用效率
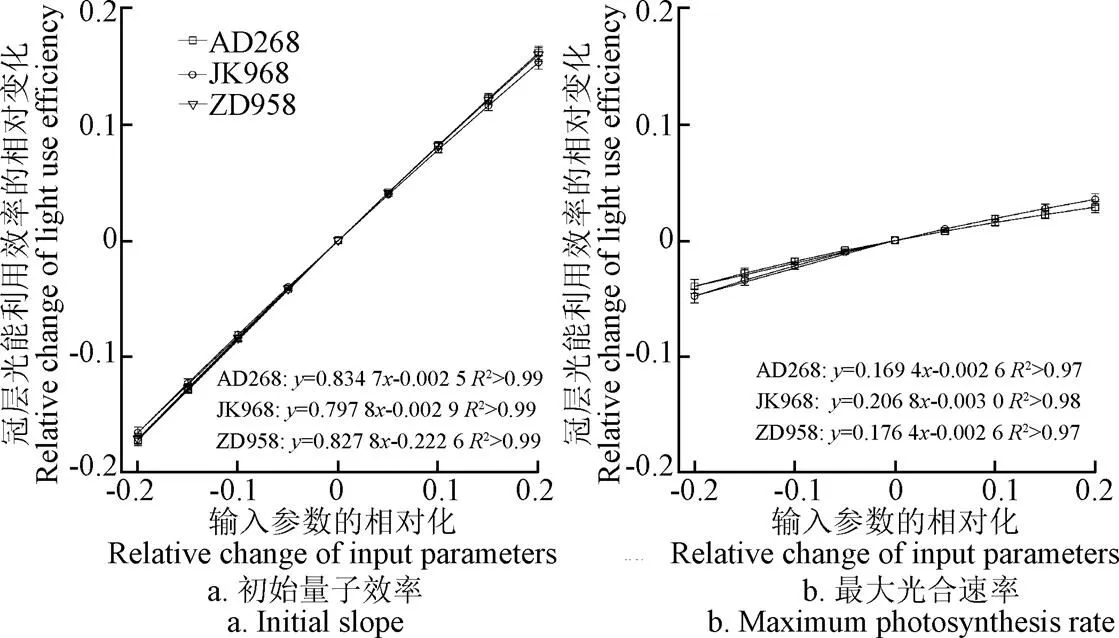
圖4 冠層光能利用效率對改變初始量子效率和最大光合速率的響應
3 討 論
三維重建技術和冠層光線分布模型相結合,可為精確測算器官尺度上的光分布狀況提供一種新技術手段,在此基礎上構建冠層光合作用模型,對于開展冠層光合速率的估算和品種高光效的評價具有重要意義。Cabrera-Bosquet等[37]將植株點云數據與冠層光線分布模型相結合,計算了不同基因型玉米混種群體內單株尺度的PAR日累積截獲量。雖然本研究所采用的冠層光分布模型對植株三維點云數據格式和器官面板數據庫有嚴格的要求[34],但該光分布計算模型為多視角成像[38-39]和LiDAR[40]生成的植株三維點云數據設置了專用接口,未來可為多源異構的植株點云數據提供光分布計算功能,而且該模型可以用于計算器官尺度的PAR截獲量,這為精準評價玉米品種光能利用效率和深入理解植株功能結構參數與冠層光能利用效率間的關系提供了關鍵技術手段。
本研究發現玉米最大冠層凈光合速率為19.6~26.2mol/(m2·s),低于采用多層模型[12,41]測算的結果(約為70mol/(m2·s),以CO2計),造成這種差異的原因一方面是多層模型沒有考慮葉片光合生理特性的冠層垂直分布,另外三維光分布模型和二維多層模型的光分布計算結果也可能存在較大差異。比較分析多種冠層光合作用模型的差異及成因并采用玉米冠層光合速率的實測值對模擬結果進行驗證將是課題組下一步的研究重點。本研究中各玉米品種的冠層PAR截獲量雖無顯著差異,但AD268的冠層日CO2凈同化量顯著高于ZD958,因此前者的RUE也顯著高于后者,大田示范推廣的結果也證實了AD268具有高產、高光效、增產潛力大的特點[42]。
本研究計算所得玉米RUE為2.81~3.22 g/MJ,略低于Lindquist等[10]在高產田試驗的測算結果(3.74 g/MJ),與徐昭等[43]在不同水氮條件下所測得抽雄期的RUE(2.35~4.90 g/MJ)相一致,這表明3DMaizeCaP模型用于測算玉米RUE是合理可行的。夏玉米在陰天狀況下的RUE與晴天狀況相比,平均提高了0.19 g/MJ,這主要是陰天狀況下玉米冠層PAR截獲量的降低幅度大于日生物量積累的降低幅度所致,該結果與Hammer和Wright[44]的研究結果相符。AD268、JK968和ZD958 3個品種的冠層PAR日累積截獲量雖無顯著差異,但由于AD268具有較高的單葉光合速率,冠層可獲得較大的日CO2凈同化量和日生物量積累,最終導致AD268的RUE顯著高于JK968與ZD958。
4 結 論
本文將植物三維模型、冠層光分布模型和葉片光合模型相耦合,實現單葉光響應曲線到冠層光合的尺度提升,構建了玉米冠層光合生產模型3DMaizeCaP,對不同株型玉米在不同環境條件下的冠層光合速率和光能利用效率進行評估。主要結論如下:
1)不同品種夏玉米葉片最大光合速率和暗呼吸速率均隨節位下降呈線性降低的垂直分布規律,矮單268的光合速率最大,而暗呼吸速率最小。
2)冠層凈光合速率日變化趨勢明顯,矮單268的冠層凈光合速率在各時刻均保持最大;矮單268的日CO2凈同化量在陰天和晴天下均顯著高于鄭單958,各葉片中第16節位的單葉日凈同化量達到最大。
3)矮單268、京科968和鄭單958的冠層PAR日累積截獲量無顯著差異(>0.05),矮單268的光能利用效率最大,在陰天和晴天下分別為3.22和3.03g/MJ。
[1] Monteith J L. Climate and the efficiency of crop production in Britain[J]. Philosophical Transactions of the Royal Society of London B, Biological Sciences, 1977, 281(980): 277-294.
[2] Zhu X G, Long S P, Ort D R. What is the maximum efficiency with which photosynthesis can convert solar energy into biomass?[J]. Current Opinion in Biotechnology, 2008, 19(2): 153-159.
[3] Zhu X G, Long S P, Ort D R. Improving photosynthetic efficiency for greater yield[J]. Annual Review of Plant Biology, 2010, 61(1): 235-261.
[4] Long S P, Zhu X G, Naidu S L, et al. Can improvement in photosynthesis increase crop yields?[J]. Plant, Cell & Environment, 2006, 29(3): 315-330.
[5] Sinclair T R, Muchow R C. Radiation use efficiency[J]. Advances in Agronomy, 1999, 65(8): 215-265.
[6] Holzworth D P, Huth N I, deVoil P G, et al. APSIM-Evolution towards a new generation of agricultural systems simulation[J]. Environmental Modelling & Software, 2014, 62: 327-350.
[7] Jones J W, Hoogenboom G, Porter C H, et al. The DSSAT cropping system model[J]. European Journal of Agronomy, 2003, 18(3/4): 235-265.
[8] van Diepen C A, Wolf J, van Keulen H, et al. WOFOST: A simulation model of crop production[J]. Soil Use and Management, 1989, 5(1): 16-24.
[9] Wang E, Martre P, Zhao Z, et al. The uncertainty of crop yield projections is reduced by improved temperature response functions[J]. Nature Plants, 2017, 3(10): 17102.
[10] Lindquist J L, Arkebauer T J, Walters D T, et al. Maize radiation use efficiency under optimal growth conditions[J]. Agronomy Journal, 2005, 97(1): 72-78.
[11] Sinclair T R, Horie T. Leaf nitrogen, photosynthesis, and crop radiation use efficiency: A review[J]. Crop Science, 1989, 29(1): 90-98.
[12] 于強,王天鐸,劉建棟,等. 玉米株型與冠層光合作用的數學模擬研究:Ⅰ.模型與驗證[J]. 作物學報,1998(1):7-15.
Yu Qiang, Wang Tianduo, Liu Jiandong, et al. A mathematical study on crop architecture and canopy photosynthesis I. Model[J]. Acta agronomica sinica, 1998(1): 7-15. (in Chinese with English abstract)
[13] Yin X, Laar H H V. Crop Systems Dynamics: An Ecophysiological Simulation Model for Genotype-by- environment Interactions[M]. Wageningen: Wageningen Academic Publishers, 2005.
[14] Wu A, Hammer G L, Doherty A, et al. Quantifying impacts of enhancing photosynthesis on crop yield[J]. Nature Plants, 2019, 5(4): 380-388.
[15] Song Q, Zhang G, Zhu X G. Optimal crop canopy architecture to maximise canopy photosynthetic CO2uptake under elevated CO2? a theoretical study using a mechanistic model of canopy photosynthesis[J]. Functional Plant Biology, 2013, 40(2): 108-124.
[16] 宋青峰,曲明南,徐建龍,等. 冠層光能利用效率改良的理論、分子途徑及展望[J]. 生命科學,2018,30(10):1044-1050.
Song Qingfeng, Qu Mingnan, Xu Jianlong, et al. The canopy light use efficiency[J]. Chinese Bulletin of Life Sciences, 2018, 30(10): 1044-1050. (in Chinese with English abstract)
[17] Spitters C J T. Separating the diffuse and direct component of global radiation and its implications for modeling canopy photosynthesis.II. Calculation of canopy photosynthesis[J]. Agricultural and Forest Meteorology, 1986, 38(1/2/3): 231-242.
[18] Niinemets ü, Anten NPR. Packing the photosynthetic machinery: From leaf to canopy[C]//Laisk A, Nedbal L, Govindjee. Photosynthesis in silico: Understanding Complexity from Molecules to Ecosystems. Dordrecht: Springer Netherlands, 2009: 363-399.
[19] Sands P. Modelling canopy production. II. from single-leaf photosynthesis parameters to daily canopy photosynthesis[J]. Functional Plant Biology, 1995, 22(4): 603-614.
[20] Goudriaan J, Laar H H V. Modelling Potential Crop Growth Processes Textbook with Exercises[M]. Dordrecht: Springer Netherlands, 1994.
[21] de Pury D G G, Farquhar GD. Simple scaling of photosynthesis from leaves to canopies without the errors of big-leaf models[J]. Plant, Cell & Environment, 1997, 20(5): 537-557.
[22] Vos J, Evers J B, Buck-Sorlin G H, et al. Functional-structural plant modelling: A new versatile tool in crop science[J]. Journal of Experimental Botany, 2010, 61(8): 2101-2115.
[23] 劉镕源,黃文江,任華忠,等. 夏玉米冠層光合有效輻射垂直分布模型[J]. 農業工程學報,2011,27(9):115-121.
Liu Rongyuan, Huang Wenjiang, Ren Huazhong, et al. Photosynthetically active radiation vertical distribution model in maize canopy[J]. Transactions of the Chinese Society of Agricultural Engineering (Transactions of the CSAE), 2011, 27(9): 115-121. (in Chinese with English abstract)
[24] Wu A, Doherty A, Farquhar G D, et al. Simulating daily field crop canopy photosynthesis: An integrated software package[J]. Functional Plant Biology, 2018, 45(3): 362-377.
[25] Wu A, Song Y, van Oosterom E J, et al. Connecting biochemical photosynthesis models with crop models to support crop improvement[J]. Frontiers in Plant Science, 2016, 7: 1518.
[26] Hemmerling R, Kniemeyer O, Lanwert D, et al. The rule-based language XL and the modelling environment GroIMP illustrated with simulated tree competition[J]. Functional Plant Biology, 2008, 35(10): 739-750.
[27] Louarn G, Chenu K, Fournier C, et al. Relative contributions of light interception and radiation use efficiency to the reduction of maize productivity under cold temperatures[J]. Functional Plant Biology, 2008, 35(10): 885-899.
[28] Evers J B, Vos J, Yin X, et al. Simulation of wheat growth and development based on organ-level photosynthesis and assimilate allocation[J]. Journal of Experimental Botany, 2010, 61: 2203-2216.
[29] 葉子飄,于強. 光合作用光響應模型的比較[J]. 植物生態學報,2008,32(6):1356-1361.
Ye Zipiao, Yu Qiang. Comparison of new and several classical models of photosynthesis in response to irradiance[J]. Chinese Journal of Plant Ecology, 2008, 32(6): 1356-1361. (in Chinese with English abstract)
[30] 溫維亮,郭新宇,趙春江,等. 基于三維數字化的玉米株型參數提取方法研究[J]. 中國農業科學,2018,51(6):1034-1044.
Wen Weiliang, Guo Xinyu, Zhao Chunjiang, et al. Research on maize plant type parameter extraction by using three dimensional digitizing data[J]. Scientia Agricultura Sinica, 2018, 51(6): 1034-1044. (in Chinese with English abstract)
[31] 溫維亮,郭新宇,盧憲菊,等. 玉米器官三維模板資源庫構建[J]. 農業機械學報,2016,47(8):266-272.
Wen Weiliang, Guo Xinyu, Lu Xianju, et al. Three-dimensional template resource library construction of maize organs[J]. Transactions of the Chinese Society for Agricultural Machinery, 2016, 47(8): 266-272. (in Chinese with English abstract)
[32] 溫維亮,趙春江,郭新宇,等. 基于t分布函數的玉米群體三維模型構建方法[J]. 農業工程學報,2018,34(4):192-200.
Wen Weiliang, Zhao Chunjiang, Guo Xinyu, et al. Construction method of three-dimensional model of maize colony based on t-distribution function[J]. Transactions of the Chinese Society of Agricultural Engineering (Transactions of the CSAE), 2018, 34(4): 192-200. (in Chinese with English abstract)
[33] 溫維亮. 玉米株型冠層三維數字化與結構解析技術研究[D]. 北京:北京工業大學, 2017.
Wen Weiliang. Research on Three Dimensional Digitization and Architecture Analysis of Maize Plant and Canopy[D]. Beijing: Beijing University of Technology, 2017. (in Chinese with English abstract)
[34] Wen W, Guo X, Li B, et al. Estimating canopy gap fraction and diffuse light interception in 3D maize canopy using hierarchical hemispheres[J]. Agricultural and Forest Meteorology, 2019, 276-277: 107594.
[35] 張寧,張慶國,于海敬,等. 作物生長模擬模型的參數敏感性分析[J]. 浙江大學學報:農業與生命科學版,2018,44(1):107-115.
Zhang Ning, Zhang Qingguo, Yu Haijing, et al. Sensitivity analysis for parameters of crop growth simulation model[J]. Journal of Zhejiang University: Agriculture and Life Sciences, 2018, 44(1): 107-115. (in Chinese with English abstract)
[36] Wang Y, Wen W, Wu S, et al. Maize plant phenotyping: comparing 3D laser scanning, multi-view stereo reconstruction, and 3D digitizing estimates[J]. Remote Sensing, 2018, 11(1): 63.
[37] Cabrera-Bosquet L, Fournier C, Brichet N, et al. High-throughput estimation of incident light, light interception and radiation-use efficiency of thousands of plants in a phenotyping platform[J]. New Phytologist, 2016, 212(1): 269-281.
[38] 胡鵬程,郭焱,李保國,等. 基于多視角立體視覺的植株三維重建與精度評估[J]. 農業工程學報,2015,31(11):209-214.
Hu Pengcheng, Guo Yan, Li Baoguo, et al. Three- dimensional reconstruction and its precision evaluation of plant architecture based on multiple view stereo method[J]. Transactions of the Chinese Society of Agricultural Engineering (Transactions of the CSAE), 2015, 31(11): 209-214. (in Chinese with English abstract)
[39] 韓東,楊貴軍,楊浩,等. 基于立體視覺的玉米雄穗三維信息提取[J]. 農業工程學報,2018,34(11):166-173.
Han Dong, Yang Guijun, Yang Hao, et al. Three dimensional information extraction from maize tassel based on stereoscopic vision[J]. Transactions of the Chinese Society of Agricultural Engineering (Transactions of the CSAE), 2018, 34(11): 166-173. (in Chinese with English abstract)
[40] 郭慶華,吳芳芳,龐樹鑫,等. Crop 3D—基于激光雷達技術的作物高通量三維表型測量平臺[J]. 中國科學:生命科學,2016,46(10):1210-1221.
Guo Qinghua, Wu Fangfang, Pang Shuxin, et al. Crop 3D: A platform based on LiDAR for 3D high-throughputcrop phenotyping[J]. Scientia Sinica Vitae, 2016, 46(10): 1210-1221. (in Chinese with English abstract)
[41] Grant R F, Peters D B, Larson E M, et al. Simulation of canopy photosynthesis in maize and soybean[J]. Agricultural and Forest Meteorology, 1989, 48(1): 75-92.
[42] 段美生,丁書英,張銀生,等. 玉米新品種矮單268適宜栽培密度研究及示范推廣[J]. 中國農技推廣,2017,33(12):16-18.
Duan Meisheng, Ding Shuying, Zhang Yingsheng, et al. Study on the optimal plant density and demonstration and extension of new maize cultivar AD268[J]. China Agricultural Technology Extension, 2017, 33(12): 16-18. (in Chinese with English abstract)
[43] 徐昭,史海濱,李仙岳,等. 水氮限量供給對鹽漬化農田玉米光能利用與產量的影響[J]. 農業機械學報,2018,49(12):281-291.
Xu Zhao, Shi Haibin, Li Xianyue, et al. Effect of limited irrigation and nitrogen rate on radiation utilization efficiency and yield of maize in salinization farmland[J]. Transactions of the Chinese Society for Agricultural Machinery, 2018, 49(12): 281-291. (in Chinese with English abstract)
[44] Hammer G, Wright G. A theoretical analysis of nitrogen and radiation effects on radiation use efficiency in peanut[J]. Australian Journal of Agricultural Research, 1994, 45(3): 575-589.
Modelling maize photosynthesis and light utilization based on a 3D maize canopy model
Gu Shenghao, Wang Yongjian, Wen Weiliang, Lu Xianju, Yu Zetao, Guo Xinyu※
(1.,100097,; 2.,100097,; 3.,100097,)
Light distribution and leaf photosynthesis characteristics are highly heterogeneous within a crop canopy. Maize canopy photosynthesis model based on a 3D canopy structure is an important approach to accurately evaluating radiation use efficiency for cultivars. In this study, we built a maize photosynthetic production model 3DMaizeCaP via coupling canopy 3D architecture model, radiative flux distribution model, leaf photosynthesis model and radiation utilization model. In this study, three cultivars with different plant architecture, i.e., AD268, JK968 and ZD958, and two typical weather conditions, i.e., a sunny day and an overcast day were used. In order to unravel the responses of canopy photosynthesis rate and radiation use efficiency to cultivar and environment, a simulation study combined with field experiment was performed. The results showed that the maximum photosynthesis rate and dark respiration rate decreased linearly with decreasing leaf rank for AD268, JK968 and ZD958. The distribution of both the maximum photosynthesis rate and dark respiration rate of individual leaves showed a vertical profile from the top to the bottom of the maize canopy. The AD268 had the highest maximum photosynthesis rate and the lowest dark respiration rate among three cultivars. The diurnal course of canopy photosynthesis rate was characterized evidently that canopy photosynthesis rate increased in the morning and reached the maximum value at 12:00 of noon on an overcast day and at 11:00 on a sunny day and then decreased in the afternoon for all cultivars. The maximum canopy photosynthesis rate of AD268 was 21.6mol CO2/(m2·s) on an overcast day and was 26.2mol CO2/(m2·s) on a sunny day, which were significantly higher than that of JK968 (20.8mol CO2/(m2·s) and 24.9mol CO2/(m2·s)) and of ZD958 (19.6mol CO2/(m2·s) and 24.4mol CO2/(m2·s)). The daily net assimilated CO2of AD268 was significantly (<0.05) higher than that of ZD958. In comparison with ZD958, the daily net assimilated CO2increased by 14.8% and 12.4% on a sunny and an overcast day respectively. The plant architecture of AD268 was significantly different with other cultivars (<0.05). However, there was no significant difference in the daily intercepted photosynthetic absorbed radiation between cultivars (>0.05). The leaf at 16th main stem phytomer rank produced the highest daily net assimilated CO2among individual leaves at the leaf level. The radiation use efficiency of AD268 was 3.22 and 3.03 g/MJ under a sunny and an overcast condition, respectively, indicating a 4.5% and a 5.6% increase compared to JK968 and a 7.7% and a 7.8% compared to ZD958. The canopy radiation use efficiency of maize was more sensitive to the initial slope of light response curve than to the maximum photosynthesis rate (<0.05). From the point view of improving canopy radiation use efficiency for maize, designing a maize ideotype that has a more compact plant architecture and higher leaf photosynthetic capacity was suggested for breeding in the future. This study could provide not only an approach for quantitatively estimating canopy photosynthesis rate of maize but also an evaluation basis as well as a phenotyping technique for breeding cultivars with high photosynthetic efficiency.
photosynthesis; crops; models; canopy light distribution; 3-D point cloud; plant structural-functional model; light response curve; radiation use efficiency
顧生浩,王勇健,溫維亮,盧憲菊,于澤濤,郭新宇. 基于三維冠層模型的玉米光合作用和光能利用模擬[J]. 農業工程學報,2019,35(20):178-184.doi:10.11975/j.issn.1002-6819.2019.20.022 http://www.tcsae.org
Gu Shenghao, Wang Yongjian, Wen Weiliang, Lu Xianju, Yu Zetao, Guo Xinyu. Modelling maize photosynthesis and light utilization based on 3D maize canopy model[J]. Transactions of the Chinese Society of Agricultural Engineering (Transactions of the CSAE), 2019, 35(20): 178-184. (in Chinese with English abstract) doi:10.11975/j.issn.1002-6819.2019.20.022 http://www.tcsae.org
2019-06-03
2019-09-27
國家重點研發計劃課題(2016YFD0300605);國家自然科學基金(31871519);北京市農林科學院創新能力建設專項(KJCX20180423);北京市農林科學院科研創新平臺建設(PT2019-24)
顧生浩,博士,助理研究員,主要從事作物生長模擬模型研究。Email:gu.shenghao@outlook.com
郭新宇,博士,研究員,主要從事數字植物理論研究。Email:guoxy@nercita.org.cn
10.11975/j.issn.1002-6819.2019.20.022
S127
A
1002-6819(2019)-20-0178-07
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
網絡安全與數據管理(2022年1期)2022-08-29 03:15:20
導航定位學報(2022年4期)2022-08-15 08:27:00
中學生數理化·中考版(2022年8期)2022-06-14 06:55:24
新世紀智能(數學備考)(2021年9期)2021-11-24 01:14:36
成都醫學院學報(2021年2期)2021-07-19 08:35:14
新世紀智能(數學備考)(2020年9期)2021-01-04 00:25:14
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
數學物理學報(2020年2期)2020-06-02 11:29:24
光學精密工程(2016年6期)2016-11-07 09:07:19
