基于表型和SRAP標記的盆栽菊遺傳多樣性分析
2020-01-04 07:14:15沈瑤王晗璇侯海嫻吳智明周厚高
熱帶作物學報 2020年11期
沈瑤 王晗璇 侯海嫻 吳智明 周厚高
摘 ?要:基于20個表型性狀和相關序列擴增多態性(sequence-related amplified polymorphism,SRAP)標記技術對30個國外引進盆栽菊品種遺傳多樣性進行研究。結果表明,參試盆栽菊品種表型變異豐富,性狀變異系數為14.67%~ 78.25%,最低和最高變異系數分別為冠幅和舌狀小花數。性狀主成分分析結果顯示,前5個主成分貢獻率達76.68%,綜合反映了花型大小、葉片形態和花色的重要性。基于表型性狀的聚類分析可將30個盆栽菊品種劃為3個類群:第Ⅰ類群包含14個品種;第Ⅱ類群包含2個品種;第Ⅲ類群包含14個品種。表型性狀的聚類結果與花徑和觀花期有一定聯系。SRAP分析篩選到11對多態性引物,獲得1866個多態性位點,多態性比例為92.61%。品種間的遺傳相似系數為0.67~0.85,表明30個品種存在一定的遺傳變異。基于品種間遺傳距離構建的NJ(neighbor joining)進化樹結果顯示,30個盆栽菊品種可劃分為3個類群:第Ⅰ類群包含2個亞群,共17個品種;第Ⅱ類群包含10個品種;第Ⅲ類群包含3個品種。2種分類方法均將研究對象分為3個類群,但這3個類群的品種組成并不一致,推測可能和菊花材料本身復雜的遺傳背景與試驗選材的局限性有關。
關鍵詞:盆栽菊花;遺傳多樣性;表型;SRAP標記;聚類分析
中圖分類號:S682.1 ? ? ?文獻標識碼:A
Genetic Diversity Analysis of the Potted Chrysanthemum Based on Phenotype and SRAP Marker
SHEN Yao, WANG Hanxuan, HOU Haixian, WU Zhiming, ZHOU Hougao*
School of Horticulture and Landscape Architecture, Zhongkai University of Agriculture and Engineering, Guangzhou, Guangdong 510225, China
Abstract: In order to better protect chrysanthemum germplasm resources and lay a foundation for hybrid breeding, the genetic diversity of 30 potted chrysanthemum varieties were studied based on 20 morphological and SRAP markers. The results showed that the phenotypic variation of potted chrysanthemum varieties was abundant. The coefficient of variation of characters was ranged from 14.67% to 78.25%, which were crown width and tongue floret number. The results of principal component analysis showed that the contribution rate of the first five principal components was 76.68%, which comprehensively reflected the importance of flower size, leaf shape and flower color. Based on the cluster analysis of phenotypic traits, 30 potted chrysanthemum cultivars could be divided into three groups. GroupⅠ, GroupⅡ and Group Ⅲ consisted of 14, 2 and 14 varieties correspondingly. The clustering results based on phenotypic traits were related to flower diameter and natural florescence period. Based on SRAP marker technology, 11 pairs of polymorphic primers were screened out and a total of 1866 polymorphic loci were obtained, with the polymorphic proportion as high as 92.61%. The genetic similarity coefficient between cultivars was ranged from 0.67 to 0.85, indicating that there were some genetic variations in the 30 cultivars. The samples could be divided into three groups based on the NJ tree. GroupⅠ, Group Ⅱ and Group Ⅲ consisted 17, 10 and 3 varieties correspondingly. However, the clustering results of the two grouping techniques showed that there was no good correlation between the two markers, which might be related to the complex genetic background of chrysanthemum materials and the limitations of experimental selection.
Keywords: potted chrysanthemum; genetic diversity; phenotype; SRAP marker; cluster analysis
DOI: 10.3969/j.issn.1000-2561.2020.11.003
菊花(Chrysanthemum × morifolium Ramat.)起源于中國,花型、花色和株型等千姿百態,品種資源異常豐富。據報道,全世界菊花品種約有20 000~30 000種,中國約有3 000多種[1]。栽培菊花大多為六倍體(2n = 6x = 54)或非整倍體,基因組高度雜合且十分龐大(約12.381~24.802 Gb),遺傳背景非常復雜[2]。菊花種質資源的鑒定、保存和評價是生產利用和選育菊花新品種的基礎,對菊花品種更新和產業升級具有重要意義。國內外有眾多學者先后對菊花種質資源進行了表型性狀研究,并對菊花種質資源進行分析和整理,為加快優良種質資源的利用和菊花育種奠定了堅實基礎[3-6]。近年來,隨著生物技術的快速發展,如RAPD[7-8]、AFLP[9-10]、ISSR[11-12]、SSR[13-15]和SRAP[16-19]等多種分子標記技術也先后應用于菊花種質資源遺傳多樣性研究中,結果顯示菊花在基因型上有豐富的遺傳多樣性。然而,結合表型性狀和SRAP分子標記對盆栽小菊品種進行遺傳多樣性研究還鮮見報道。
本研究基于表型性狀和SRAP分子標記技術,對國外引進的30份盆栽菊花品種資源進行遺傳多樣性分析,探討其遺傳多樣性在形態和DNA水平上的關聯,旨在為盆栽菊雜交育種過程中親緣關系分析、雜交親本的選配提供一定的參考。
1 ?材料與方法
1.1 ?材料
試驗材料為課題組從國外引進的30個盆栽小菊品種(表1)。2017年下半年栽植于廣州市白云區寮采村白云現代農業示范基地。SRAP引物為Li等[20]公布的通用引物,正/反向引物各8條,共產生64對引物組合。所有引物經熒光標記,供機器讀圖。
1.2 ?方法
1.2.1 ?表型性狀的測定 ?本試驗選取14個數量性狀(花徑、花數、觀花期、花梗長、頭狀花序高度、舌狀小花長度、舌狀小花寬度、葉長、葉寬、葉數、株高、莖粗、冠幅、節間距)和6個質量性狀與多態性狀(花色、花心顏色、葉緣裂刻深淺、莖顏色、葉基部形態和抗旱性)共20個表型性狀,依據《植物新品種特異性、一致性和穩定性(DUS)測試指南—菊花》(中華人民共和國農業行業標準NY/T 2228?2012)在盛花期對各觀賞性狀進行觀測。每個品種調查10株,每個數量性狀測量3次,取平均值。變異系數(CV)=(標準差SD/平均值 )×100%。
1.2.2 ?SRAP擴增與引物篩選 ?采用常規CTAB法提取菊花葉片DNA。SRAP擴增體系總體積為25 μL,其中含模板DNA 2 μL,10×PCR buffer(含MgCl2)2.5 μL,10 mmol/L dNTPs 0.5 μL,10 μmol/L上下游引物各0.5 μL,Taq DNA聚合酶0.5 μL,ddH2O 18.5 μL。擴增反應程序為:94 ℃預變性5 min;94 ℃ 1 min,35 ℃ 1 min,72 ℃ 1 min,5個循環;然后再94 ℃ 1 min,50 ℃ 1 min,72 ℃ 1 min,35個循環;最后72 ℃延伸10 min,4 ℃保存。擴增產物在聚丙烯酰胺凝膠上電泳,保存電泳膠圖,由北京鼎國生物技術公司完成。
1.3 ?數據統計與處理
1.3.1 ?表型數據的統計分析與作圖 ? 數量性狀數據直接記錄,質量性狀數據轉換成相應數字代碼。具體包括:花色(白色或近白色=1,黃綠色= 2,綠色=3,淡黃色=4,黃色=5,橙色=6,粉紅色=7,紅色=8,淺紫色=9,紫色=10);花心顏色(白色=1,綠色=2,黃綠色=3,淡黃色=4,黃色= 5,橙黃色=6,橙色=7,紅棕色=8,棕色=9,棕黑=10,紫黑色=11);葉柄姿態(極向上=1,向上=3,平伸=5,向下=7,下垂=9);葉基部形態(銳角=1,鈍角=2,圓=3,平截=4,心形=5);葉緣裂刻深淺(淺=3,中=5,深=7);莖顏色(綠色=1,綠棕色或綠紫色=2,棕色=3,紫色=4);抗旱性(根據自然環境10 d不澆水后植株葉片的萎蔫程度來評價,抗旱=5,中抗=3,不抗旱=1),將相應數值錄入Excel 2010。表型性狀的變異分析、主成分分析和基于歐式距離的聚類分析與作圖均用SPSS18.0軟件完成。
1.3.2 ?SRAP條帶統計分析與作圖 ?利用GENESCAN 3.1軟件分析保存的膠圖。參照熒光標記Marker(Rox500)大小(70、80、90、100、120、140、160、180、190、200、220、240、260、280、300、320、340、360、380、400、425、450、475、490、500 bp,共25條帶)在70~500 bp范圍內每2 bp讀1次數,共讀取216個數(相當于216個位點)。而后通過Binthere 軟件將提取的片段大小結果轉變為0-1矩陣(有帶的記為“1”,無帶的記為“0”)。Excel 2010統計總位點數和多態性位點數,計算多態性比率。用MEGA-X軟件根據遺傳距離進行NJ(neighbor joining)聚類分析和作圖。
2 ?結果與分析
2.1 ?盆栽菊品種表型性狀的多樣性分析
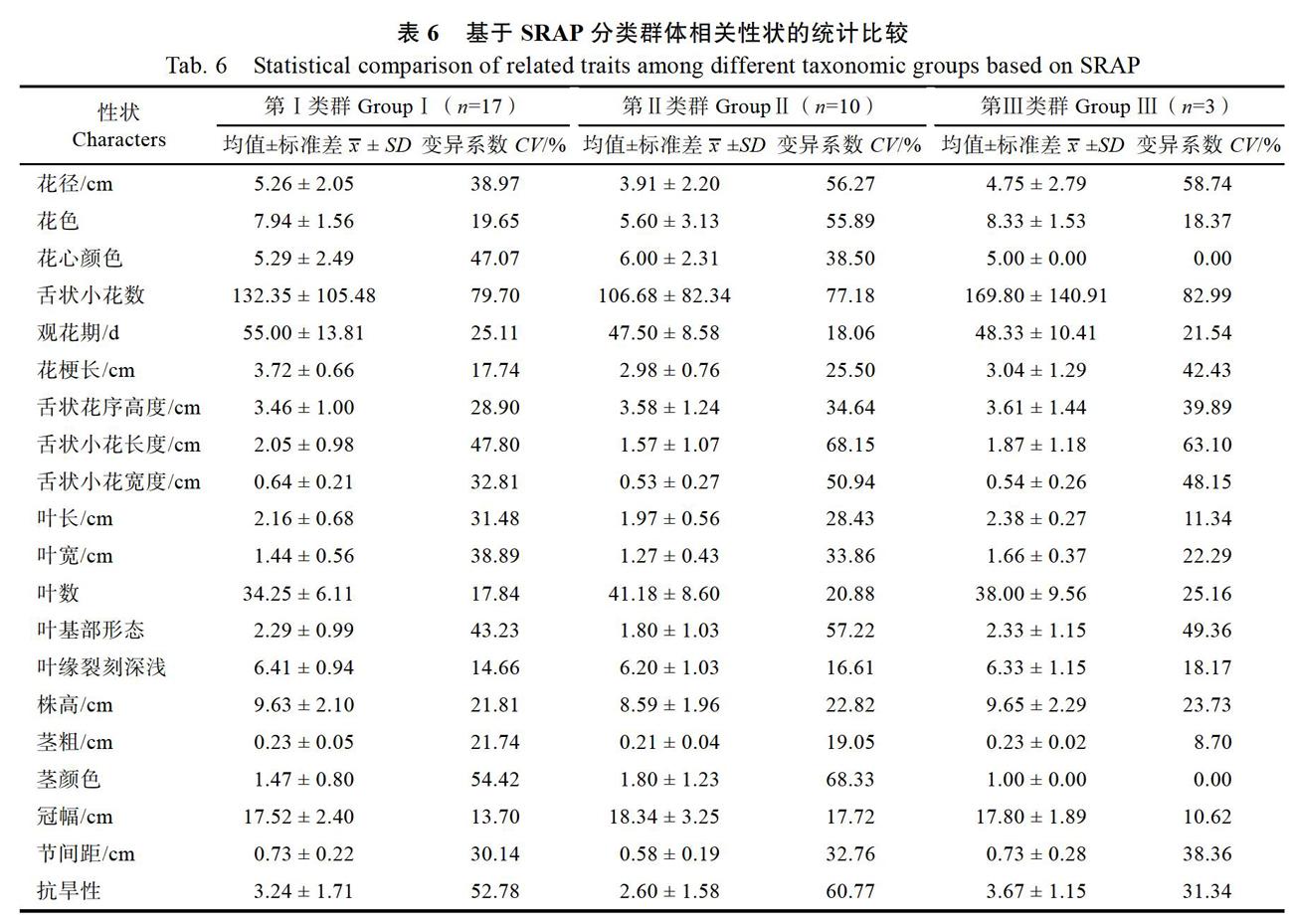
對30個盆栽菊品種的20個農藝性狀進行了調查統計分析。結果表明,不同盆栽菊品種各農藝性狀表現出不同程度的多樣性(表2),變異系數為14.67%~78.25%。調查的20個性狀中,舌狀小花數的變異系數最大,為78.25%;其次為莖顏色,變異系數為61.12%;變異系數在40%以上的性狀還有舌狀小花長度、抗旱性、葉基部形態、花徑和花心顏色等。變異系數小于20%的性狀有冠幅(14.67%)和葉緣裂刻深淺(15.14%)。
2.2 ?表型性狀的主成分分析
對30個盆栽菊品種20個表型性狀進行主成分分析,提取特征值大于1的前5個主成分,累計貢獻率達76.68%(表3)。第1主成分特征值為7.60,貢獻率為38.02%,對應的特征向量中花徑相關系數最大(0.94),其次為舌狀小花長度(0.88)、節間距(0.81)、舌狀小花寬度(0.78)等,反映了花序大小等狀況,可認為是花形態因子;第2主成分特征向量絕對值較大的性狀為葉寬(0.81)、葉長(0.72),主要反映了葉的性狀,可認為是葉形態因子;第3主成分特征值為1.96,貢獻率為9.78%,對應的特征向量中相關系數較大的有舌狀小花數(0.58)、花心顏色(0.56)和花色(?0.54),可概況為花色因子;第4、5主成分特征向量花梗長(0.54)、花心顏色(0.67)和莖顏色(0.59)等,可以看作花形態因子的補充。這些因子的綜合表現主要是盆栽菊的花部,包括花徑、舌狀小花長和寬、舌狀小花數、花色和花心顏色等。
2.3 ?基于表型性狀的聚類分析
將表型性狀的原始數據進行標準化處理,采用組間聯接進行聚類分析,在歐式距離為17.5可以將30個盆栽菊品種分為3個類群(圖1)。
第Ⅰ類群包含2個亞群,共14個品種。其中第1亞群含13個品種:‘西奧(8號)、‘卡斯蒂赫(30號)、‘紅粉佳人(13號)、‘粉草莓(3號)、‘馬里奧(14號)、‘德里粉(2號)、‘麥克(23號)、‘黃草莓(9號)、‘凱撒(18號)、‘羅賓漢粉(20號)、‘迪迪(24號)、‘威廉(19號)和‘夏之戀(15號);第2亞群包含1個品種:‘綠色心情(29號)。
第Ⅱ類群包含2個品種:‘羅馬里奧粉(1號)和‘紅日(5號)。
第Ⅲ類群包含3個亞群,共14個品種。其中第1亞群包含2個品種:‘驕陽多彩(21號)和‘驕陽輝煌(28號);第2亞群包含6個品種:‘朝陽熱情(10號)、‘滿天星火紅(12號)、‘滿天星蓬勃(11號)、‘朝陽繁華(22號)、‘滿天星蜜桃(4號)和‘拉米雷斯(6號);第3亞群包含6個品種:‘滿天星白(26號)、‘小乒純潔(27號)、‘驕陽自由(16號)、‘驕陽卡薩(17號)、‘小行星(7號)和‘小乒糖心(25號)。
根據分類結果,再將表型數據進行分類統計,查找類群與表型的關系。從整體看(表4),第Ⅰ類群花徑、舌狀小花數、舌狀小花長度和寬度、冠幅等性狀均值高于其他兩個類群;第Ⅱ類群的葉長、葉寬、株高、徑粗和節間距等性狀的均值高于類群Ⅰ和類群Ⅲ,植株生長勢相對較強;第Ⅲ類群花徑、花期、花梗長、舌狀花序高度、舌狀小花長度和寬度、葉長和葉寬、株高、徑粗、冠幅、節間距等性狀的均值都是3個類群中最小的,且抗旱性表現相對較差,所有性狀中僅葉數和葉緣裂刻深淺2個性狀均值最大。由此說明,分類結果與表型性狀之間有一定的聯系。
2.4 ?SRAP引物篩選及擴增結果多態性分析
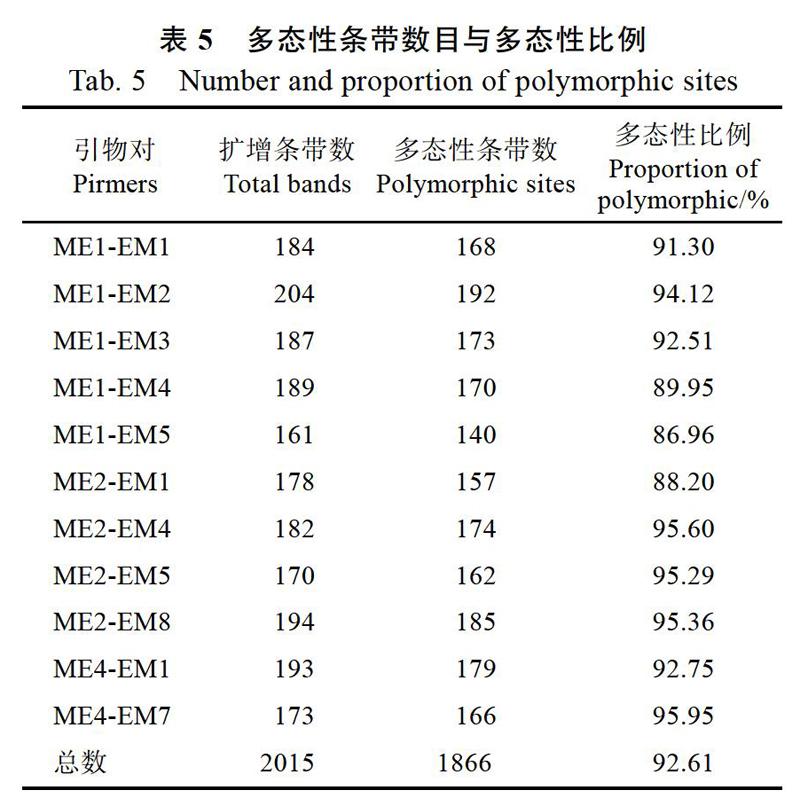
從64對引物中篩選出11對能擴增出較多特異性條帶、且條帶清晰穩定的引物。11對引物對30個盆栽菊品種進行SRAP擴增,共獲得2015條清晰可辨讀條帶,其中1866條具有多態性,多態性比例為92.61%(表5)。
2.5 ?基于SRAP的聚類分析
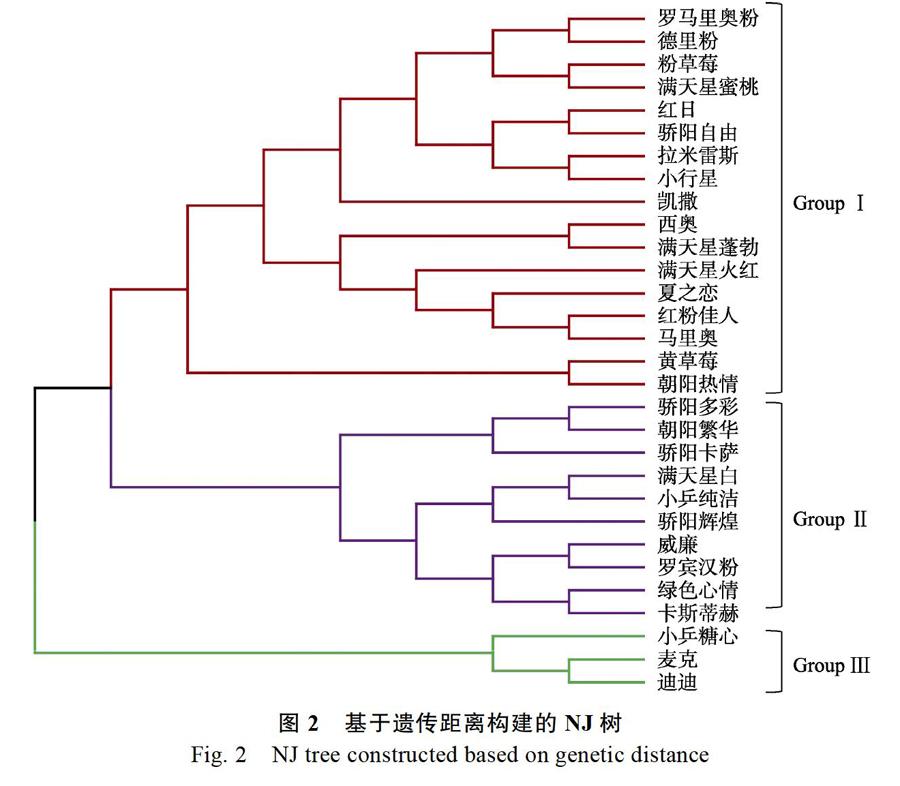
30份盆栽菊種質的遺傳相似系數變化范圍為0.67~0.85,表明30個品種間存在一定的遺傳變異。基于30個盆栽菊品種間的遺傳距離,采用neighbor joining算法構建了NJ樹(圖2)。結果表明,30個盆栽菊品種可劃分為3個類群:
第Ⅰ類群包含2個亞群,共17個品種。其中第1亞群包含15個品種:‘羅馬里奧粉(1號)、‘德里粉(2號)、‘粉草莓(3號)、‘滿天星蜜桃(4號)、‘紅日(5號)、‘驕陽自由(16號)、‘拉米雷斯(6號)、‘小行星(7號)、‘凱撒(18號)、‘西奧(8號)、‘滿天星蓬勃(11號)、‘滿天星火紅(12號)、‘夏之戀(15號)、‘紅粉佳人(13號)、‘馬里奧(14號);第2亞群包含2個品種:‘黃草莓(9號)和‘朝陽熱情(10號)。
第Ⅱ類群包含10個品種:‘驕陽多彩(21號)、‘朝陽繁華(22號)、‘驕陽卡薩(17號)、‘滿天星白(26號)、‘小乒純潔(27號)、‘驕陽輝煌(28號)、‘威廉(19號)、‘羅賓漢粉(20號)、‘綠色心情(29號)和‘卡斯蒂赫(30號)。
第Ⅲ類群包含3個品種:‘小乒糖心(25號)、‘麥克(23號)和‘迪迪(24號)。
根據SRAP分類結果,將同一類群中個體的表型數據進行分類統計,查找類群與表型的可能關系(表6)。結果顯示,不論是葉長、葉寬、葉數,還是株高、莖粗、節間距和冠幅等,各類群差異不明顯。第Ⅰ類群花徑稍大,花期稍長,但差異不明顯。說明SRAP分類結果與表型性狀的相關性較差。
3 ?討論
3.1 ?盆栽菊表型遺傳多樣性
形態學標記作為傳統的分類方法,因其操作簡單,非常直觀,在園藝植物種質資源的整理與評價及遺傳多樣性分析中經常用到[21-28],在菊花上也有不少報道[1, 4, 12, 17, 29-33]。收集整理和積累菊花相關的形態學數據是應用現代生物技術加速菊花種質改良的基礎。本研究對30個盆栽菊品種的20個表型性狀進行統計,并對獲得的信息數據進行數量分類學聚類,結果顯示,供試盆栽菊品種在形態學水平上有非常豐富的變異,特別是舌狀小花數量、莖顏色等變異系數高。對20個表型性狀進行主成分分析,發現花徑、舌狀小花長度、舌狀小花寬度和花期等是影響花部形態的主要因子。基于表型性狀的聚類結果按照花徑?觀花期分類:第I類群平均花徑6.66 cm,觀花期長(60 d);第II類群平均花徑5.51 cm,觀花期中等(55 d);第III類群平均花徑2.75 cm,觀花期短(43 d)。花色在聚類結果中的規律不是很明顯,研究結果與張冬菊等[34]的報道一致。
3.2 ?盆栽菊SRAP標記遺傳多樣性
SRAP標記具有操作簡便、重復性好、多態性高、成本相對較低等優勢,是目前植物遺傳多樣性及親緣關系研究中最常用的分子標記技術之一,在國蘭[35]、鳶尾[36-37]、杜鵑花[38]、唐菖蒲[28]、萬壽菊[39]及菊花[17-18]等多種觀賞園藝植物遺傳多樣性研究中有相關報道。
本研究利用SRAP標記技術對從國外引進的30個盆栽菊品種進行遺傳多樣性分析,從64對引物組合中篩選獲得11對多態性引物,共擴增得到1866個多態性位點,多態性比例達92.61%,顯著高于已報道的利用RAPD標記技術分析菊花遺傳多樣性的多態性比例(69.00%)[7],與已報道的利用AFLP標記技術分析遺傳多樣性的多態性比例(92.80%)[40]和利用ISSR標記技術分析遺傳多樣性的多態性比例(92.50%)[41]等相當,說明SRAP標記有比較豐富的遺傳信息。依據多態性標記位點建立的30個盆栽菊品種NJ樹結果顯示,30個材料可歸為3個類群,但歸類的結果與表型性狀的聚類分析結果有差異。
3.3 ?基于表型和SRAP分類方法結果的比較分析
本研究基于表型和SRAP標記技術分別對30個國外引進的盆栽菊品種進行聚類分析,2種分類方法均將研究對象分為3個類群,但是這3個類群的品種組成并不一致。這一結果與很多以往的研究報道一致[21, 28, 42-46]。造成2種分類方法結果不一致的原因是多方面的,首先菊花本身的遺傳背景十分復雜,而本研究利用的形態性狀不足、分子標記探查到的遺傳差異有限,不能全面反映品種的遺傳信息。其次,形態標記和分子標記是2種不同形式的標記,分子標記反映的是種質間基因型的差異,而形態標記揭示的是某些功能基因在內部和外部環境的共同作用下表達的差異,即基因與環境互作的結果。表型的表現往往是非常復雜的,即使遺傳基礎很相似,即DNA水平的差異小,但基因受轉錄水平、翻譯水平的調控機制異常復雜,因此表型常常因基因間的互作或調控呈現復雜多樣,這也是許多作物中表型分類與分子標記分類不完全一致的主要原因。
總之,基于表型和分子標記技術分析物種的遺傳多樣性都是可行且十分必須的。盡管聚類結果可能出現不一致的情況,甚至和傳統的分類結果出現很大差異。在今后的研究中,可以將三者或更多的分析方法相結合,綜合運用多種評價體系,將更有利于對不同品種的來源、特性進行更為準確地分析,能更好地評價菊花種質的遺傳多樣性。
參考文獻
[1] 張莉俊, 戴思蘭. 菊花種質資源研究進展[J]. 植物學報, 2009, 44(5): 526-535.
[2] Sasaki K, Mitsuda N, Nashima K, et al. Generation of expressed sequence tags for discovery of genes responsible for floral traits of Chrysanthemum morifolium by next-generation sequencing technology[J]. BMC Genomics, 2017, 18(1): 683.
[3] 郝京輝, 游 ?捷, 秦賀蘭, 等. 菊花品種的特異性、一致性和穩定性的研究[J]. 中南林學院學報, 2003(5): 14-18.
[4] 許瑩修. 菊花形態性狀多樣性和品種分類的研究[D]. 北京: 北京林業大學, 2005.
[5] 邵清松, 郭巧生, 張志遠. 藥用菊花種質資源遺傳多樣性的ISSR分析[J]. 中草藥, 2009(12): 1971-1975.
[6] 楊旭旭. 菊花品種表型性狀與SCoT分子標記的關聯分析[D]. 開封: 河南大學, 2015.
[7] 秦賀蘭, 游 ?捷, 高俊平. 菊花18個品種的RAPD分析[J]. 園藝學報, 2002(5): 488-490.
[8] 徐文斌, 郭巧生, 王長林. 藥用菊花遺傳多樣性的RAPD分析[J]. 中國中藥雜志, 2006(1): 18-21.
[9] 吳在生, 李海龍, 劉建輝, 等. 65個菊花栽培品種遺傳多樣性的AFLP分析[J]. 南京林業大學學報(自然科學版), 2007(5): 67-70.
[10] 劉路賢. 菊花栽培品種遺傳多樣性的AFLP分析及SSR分子標記的開發[D]. 開封: 河南大學, ?2013.
[11] 程 ?華, 李琳玲, 張心玲, 等. 應用ISSR-PCR對10個菊花品種進行遺傳多樣性分析[J]. 湖北農業科學, 2011(20): 4292-4297.
[12] 歐陽彩虹. 44個菊花(Dendranthema×gradifolium (Ramat) Kitam.) 品種的形態學研究與ISSR分析[D]. 重慶: 西南大學, 2010.
[13] Zhang Y, Wang C, Ma H, et al. Assessing the genetic diversity of Chrysanthemum cultivars with microsatellites[J]. Journal of the American Society for Horticultural Science, 2013, 138(6): 479-486.
[14] Wang H B, Jiang J F, Chen S M, et al. Next-generation sequencing of the Chrysanthemum nankingense (Asteraceae) transcriptome permits large-scale unigene assembly and SSR marker discovery[J]. PLoS One, 2013, 8(4): e62293.
[15] Feng S G, He R F, Lu J J, et al. Development of SSR mar-kers and assessment of genetic diversity in medicinal Chrysanthemum morifolium cultivars[J]. Front Genet, 2016(7): 113.
[16] Zhang F, Chen S M, Chen F D, et al. SRAP-based mapping and QTL detection for inflorescence-related traits in Chrysanthemum (Dendranthema morifolium)[J]. Molecular Breeding, 2011, 27(1): 11-23.
[17] 李仁偉,王 ?晨,戴思蘭,等. 菊花品種表型性狀與SRAP分子標記的關聯分析[J]. 中國農業科學, 2012, 45(7): 1355-1364.
[18] 張冬菊, 李世超, 吳鵬夫, 等. 基于表型和SRAP標記的切花菊品種遺傳多樣性分析[J]. 園藝學報, 2014, 41(1): 118-130.
[19] Li P R, Zhang F, Chen S M, et al. Genetic diversity, population structure and association analysis in cut Chrysanthemum (Chrysanthemum morifolium Ramat.)[J]. Molecular Genetics and Genomics, 2016, 291(3): 1117-1125.
[20] Li G, Quiros C F. Sequence-related amplified polymorphism (SRAP), a new marker system based on a simple PCR reaction: its application to mapping and gene tagging in Brassica[J]. Theoretical & Applied Genetics, 2001, 103(2-3): 455-461.
[21] 傅鴻妃, 呂曉菡, 陳建瑛,等. 辣椒種質表型性狀與SSR分子標記的遺傳多樣性分析[J]. 核農學報, 2018, 32(7): 1309-1319.
[22] 赫 ?衛, 張 ?慧, 董延龍, 等. 辣椒種質資源形態學性狀相關性、主成分與聚類分析[J]. 北方園藝, 2018(4): 9-17.
[23] 黃如葵, 孫德利, 張 ?曼, 等. 苦瓜遺傳多樣性的形態學性狀聚類分析[J]. 廣西農業科學, 2008(3): 351-356.
[24] 夏碧波, 李 ?穎, 王恒明, 等. 國外引進辣椒資源形態學性狀的聚類分析[J]. 分子植物育種, 2017, 15(8): 3318-3330.
[25] 楊婷婷, 于澤群, 夏樂晗, 等. 君遷子(Diospyroslotus)種質資源形態學性狀的聚類分析[J]. 果樹學報, 2014, 31(4): 566-573.
[26] 袁海靜, 安 ?巍, 李立會, 等. 中國枸杞種質資源主要形態學性狀調查與聚類分析[J]. 植物遺傳資源學報, 2013, 14(4): 627-633.
[27] 趙德新, 孫治強, 任子君, 等. 茄子形態學性狀主成分分析及聚類分析[J]. 河南農業大學學報, 2009, 43(4): 393-397.
[28] 劉 ?晨, 高明偉, 劉 ?超, 等. 基于表型和SRAP標記的唐菖蒲品種遺傳多樣性分析[J]. 中國農業大學學報, 2016, 21(5): 57-65.
[29] 邵清松, 郭巧生, 李育川, 等. 藥用菊花種質資源形態變異的數量分析[J]. 中國中藥雜志, 2011(10): 1261-1265.
[30] 吳國盛, 陳發棣, 陳素梅, 等. 部分菊屬與亞菊屬植物的形態學聚類及親緣關系分析[J]. 南京農業大學學報, 2009, 32(1): 155-159.
[31] 楊 ?秋. 昆明地區菊花種質資源的調查、分類及評價[D]. 重慶: 西南林學院, 2007.
[32] 楊 ?雨. 開封市菊花品種資源多樣性的研究[D]. 開封: 河南大學, 2008.
[33] 楊 ?旭, 毛 ?燕, 吳德智, 等. 50個大菊品種形態學性狀的比較[J]. 浙江農業科學, 2016(6): 864-867.
[34] 張冬菊, 李世超, 吳鵬夫, 等. 基于表型和SRAP標記的切花菊品種遺傳多樣性分析[J]. 園藝學報, 2014, 41(1): 118-130.
[35] 唐源江, 曹雯靜, 吳坤林. 基于SRAP標記的國蘭種質資源遺傳多樣性分析及分子身份證構建[J]. 中國農業科學, 2015, 48(9): 1795-1806.
[36] 許玉鳳, 閆小風, 邵美妮, 等. 基于SRAP的15種鳶尾的遺傳多樣性和親緣關系分析[J]. 沈陽農業大學學報, 2015, 46(5): 568-574.
[37] 童 ?俊, 毛 ?靜, 董艷芳, 等. 鳶尾屬部分種質資源遺傳多樣性的SRAP分析[J]. 湖北農業科學, 2019, 58(4): 88-92.
[38] 肖 ?政, 蘇家樂, 劉曉青, 等. 杜鵑花種質資源遺傳多樣性的SRAP分析[J]. 江蘇農業學報, 2016(2): 442-447.
[39] 侯志強, 唐 ?楠, 冶有林, 等. 萬壽菊雜交后代優選株系的SRAP-PCR遺傳多樣性分析[J]. 基因組學與應用生物學, 2017, 36(10): 4295-4306.
[40] 韓 ?潔, 胡 ?楠, 李玉閣, 等. 菊花品種資源遺傳多樣性的AFLP分析[J]. 園藝學報, 2007(4): 1041-1046.
[41] 繆恒彬, 陳發棣, 趙宏波. 85個大菊品種遺傳關系的ISSR分析[J]. 園藝學報, 2007(5): 1243-1248.
[42] 李可峰, 韓太利, 董貴俊, 等. 用形態與分子標記研究石刁柏種質資源遺傳多樣性[J]. 植物遺傳資源學報, 2006(1): 59-65.
[43] 沈程文, 寧正祥, 黃建安, 等. 基于表型參數及SRAP標記的廣東茶樹種質遺傳多樣性[J]. 應用生態學報, 2009, 20(7): 1551-1558.
[44] 許先松, 劉志欽, 林曉丹, 等. 基于形態及SRAP標記的辣椒資源遺傳多樣性及親緣關系比較[J]. 福建農林大學學報(自然科學版), 2011, 40(1): 48-53.
[45] 趙 ?靚, 羅燕杰, 肖思文, 等. 基于表型和SSR標記的梅花種質資源遺傳多樣性分析[J]. 分子植物育種, 2019, 17(13): 4458-4469.
[46] 張耀元, 任彥鑫, 禾 ?璐, 等. 基于表型性狀和SSR標記的山西省青狗尾草資源遺傳多樣性分析[J]. 中國農業大學學報, 2016, 21(10): 11-22.
收稿日期 ?2019-12-24;修回日期 ?2020-01-27
基金項目 ?廣州市科技計劃民生專題項目(No. 201903010053)。
作者簡介 ?沈 ?瑤(1981—),女,碩士研究生,研究方向:園林植物與觀賞園藝。*通信作者(Corresponding author):周厚高(ZHOU Hougao),E-mail:zhouhougao@163.com。
