棉花2個胚胎發育晚期富集蛋白基因的克隆與表達分析
2020-01-07 09:58:54甄軍波宋世佳劉琳琳李海山遲吉娜
華北農學報 2019年6期
甄軍波,劉 迪,宋世佳,劉琳琳,蔡 肖,張 曦,李海山,遲吉娜
(1.河北省農林科學院 棉花研究所,農業部黃淮海半干旱區棉花生物學與遺傳育種重點實驗室,河北 石家莊 050051; 2.河北省農林科學院,河北 石家莊 050051)
胚胎發育晚期富集蛋白(Late embryogenesis abundant proteins, LEA)最早在棉花中發現,是在棉花種子成熟后期大量積累的一類親水蛋白,在水稻、大麥、小麥、玉米和擬南芥等植物中均有LEA家族基因的報道。根據LEA家族基因氨基酸的序列特征,可以將LEA蛋白家族分為LEA1、LEA2、LEA3、LEA4、LEA5、LEA6、SMPS和dehydrins 8個亞組,每一個亞組都具有獨特的氨基酸保守區域[1]。研究表明,LEA家族基因不僅存在于植物的種子中,在幼苗、芽和根等部位也能檢測到該基因家族的存在[2-6]。同時,在細菌和無脊椎動物中也檢測到了LEA蛋白的存在。LEA家族基因廣泛參與植物生長發育、形態建成和衰老等生物學過程,在高鹽、干旱等非生物脅迫下,LEA蛋白還能夠對植株起到關鍵的保護作用[6-7]。Wang等[8]從丹參中分離得到LEA2家族的SmLEA2基因,該基因能顯著提高轉基因丹參的耐旱性和耐鹽性。Lü等[9]從百慕大草中克隆得到了2個dehydrins亞家族基因,CdDHN4-L和CdDHN4-S,2個基因編碼的氨基酸序列相差1個φ片段,均能夠顯著提高擬南芥對于高鹽脅迫的抗性以及大腸桿菌對于高鹽和極端溫度的耐性,并且轉CdDHN4-L比轉CdDHN4-S的效果更加明顯。LEA蛋白家族在植物應答生物脅迫中也發揮重要作用,玉米LEA3亞組成員在煙草中過表達后,提高了對假單胞致病菌的耐性[10],在大腸桿菌中過表達擬南芥LEA2和LEA4亞組成員能顯著抑制細菌的生長[11]。
研究人員最早從棉花中發現LEA蛋白家族,并對LEA基因家族的序列特征和表達模式進行了初步的研究[4, 12-14]。Luo等[15-16]從棉花中分離得到LEA4亞組的D113基因的啟動子片段,啟動子序列中含有與LEA蛋白特異表達相關的順式作用元件,并能夠受ABA、高鹽和干旱脅迫誘導表達。劉峰等[17]從棉花新陸早33中分離得到了D34基因啟動子的同源序列,并驗證了其表達模式。Magwanga等[6]利用公布的棉花基因組數據,分別從陸地棉、亞洲棉和雷蒙德氏棉中分離得到242,136和142個LEA家族基因,并將這些家族成員分為8個亞組,大多數LEA家族成員與棉花耐旱性有關,LEA基因家族成員在耐旱材料夏威夷棉中的表達量明顯高于干旱敏感的陸地棉材料。Magwanga等[18]的研究表明,陸地棉LEA2家族成員,CotAD_20020、CotAD_21924和CotAD_59405能顯著提高轉基因擬南芥的根長和耐旱性。可見,棉花中LEA基因家族在棉花逆境脅迫應答中發揮著重要的作用。本研究從棉花均一化cDNA文庫中篩選得到2個棉花LEA家族的基因,命名為GhLEA和GhLEAG1,對其進行了生物信息學分析和非生物脅迫條件下的表達模式分析,旨在為進一步研究棉花LEA家族基因的功能提供參考。
1 材料和方法
1.1 試驗材料
供試陸地棉品種冀228為生物技術課題組保存。
1.2 試驗方法
1.2.1 脅迫處理方法 參照Zhang等[19]、甄軍波等[20-21]的方法種植冀228水培苗,并對幼苗進行NaCl處理(150 mmol/L)、ABA(100 μmol/L)處理和17% PEG6000處理,對照和脅迫處理的根、莖和葉分別于脅迫處理后0,1,2,3,6,24 h取樣;纖維取樣:棉花開花后8DPA、12DPA、16DPA、20DPA、24DPA、30DPA和35DPA樣品分別取樣。所有樣品均液氮速凍,用于RNA提取。
1.2.2 棉花RNA的提取和反轉錄 采用天根試劑盒(DP441)提取棉花RNA;采用TaKaRa (RR0037A)試劑盒合成cDNA第一鏈。
1.2.3 LEA基因的克隆和序列分析 在對文庫測序結果的分析過程中,從中篩選得到59號和378號2個克隆,用ORFfinder 對59號和378號克隆包含完整的開放閱讀框進行預測,利用BlastX進行基因的同源性比對,用ClustalX和MEGA 4.0進行序列比對并構建系統進化樹。利用DANMAN預測蛋白理化性質,NetPhos 3.1預測氨基酸序列磷酸化位點,SOMPA和Phyre2分別預測蛋白質的二級結構和三級結構,MEME預測氨基酸序列的保守motif,并用TBtools繪制motif組成圖[21]。
1.2.4 棉花LEA基因表達分析 用Primer Premier 5軟件設計LEA基因的熒光定量PCR引物(表1),選取Histone3為內參基因,qRT-PCR反應參照TB GreenTMPremix ExTaqTMⅡ(Tli RNaseH Plus)試劑盒說明書:95 ℃ 30 s;95 ℃ 15 s,60 ℃ 15 s,72 ℃ 30 s,40個循環,最后進行溶解曲線分析,采用Bio-rad熒光定量PCR儀,試驗進行3次重復。用Bio-Rad CFX Manager(2-ΔΔCT法)和Origin 6.0分析處理試驗結果[21]。

表1 試驗中所用的引物Tab.1 Primers used in this study
2 結果與分析
2.1 棉花LEA基因的克隆和序列分析
通過對本研究室前期構建的cDNA文庫進行分析,59號克隆和378號克隆均包含一個完整的開放閱讀框。PCR擴增2個基因的全長,與cDNA文庫測序結果完全一致。在NCBI網站,利用BlastX程序進行分析比對,59號克隆和378號克隆分別編碼一個棉花胚胎發育晚期富集蛋白家族基因成員,根據比對結果,將其命名為GhLEA和GhLEAG1,登錄號分別為KF906314和KF906316。GhLEA全長948 bp,編碼315個氨基酸,等電點4.4,分子質量為35 ku,GhLEAG1全長756 bp,編碼251個氨基酸,等電點10.76,分子質量27 ku,2個基因DNA序列相似度為36.93%,氨基酸序列相似度為10.44%。
2.2 棉花GhLEA和GhLEAG1基因進化樹及保守基序分析
選取GhLEA、GhLEAG1、SmLEA(AAU29064.3)、SmLEA2(HQ676610)和部分擬南芥LEA家族成員構建了氨基酸序列進化樹(圖1),結果表明,GhLEA、SmLEA和SmLEA2同屬于LEA2家族,GhLEA和SmLEA2具有更近的親緣關系,其與At2G44060和SmLEA2的motif組成模式一致,均包含了motif1/2/6/7,motif6在LEA2家族中高度保守。GhLEAG1與擬南芥LEA3家族的成員劃分在一個亞組,但分支成員之間的motif組成不完全相同,GhLEAG1由motif3/9組成,AT3G53770和AT4G15910由motif3/10組成,GhLEA5-D、AT4G02380和AT1G02820則由motif3/8組成,可見motif3在LEA3家族成員中高度保守。

A.棉花GhLEA、GhLEAG1和部分擬南芥LEA氨基酸序列進化樹分析; B.棉花GhLEA、GhLEAG1和部分擬南芥LEA蛋白保守基序組成。 A. Phylogenetic tree analysis of GhLEA, GhLEAG1 and closely related Arabidopsis LEA proteins; B. Motif analysis of GhLEA, GhLEAG1 and closely related Arabidopsis LEA proteins.
2.3 棉花LEA基因蛋白質結構分析
參考甄軍波等[21]的方法,利用NetPhos分析GhLEA和GhLEAG1的磷酸化位點(圖2),GhLEA肽鏈具有較多的絲氨酸磷酸化位點,蘇氨酸和酪氨酸磷酸化位點較少,GhLEAG1肽鏈中蘇氨酸磷酸化位點明顯增多,酪氨酸磷酸化位點較少。用SOPM法分析GhLEA蛋白的二級結構,GhLEA蛋白包括22.22%的α-螺旋,25.71%的延伸鏈,6.03%的β-轉角和46.03%的無規則卷曲,GhLEAG1蛋白包括18.33%的α-螺旋,33.07%的延伸鏈,3.59%的β-轉角和45.02%的無規則卷曲,通過預測2個蛋白的三級結構可以看出(圖3),兩者在三級結構上也存在明顯的差別。
2.4 棉花LEA基因表達模式分析
分別研究了GhLEA和GhLEAG12個基因在纖維中的表達模式。結果表明,GhLEA在棉纖維發育20DPA時表達量達到峰值,同時,在根、莖和葉片中,GhLEA在根中的表達量最高,在葉片中的表達量最低(圖4-A)。GhLEAG1在棉纖維中的表達量峰值同樣出現在20DPA,在莖中的表達量最高(圖4-B)。GhLEA可能只在棉纖維發育時期中的伸長期發揮作用,而GhLEAG1在伸長期及次生壁加厚期均可能發揮比較重要的作用。

A.GhLEA; B.GhLEAG1。
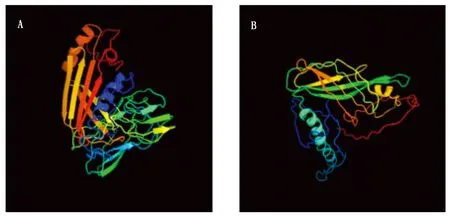
A.GhLEA三維結構; B.GhLEAG1三維結構。 A.Predicted 3D structure of GhLEA; B. Predicted 3D structure of GhLEAG1.

誤差線上的不同大寫字母表示0.01水平差異顯著(P<0.01)。 A. GhLEA基因在不同組織中的表達模式; B.GhLEAG1基因在不同組織中的表達模式。 Bars superscripted by different capital letters mean significant difference(P<0.01). A.Tissue expression analysis of GhLEA; B.Tissue expression analysis of GhLEAG1.
分別驗證了GhLEA和GhLEAG1在NaCl、PEG和ABA非生物脅迫條件下的表達模式。在NaCl處理條件下,GhLEA在根、莖和葉片中的表達量均有升高,在莖和葉片中的表達量峰值現在處理后3 h,在根中的表達量峰值則出現在處理后6 h。在PEG處理條件下,GhLEA在根的表達量逐漸下降,在莖中的表達量呈先降后升的趨勢,在處理后6 h表達量達到最低,24 h后表達量恢復至對照水平。葉片中的表達量則呈上升趨勢,峰值出現在處理后3 h。在棉花幼苗受到ABA處理條件下,GhLEA的表達量峰值出現在處理后3 h(圖5-A)。通過圖5-B可以看出,GhLEAG1的表達模式同GhLEA具有明顯的區別。在高鹽脅迫條件下,GhLEAG1在根中的表達量迅速下降,在莖中的表達量則是輕微的波動,而在葉片中的表達量則是顯著提高,并且在處理后3 h達到峰值。在受到PEG處理時,GhLEAG1在根中的表達量同樣是迅速下降,在莖中的表達量在6 h開始下降,在葉片中的表達量則出現在處理后6 h達到峰值。GhLEAG1同樣受ABA誘導表達,表達量在處理后6 h達到峰值。

誤差線上的不同大寫字母表示0.01水平差異顯著(P<0.01)。A.GhLEA基因表達模式分析; B.GhLEAG1基因表達模式分析。 Bars superscripted by different capital letters mean significant difference(P<0.01).A. Expression analysis of GhLEA; B.Expression analysis of GhLEAG1.
3 討論
棉花中前期已經報道了GhD-7、GhD11、GhD-19、GhD-29、GhLEA14-A、GhLEA5-D、GhLEAD-113、GhLEAD-34等8個LEA蛋白家族成員,根據其氨基酸序列的相似度和motif組成,這8個家族成員分別屬于LEA1-5及Dehydrin家族成員[8]。陸地棉LEA2蛋白家族成員最多,有157個家族成員,根據其序列和motif結構組成可以將其分為6個亞組[18]。
本研究從棉花均一化cDNA文庫中的2個LEA蛋白家族成員GhLEA和GhLEAG1,分別編碼315,251個氨基酸,與棉花中已報道的8個LEA蛋白家族成員分析比對,GhLEA和GhLEAG1屬于新的LEA家族成員。研究結果表明,GhLEA屬于棉花LEA2中的一員,GhLEAG1則屬于LEA3亞組,qRT-PCR結果表明,兩者在棉花纖維、根、莖和葉片中均有表達,兩者均受高鹽、PEG和ABA等非生物脅迫誘導表達,但是GhLEA和GhLEAG1可能在棉纖維發育的伸長期和次生壁加厚期分別發揮作用,兩者在蛋白質二級結構和三級結構上也具有比較明顯的差異。Gao等[22]研究結果表明,即使在同一個亞組中,不同成員由于其motif組成及表達模式的差異,在功能上也會有所差異。從構建的系統進化樹可以看出,GhLEA與丹參SmLEA和SmLEA2具有較近的親緣關系,都屬于LEA2家族成員,但是由于SmLEA由motif4/5/6組成,GhLEA、SmLEA2和SmLEA分別在不同的分支上。Hundertmark和Hincha[3]研究表明,At2G44060能夠受脅迫誘導表達。Wang等[8]和Wu等[23]的研究結果也表明,SmLEA和SmLEA2均能夠提高大腸桿菌和轉基因丹參的耐鹽性,但是其組織部位表達特異性存在差異,可能與其motif組成有關。GhLEA也能受到高鹽等非生物脅迫的誘導表達,且GhLEA與At2G44060和SmLEA2的motif組成模式完全一致,推測其可能與SmLEA2具有相似的功能。植物LEA3家族成員也可能參與生物和非生物脅迫應答[5]。GhLEAG1則屬于棉花LEA3家族,與棉花中已經克隆的GhLEA5-D同在一個亞組,但是motif組成模式具有一定的差別,可能屬于LEA3亞家族中的一個小分支,與GhLEA5-D等基因發揮不同的作用。
